 2020, Vol. 40
2020, Vol. 40文章信息
- 李林, 魏识广, 练琚愉, 曹洪麟
- LI Lin, WEI Shiguang, LIAN Juyu, CAO Hongling
- 亚热带不同纬度植物群落物种多样性分布规律
- Distributional regularity of species diversity in plant community at different latitudes in subtropics
- 生态学报. 2020, 40(4): 1249-1257
- Acta Ecologica Sinica. 2020, 40(4): 1249-1257
- http://dx.doi.org/10.5846/stxb201812012615
-
文章历史
- 收稿日期: 2018-12-01
- 网络出版日期: 2019-11-06






2. 广西师范大学生命科学学院, 珍稀濒危动植物生态与环境保护教育部重点实验室, 桂林 541006;
3. 广西珍稀濒危动物生态学重点实验室, 桂林 541006;
4. 中国科学院华南植物园, 广州 510650
2. College of Life Science, Guangxi Normal University, Key Laboratory of Ecology of Rare and Endangered Species and Environmental Protection, Ministry of Education, Guilin 541004, China;
3. Key Laboratory of Ecology of Rare and Endangered Animal Ecology of Guangxi(Guangxi Normal University), Guilin 541006, China;
4. South China Botanical Garden, Chinese Academy of Science, Guangzhou 510650, China
生物多样性空间分布相关机制一直是生态学、生物地理学和保护生物学研究的热点问题[1]。研究森林群落物种多样性在纬度梯度上的变化特征是揭示生物多样性与生态因子两者相互关系的重要方式[2]。有研究通过数据挖掘发现物种丰富度与纬度有明显的负相关性, 随纬度升高而降低。Simpson多样性指数和Shannon-Wiener指数与纬度的关系同物种丰富度类似, 均呈现出随纬度升高而降低的趋势;优势度与纬度之间呈现正相关, 即表现为随纬度升高优势度增大的趋势。也有前人的研究证实:在排除经度干扰之后, 植物群落的物种多样性是随着纬度的升高而降低的, 而植物群落物种重要值随着纬度的升高、物种多样性的降低, 其值是增大的, 也就表明物种多样性低的植物群落中其优势树种很明显, 此优势树种在该群落中也就起着主导决定性作用, 是不可替代的。土壤含水率随纬度的升高会降低, 这就表明纬度低的地区森林植被的含水量比纬度高的地区要高[3], 故间接导致纬度低的群落其植物物种类型更多样化。
我国亚热带地区地域广阔, 亚热带地区森林的物种多样性高于北美同纬度地区, 而暖温带地区森林群落的物种多样性低于北美同纬度地区[2, 4]。常绿阔叶林为我国亚热带森林的地带性植被, 属于亚热带森林生物多样性的主体。但由于纬度分布和海拔等因素的不同, 各常绿阔叶林群落的物种组成、优势种的优势度、以及多样性程度都存在差异。基于此, 本研究选取两广地区的猫儿山, 南岭以及鼎湖山三个国家级自然保护区内的四个常绿阔叶林群落进行比较研究, 探索不同纬度、海拔的群落多样性分布规律。为亚热带, 特别是南亚热带森林群落的保护提供理论依据。
1 研究地概况研究地选取在两广地区的猫儿山, 南岭以及鼎湖山三个国家级自然保护区内。猫儿山国家级自然保护区地处广西壮族自治区桂林市兴安县境内, 在东经110°30′, 北纬25°56′, 属中亚热带湿润山地季风气候区[5], 研究样地位于保护区海拔1850—1950m的八角田中山盆地和海拔1450—1500m的红军亭附近, 文中分别称为八角田样地和红军亭样地。八角田样地属于常绿落叶针阔叶混交林, 红军亭样地林型为常绿落叶阔叶混交林。
