 2020, Vol. 40
2020, Vol. 40文章信息
- 纪娇娇, 郑蔚, 杨智杰, 王全成, 熊德成, 胥超, 杨玉盛
- JI Jiaojiao, ZHENG Wei, YANG Zhijie, WANG Quancheng, XIONG Decheng, XU Chao, YANG Yusheng
- 亚热带森林转换对土壤微生物呼吸及其熵值的影响
- Effects of subtropical forest conversion on soil microbial respiration and its entropy
- 生态学报. 2020, 40(3): 800-807
- Acta Ecologica Sinica. 2020, 40(3): 800-807
- http://dx.doi.org/10.5846/stxb201810142226
-
文章历史
- 收稿日期: 2018-10-14
- 网络出版日期: 2019-11-20






2. 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007
2. Cultivation Base of State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou 350007, China
土地利用变化是仅次于化石燃料向大气排放CO2的重要源[1-2], 其中87%源于人为驱动的森林转换, 是导致CO2浓度急剧增加的重要原因之一[3]。我国的亚热带地区是重要的商品林基地, 大面积的天然林被采伐转变为人工林[4], 导致土壤有机碳的数量和质量大幅度下降, 损失幅度高于全球平均水平[5-6]。同时, 森林转换引起的凋落物和细根归还、土壤环境等变化, 改变了土壤微生物群落结构、和活性, 对土壤碳循环产生重要影响[7-9]。因此, 探讨亚热带森林转换对土壤碳动态的影响具有重要的科学意义。
土壤微生物呼吸及其熵值(土壤代谢熵、微生物熵)是指示土壤碳代谢活性的重要参数, 是土壤质量的敏感性指标, 能够较早的揭示环境或生物因子改变对土壤的影响[10-11]。大量研究表明, 森林转换会给土壤环境带来显著变化从而影响微生物呼吸及其熵值[12-13]。Fang等[14]在温带长白山的研究发现, 天然林转换为白桦人工林和云杉人工林后, 0—10 cm土层土壤微生物生物量碳分别下降30%和41%, 而微生物熵和代谢熵没有显著变化。与温带地区不同, 亚热带地区经过森林转换后, 林分趋于单一, 群落结构变得简单[15], 土壤可溶性有机碳下降[16], 土壤微生物可利用基质含量下降, 严重影响微生物活性与微生物呼吸代谢过程。Yan等[17]在亚热带三峡库区的研究表明, 天然林转换为果树后, 0—20 cm土层土壤微生物呼吸下降了45%, 微生物熵仅为天然林的24%。目前关于亚热带森林转换后微生物呼吸熵值的研究较少且多集中在表层土壤。而对深层土壤缺乏关注。已有研究表明, 深层土壤(20—300 cm)碳储量约为表层土壤的3倍[18], 深层土壤碳动态的微小变化可能对对全球碳循环产生重要影响。在森林生态系统中, 枯死根系及凋落物经分解后的可溶性有机质会通过降水淋溶以及矿质土壤吸附等方式进入深层土壤并对深层土壤碳矿化及其储量产生影响[19-20]。因此深入研究森林转换对不同深度土壤微生物呼吸及其熵值的影响对维持和提高森林土壤质量以及进一步明确森林转换后土壤质量生态过程具有重要意义。
米槠是我国亚热带常绿阔叶林中的优势种, 伴随着南方商品林基地建设, 大量常绿阔叶林被转换为杉木、马尾松等速生造林树种[21-22]。因此本文以福建三明森林生态系统与全球变化定位观测研究站内的米槠天然林、杉木人工林和马尾松人工林的0—10 cm、10—20 cm、20—40 cm以及40—60 cm土层土壤为研究对象, 对天然林转换为人工林后土壤微生物呼吸及其熵值的变化进行研究, 探讨亚热带森林转换后对不同深度土壤质量的影响, 为深入了解森林转换对亚热带森林土壤碳动态的影响提供理论基础。
1 研究区与研究方法 1.1 研究区概况本研究试验样地设置在福建省三明森林生态系统与全球变化定位观测研究站-陈大观测点(26°19′N, 117°36′E), 该区东南接戴云山, 西北连武夷山脉, 主要地形为低山丘陵, 平均海拔330 m。属于亚热带季风气候区, 年均温约为19.1℃, 降水多集中在3—8月份, 年平均降水量为1750 mm, 该区域土壤主要为粗晶花岗岩发育形成的红壤。该区域主要树种为米槠(Castanopsis carlesii)、闽粤栲(Castanopsis fissa)木荷(Schima superba)等, 以米槠为优势树种。林下植被主要有毛冬青(Ilexpubesces)、狗骨柴(Diplospora dubia)、矩圆叶鼠刺(Itea oblonga)、沿海紫金牛(Ardisia punctate)、狗脊蕨(Woodwardia japonica)。
1.2 样地设置天然林为近200年无人为干扰的米槠林, 在米槠天然林的基础上于1976年对试验地进行火烧和皆伐, 人为种植杉木与马尾松。米槠天然林凋落物含量为8899.1 kg hm-2 a-1, 马尾松人工林和杉木人工林分别为6189.8 kg hm-2 a-1和5306.9 kg hm-2 a-1。分别在米槠天然林、杉木人工林与马尾松人工林的上、中、下坡位各设置1个20 m×20 m试验样方, 共9个试验样方, 同一林分上、中、下坡位的样方为试验重复。
1.3 土壤样品的采集与处理2017年7月, 在实验样地采用S型法选取采样点, 去除地表凋落物后利用土钻法在9个样方分别采集0—10 cm、10—20 cm、20—40 cm、40—60 cm土壤样品, 共计36个土样。所有土样均立即放入保温箱内并立即带回实验室处理。土样去除杂质、根系和凋落物等, 分成两份, 一份过2 mm筛网后于4℃冰箱冷藏保存, 用于土壤微生物生物量碳、微生物呼吸速率的测定。另一份在一周内自然风干并过0.45 μm筛网后进行土壤总有机碳、总氮等理化性质的测定。
1.4 土壤样品的测定土壤总有机碳、总氮采用碳氮元素分析仪(Elemental EL MAX CNS analyzer, 德国)进行测定。该研究区土壤样品均为酸性土壤, 无机碳含量极少, 元素分析法通过高温燃烧的杜马斯法[23]测定有机碳。
采用重量法测定土壤含水率。用0.1 g感量的天平, 称取土样的质量, 记录鲜土土样的质量mt;将土样放置105℃的烘箱烘干至衡重后用天平秤得干土的重量ms, 根据以下公式进行进一步的计算:
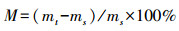
|
(1) |
式中, M指样品含水率, %。
微生物呼吸速率采用室内培养法进行[24]。本实验每个处理设置3个平行, 称取相当于干土10 g的鲜土, 放置于培养瓶中, 保鲜膜封口并扎孔以便于培养瓶内土壤透气, 在22℃的无照明培养瓶预培养10 h, 使其适应培养环境后, 将培养瓶取出来进行洗气后用Li—840测