南岭国家级自然保护区位于广东省北部南岭山脉的核心地带。东经112°41′—113°15′, 北纬24°39′—28°08′。南岭山地属亚热带季风气候[6], 样地取址于在大顶山保护站桐桥的丘陵低山常绿阔叶林区域内, 海拔479—579m。
鼎湖山国家级自然保护区位于广东肇庆市东北郊, 地处北回归线南侧, 植被地带性独一无二, 位于东经112°30′39″—112°33′41″, 北纬23°09′21″—23°11′30″, 属南亚热带季风湿润型气候, 样地植被类型为南亚热带常绿季雨林, 具有亚热带向热带林过渡的特征, 海拔600—700m[7]。
2 研究方法 2.1 样地调查在猫儿山自然保护区八角田、红军亭与南岭自然保护区大顶山保护站和鼎湖山自然保护区, 用相邻格子法顺序取样分别设置建立100 m × 100 m的方形样地, 面积为1 hm2, 以20 m × 20 m为大样方单元, 共有25个大样方, 用经纬仪精确测量后设立样方角桩, 在每个样方的角桩上标明数字以辨别它在样地上的位置。每个样方内再划分成16个5 m × 5 m的小样方(小格), 对小样方同样按顺序进行编号。之后对样地内的木本植物(DBH≥1 cm)进行每木检尺, 记录其种名、胸径、树高、枝下高、冠幅、生活型、环境因子, 测定坐标并编号挂永久标牌。
2.2 种-个体曲线统计方法在上述四个样地中, 物种调查原始数据经过核对检验后, 按巢氏取样法, 首次随机取样200株, 统计其内的物种数, 而后每次增加200株, 直至取样1ha样地全部植株而统计每次对应的物种数, 把四个样地统计的一系列数据对应到XY坐标图绘制成种-个体曲线。
2.3 径级统计按5cm为基本单位进行径级划分和数据统计, 即从0—5cm为第一个径级计算此区间的植株个体数, 5—10cm为第二个径级, 之后以此类推, 统计后数据见图 2。
2.4 重要值计算
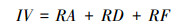
|
式中IV为乔木树种的重要值, RA为相对多度, 等于样地中某种乔木的多度占样地内所有乔木株数之和的百分比, RA在一定程度上决定种群的能流、资源利用及生产力;RD为相对显著度, 是某乔木的显著度占样地所有乔木总显著度的百分比, 在某种程度上是植物利用及影响环境程度的指标;RF为相对频度, 是群落中某乔木出现的频度占群落乔木出现总频度的百分比。
2.5 多样性指数计算本文选用3种α多样性指数进行衡量4个样地的物种多样性, 依次为Shannon-Weiner指数[8]、Simpson指数[9]、Pielou指数[10]。Shannon-Weiner指数的计算公式:
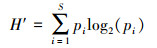
|
式中, p为物种i的相对多度, S为种研究样地中的物种数。
任意两个体其为不同物种的概率为p时, Simpson指数D [9]:
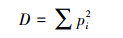
|
Simpson指数也常被称为优势度指数, 因为它很大程度上受优势种的影响。Simpson指数值增大时候表示多样性减小, 因而又常把Simpson指数表述为1-D或者1/D (本文用1-D来表征样性, 值越大表明多样性越高)。
Pielou的均匀度指数的计算公式[10]:
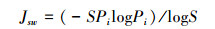
|
式中, P为物种i的相对多度, S为种研究样地中的物种数。值越大表明多样性越高。
除计算α多样性指数外, 本研究还计算了两个β多样性指数:Sorensen指数, Jaccard指数, 两者的计算公式:
Jaccard指数
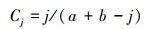
|
Sorensen指数

|
式中, j为两个群落或样地共有种数, a和b分别为样地A和样地B的物种数[11]。
Sorensen指数和Jaccard指数表征四个群落两两之间的物种相似程度。当两个群落所含有的种完全相同时, 其系数为最大值1;当两个群落所含有的种完全不同时, 其系数为0;系数自0到最大值之间, 表示两个群落相似程度。
3 研究结果分析 3.1 种-个体分布对样地中的种-个体分布进行统计, 种-个体曲线显示(图 1):猫儿山的八角田样地的种-个体数分布相对红军亭样地比较平缓, 物种数随取样个体数增加而增加的趋势缓慢。红军亭样地虽与八角田样地同处猫儿山自然保护区, 但由于海拔低于八角田样地, 处于中海拔区域, 且地形复杂, 物种数随着取样面积增大, 上升趋势明显高于八角田样地。南岭样地和鼎湖山样地海拔差异较小, 但南岭样地物种数随取样个体数增大而增长的趋势最为明显, 甚至明显高于同在广东省但纬度比它更低的鼎湖山样地。

|
| 图 1 各样地的种-个体分布曲线 Fig. 1 The speicies-individual curve of each plots |
对各个样地的单个体物种的统计见表 1。纬度从低到高, 样地中的物种数呈现明显的差异, 单个体物种数所占比例呈现下降趋势。鼎湖山样地总个体数3608个, 物种95个, 其中单个体物种18个, 占到总物种数的18.9%。南岭样地总个体数最大为3326个, 分属165个物种, 其中单个体物种也最多达到28个, 占到总物种数的17%。猫儿山的八角田样地和红军亭样地地处同一纬度, 八角田样地的总个体数2654个, 物种81个, 其中单个体物种11个, 占到总物种数的13.6%;红军亭样地总个体数2471个, 共有物种89个, 其中单个体物种12个, 占到总物种数的13.5%。
| 样地 Plots |
生活型 Life form |
个体数 Number of individuals |
| 八角田样地 | 灌木 | 4 |
| 乔木 | 7 | |
| 红军亭样地 | 灌木 | 2 |
| 乔木 | 10 | |
| 南岭样地 | 灌木 | 6 |
| 乔木 | 22 | |
| 鼎湖山样地 | 灌木 | 3 |
| 乔木 | 15 |
以上结果验证了海拔的差异对于种-个体的分布起到关键作用, 个体数随海拔变化更加明显。而纬度从南到北的变化, 也使得单个体种的出现频率逐渐下降, 而单个体种对群落物种多样性的贡献也在低海拔地区更为显著。猫儿山两块样地的统计结果说明, 虽有海拔差异, 但单个体种在样地中所占的比例变化很小, 海拔差异导致的多样性变化主要来源于多个体物种的贡献。
3.2 优势种径级分布各样地的优势种按照重要值(IV)大小进行统计, 结果显示虽然都是南亚热带森林样地, 但各样地的物种多样性差异大, 最直观的表现是优势种不同。样地中重要值排名前十的优势种各不相同(表 2), 也从一个侧面证明了南亚热带区域物种多样性高。猫儿山的八角田样地最大优势种是南方铁杉(Tsuga chinensis var.tchekiangensis), 以大树维持优势, 重要值达到40.7, 但多度只有62;红军亭样地的最大优势种是水青冈(Fagus longipetiolata), 重要值达到50.3, 多度为340。南岭样地的显著优势种为仁昌厚壳桂(Cryptocarya chingii Cheng), 以多度维持优势, 多度为691, 重要值为35.3。鼎湖山样地中的第一优势种为云南银柴(Aporosa yunnanensis), 同样以多度维持优势, 多度高达897, 重要值36.9。重要值前十的物种多度占到样地总个体数的比例为鼎湖山样地(67.1%)>八角田样地(61.6%)>南岭样地(56%)>红军亭样地(54.1%)。
| 八角田样地 | 红军亭样地 | 南岭样地 | 鼎湖山样地 | |||||||||||
| 物种名 Species name |
多度 Abundance |
重要值 Important value |
物种名 Species name |
多度 Abundance |
重要值 Important value |
物种名 Species name |
多度 Abundance |
重要值 Important value |
物种名 Species name |
多度 Abundance |
重要值 Important value |
|||
| 南方铁杉 Tsuga chinensis var. tchekiangensis |
62 | 40.7 | 水青冈 Fagus longipetiolata |
340 | 50.3 | 仁昌厚壳桂 Cryptocarya chingii |
691 | 35.3 | 云南银柴 Aporusa yunnanensis |
879 | 36.9 | |||
| 黔稠 Cyclobalanopsis stewardiana |
420 | 37.1 | 榕叶冬青 Ilex ficoidea |
253 | 15.8 | 华润楠 Machilus chinensis |
169 | 19.3 | 锥栗 Castanopsis chinensis |
12 | 29.4 | |||
| 大八角 Illicium majus |
225 | 32.5 | 光叶柯 Lithocarpus mairei |
183 | 15.4 | 山羡叶刨花树 Meliosma thorelii |
293 | 17.7 | 荷木 Schima superba |
32 | 17.4 | |||
| 西南山茶 Camellia pitardii |
231 | 12.7 | 铁锥栲 Castanopsis lamontii |
61 | 13.1 | 网脉山龙眼 Helicia reticulate |
240 | 14.2 | 肖蒲桃 Acmena acuminatissima |
121 | 14.7 | |||
| 山桂花 Bennettiodendron leprosipes |
133 | 11.5 | 缺萼枫香 Liquidambar acalycina |
73 | 10.7 | 酸枣 Ziziphus jujuba |
61 | 11.2 | 光叶山黄皮 Aidia canthioides |
340 | 13.9 | |||
| 青冈 Cyclobalanopsis glauca |
187 | 11.3 | 桂南木莲 Manglietia chingii |
89 | 9.9 | 罗浮柿 Diospyros morrisana |
112 | 9.4 | 柏拉木 Blastus cochinchinensis |
346 | 13.5 | |||
| 资源木姜 Litsea pedunculata |
178 | 10.9 | 木姜叶润楠 Machilus litseifolia |
98 | 9.4 | 刨花楠 Machilus pauhoi |
48 | 9.3 | 鼎湖血桐 Macaranga sampsoni |
333 | 12.7 | |||
| 榕叶冬青 Ilex ficoidea |
80 | 8.4 | 交让木 Daphniphyllum macropodum |
91 | 9.2 | 藜蒴 Castanopsis fissa |
110 | 7.5 | 白颜树 Gironniera subaequalis |
90 | 12.3 | |||
| 凹脉杜鹃 Rhododendron davidsonianum |
76 | 8.1 | 拟榕叶冬青 Ilex subficoides |
103 | 8.6 | 米锥 Castanopsis carlesii |
59 | 7.5 | 九节 Psychotria asistica |
165 | 8.1 | |||
| 包石栎 Lithocarpus cleistocarpus |
44 | 7.7 | 包石栎 Lithocarpus cleistocarpus |
51 | 8.5 | 广东润楠 Machilus kwangtungensis |
81 | 7.0 | 红车 Syzygium rehderianum |
105 | 8.1 | |||
对各样地中的绝对优势种进行径级分布统计(图 2), 结果显示:八角田样地中的南方铁杉在样地中没有胸径小于10cm的个体, 虽然径级分布类似正态分布, 但大部分个体集中在15—35cm范围, 其在样地中的优势主要是大树在维持, 个体更新是急需关注的问题。红军亭中的水青冈和南岭样地的仁昌厚壳桂径级分布呈现明显的反“J”型分布, 表明目前种群处于一种良性发展趋势, 不存在明显的更新困难。鼎湖山样地的云南银柴也呈现类似反“J”型分布, 胸径10cm以下个体数量占绝对优势。

|
| 图 2 各样地重要值最大的优势种径级分布图 Fig. 2 The diameter at breast height (DBH) class distributions of most Ⅳ dominant species in each plots |
优势种的多度和径级分布情况从一个侧面能体现出群落的发展和演替状态。八角田样地的最大优势种南方铁山由于个体更新困难, 随着演替时间的延伸, 很有可能被次优势种取代优势地位。而其他三个群落中的第一优势种, 由于小个体居多, 将在相当长的演替过程中保持优势地位。
3.3 样地间α多样性比较各样地的物种多样性程度通过物种丰富度S、Shannon-Weiner指、Simpson指数和Pielou指数进行表征。结果显示(表 3):尽管南岭纬度高于鼎湖山, 但物种数却远多于鼎湖山样地, 处于同一纬度的红军亭样地的物种数略高于八角田样地。Shannon-Weiner指数由于是物种数和相对多度综合形成的指数, 因此也表现出南岭样地高于鼎湖山样地;而在红军亭样地地处中纬度也明显高于海拔偏高的八角田样地。受优势种影响较大的Simpson指数(也被称为优势度指数), 同样表现出南岭样地高于鼎湖山样地, 红军亭样地高于八角田样地。而表征物种个体数之间差异的物种均匀度Pielou指数也显示出:南岭样地高于鼎湖山样地, 红军亭样地高于八角田样地。
| 样地 Plots |
丰富度S Species richnessS |
Shannon-Weiner指数 Shannon-Weiner index |
Simpson指数 Simpson index |
Pielou指数 Pielou index |
| Ⅰ | 81 | 4.71 | 0.94 | 1.05 |
| Ⅱ | 89 | 5.27 | 0.96 | 1.17 |
| Ⅲ | 165 | 5.37 | 0.93 | 1.05 |
| Ⅳ | 95 | 4.42 | 0. 90 | 0.97 |
| Ⅰ.八角田样地BJT plot;Ⅱ.红军亭样地HJT plot;Ⅲ.南岭样地NL plot;Ⅳ.鼎湖山样地DH plot | ||||
由此可见, 综合比较几种α多样性指数, 他们表现出相同的趋势。海拔接近的南岭样地和鼎湖山多样性差异显著:虽然南岭样地纬度更高, 但多样性明显高于纬度更低的鼎湖山样地。而同一纬度不同海拔的八角田样地和红军亭样地相比较, 处于中纬度的红军亭样地α多样性高于海拔偏高的八角田样地。
3.4 样地间物种差异性比较Sorensen指数和Jaccard指数用来表征四个群落之间的因环境梯度不同, 不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率。统计结果一致表明(表 4):四个样地两两之间均存在明显的物种组成差异性。同纬度的八角田和红军亭样地的Sorensen指数和Jaccard指数最大, 说明它们之间的物种组成差异相比其他样地之间是最小的, 且差异主要是海拔和生境的不同造成的。猫儿山的两块样地与鼎湖山样地之间的Sorensen指数和Jaccard指数最小, 说明它们之间的物种组成差异相较于其他样地间是最大的, 这种差异不仅源于纬度相隔最远, 也源自于自于海拔相差1000m以上。海拔相近的南岭样地和鼎湖山样地之间Sorensen指数和Jaccard指数介于以上两者之间, 它们之间的物种组成差异相主要源于纬度和样地小生境的差异。
| 样地号 Plotss |
Sorensen指数 Sorensen index |
Jaccard指数 Jaccard index |
| Ⅰ和Ⅱ | 0.26 | 0.15 |
| Ⅰ和Ⅲ | 0.12 | 0.06 |
| Ⅰ和Ⅳ | 0.01 | 0.005 |
| Ⅱ和Ⅲ | 0.19 | 0.1 |
| Ⅱ和Ⅳ | 0.01 | 0.005 |
| Ⅲ和Ⅳ | 0.15 | 0.08 |
β多样性又称生境间的多样性。四个群落之间由于生境的差异, 物种组成明显不同。Sorensen指数和Jaccard指数检测出:纬度和海拔对群落β多样性都有影响。群落间纬度相差越大, 物种差异性越大。相同纬度的样地之间海拔对其β多样性的影响也是显著的。海拔相近的群落, 纬度不同也会造成物种组成差异性显著。
4 讨论与结论 4.1 海拔对亚热带植物群落物种多样性的影响种-个体数的分布规律在同一纬度上, 海拔不同分布规律不同。海拔的差异对于种-个体的分布起到关键作用, 个体数随海拔变化更加明显。在中亚热带, 海拔对山地植物群落形成起决定性作用[12]。有研究报道:植物叶片习性和温度气候调节的光合氮利用效率变化能够调节猫儿山常绿树种沿海拔形成双峰分布的机制[13]。这些也从另一个层面验证了群落组成和物种多样性随海拔变化的原因。文中猫儿山同一纬度不同海拔的八角田样地和红军亭样地相比较, 处于中纬度的红军亭样地α多样性高于海拔偏高的八角田样地。相同纬度的样地之间海拔对其β多样性的影响也是显著的。两个群落由于海拔差异, 物种丰富度和物种组成差异明显。单个体种在两个样地中所占的比例变化很小, 海拔差异导致的多样性变化主要来源于多个体物种的贡献。这些差异的来源基于两个群落生境差异性, 海拔通过水热状况影响植被类型及其分布[14], 以及海拔差异导致的植被带土壤微生物群落功能多样性差异显著[15]。
4.2 纬度对亚热带植物群落物种多样性的影响物种丰富度的纬度梯度分布格局, 即物种丰富度随纬度增加而减少, 是生态学以及生物地理学中最基本和最为广泛讨论的科学问题之一[16, 17]。文中群落在不同纬度的分布也具有随纬度降低, 物种数增加趋势越强的特点。而纬度从南到北的变化, 也使得单个体种的出现频率逐渐下降, 而单个体种对群落物种多样性的贡献也在低海拔地区更为显著。
两个低纬度群落(南岭和鼎湖山)的物种丰富度显著高于猫儿山两个群落。南岭样地和鼎湖山样地的海拔接近, 物种多样性差异仍然显著。并且南岭样地纬度更高, 但物种多样性反而明显高于纬度更低的鼎湖山样地。有悖于物种多样性随纬度变化的规律。前人的研究也发现位于南岭大顶山的常绿阔叶林群落较鼎湖山的季风常绿阔叶林具有更高的物种丰富度[18]。经过十多年的演替, 南岭大顶山常绿阔叶林群落依然保持较高的物种多样性水平, 常绿阔叶林比较年轻, 发育过程中结构和物种组成尚不稳定[19]。鼎湖山季风常绿阔叶林林龄较长, 在物种组成、结构方面比较均一。另外, 鼎湖山群落样地由于长期承担多项监测任务, 人为干扰较大, 因此统计数据结果低于其他几个群落。
β多样性是指不同群落间物种组成的差异[20], Jaccard和Sorensen指数广泛用来测定群落物种组成在时间或空间维度上的变化。同种森林群落类型内部及不同森林群落间的谱系β多样性存在显著差异, 反映了不同森林类型各自具有独特的物种和谱系组成。本研究发现四个群落之间β多样性的差异同样明显。四个群落间的物种组成差异, 随它们纬度之间的差异变化, 纬度对群落β多样性也有影响。群落间纬度相差越大, 物种差异性越大。海拔相近的群落, 纬度不同也会造成物种组成差异性显著。最近不少基于纬度梯度的研究都发现历史气候因素是决定β多样性分布格局的主要因子[20-22], 纬度变化往往通过物候条件反映出来, 从而影响一个区域的群落中的物种组成。
4.3 结论我国亚热带气候区植被资源丰富, 对其典型植物群落物种多样性分布规律探索是一项非常有意义的研究工作。文中通过一系列比较和分析, 发现四个均处于亚热带气候区的植物群落, 它们的物种多样性随海拔和纬度变化趋势明显。具体表现为:(1)海拔的差异对于种-个体的分布起到关键作用, 个体数随海拔变化更加明显。纬度从南到北单个体种的出现频率逐渐下降, 而单个体种对群落物种多样性的贡献也在低海拔地区更为显著。(2)海拔接近的南岭样地和鼎湖山样地α多样性差异显著:由于所处演替阶段不同, 以及鼎湖山样地受到人为干扰大, 物种多样性反而低于纬度较高的南岭样地。同一纬度不同海拔的猫儿山两块样地相比较, 处于中纬度的红军亭样地α多样性高于海拔偏高的八角田样地。(3)四个群落之间由于生境的差异, 物种组成明显不同。纬度和海拔对群落β多样性都有影响。群落间纬度相差越大, 物种差异性越大。相同纬度的样地之间海拔对其β多样性的影响也是显著的。海拔相近的群落, 纬度不同也会造成物种组成差异性显著。另外, 八角田样地的最大优势种南方铁山个体更新困难, 群落中现存都是大径级个体, 要引起关注。
| [1] |
胡一鸣, 梁健超, 金崑, 丁志锋, 周智鑫, 胡慧建, 蒋志刚. 喜马拉雅山哺乳动物物种多样性垂直分布格局. 生物多样性, 2018, 26(2): 191-201. |
| [2] |
孙影.森林群落物种多样性的纬度梯度性研究[D].厦门: 厦门大学, 2015. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2965138
|
| [3] |
吕巍.不同植被带森林群落木本植物多样性比较研究[D].北京: 北京林业大学, 2006. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y958760
|
| [4] |
宋永昌. 中国常绿阔叶林. 北京: 科学出版社, 2013: 23-24.
|
| [5] |
牟礼忠. 神奇的猫儿山. 广西林业, 1998(5): 38-39. |
| [6] |
段辉良, 曹福祥. 中国亚热带南岭山地气候变化特点及趋势. 中南林业科技大学学报, 2012, 32(9): 110-113. |
| [7] |
叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274-286. DOI:10.3773/j.issn.1005-264x.2008.02.005 |
| [8] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949: 121-122.
|
| [9] |
Simpson E H. Measurement of diversity. Nature, 1949, 163(4148): 688. DOI:10.1038/163688a0 |
| [10] |
Pielou E C.数学生态学引论.卢泽愚, 译.北京: 科学出版社, 1978: 33-37.
|
| [11] |
马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法Ⅱ β多样性的测度方法. 生物多样性, 1995, 3(1): 38-43. DOI:10.3321/j.issn:1005-0094.1995.01.007 |
| [12] |
段文军, 王金叶. 猫儿山自然保护区森林群落垂直格局与主导因素分析. 生态环境学报, 2013, 22(4): 563-566. DOI:10.3969/j.issn.1674-5906.2013.04.003 |
| [13] |
白坤栋, 蒋得斌, 万贤崇. 广西猫儿山不同海拔常绿树种和落叶树种光合速率与氮的关系. 生态学报, 2013, 33(16): 4930-4938. |
| [14] |
谭一波, 田红灯, 郑威, 王绍能. 广西猫儿山森林群落土壤物理性质空间变异及其环境影响因素. 广西林业科学, 2017, 46(2): 123-129. DOI:10.3969/j.issn.1006-1126.2017.02.001 |
| [15] |
宋贤冲, 郭丽梅, 田红灯, 邓小军, 赵连生, 曹继钊. 猫儿山不同海拔植被带土壤微生物群落功能多样性. 生态学报, 2017, 37(16): 5428-5435. |
| [16] |
Pianka E R. Latitudinal gradients in species diversity:a review of concepts. The American Naturalist, 1966, 100(910): 33-46. DOI:10.1086/282398 |
| [17] |
Willig M R, Kaufman D M, Stevens R D. Latitudinal gradients of biodiversity:pattern, process, scale, and synthesis. Annual Review of Ecology Evolution and Systematics, 2003, 34: 273-309. DOI:10.1146/annurev.ecolsys.34.012103.144032 |
| [18] |
彭华贵, 杜彦君, 李炯, 欧阳学军, 黄忠良. 南岭大顶山与鼎湖山常绿阔叶林种群分布格局对比研究. 生态环境, 2006, 15(4): 770-774. DOI:10.3969/j.issn.1674-5906.2006.04.023 |
| [19] |
毕肖峰, 彭华贵, 黄忠良, 曹洪麟. 南岭大顶山常绿阔叶林群落结构及其物种多样性. 生态科学, 2005, 24(2): 113-116. |
| [20] |
斯幸峰, 赵郁豪, 陈传武, 任鹏, 曾頔, 吴玲兵, 丁平. Beta多样性分解:方法、应用与展望. 生物多样性, 2017, 25(5): 464-480. |
| [21] |
王月霞, 金毅, 吴初平, 翁东明, 叶立新, 陈德良, 余建平, 刘金亮, 仲磊, 于明坚. 浙江省主要亚热带森林群落类型物种和谱系水平的α和β多样性比较. 生物多样性, 2016, 24(8): 863-874. |
| [22] |
Fernández L D, Fournier B, Rivera R, Lara E, Mitchell E A D, Hernández C E. Water-energy balance, past ecological perturbations and evolutionary constraints shape the latitudinal diversity gradient of soil testate amoebae in south-western South America. Global Ecology and Biogeography, 2016, 25(10): 1216-1227. DOI:10.1111/geb.12478 |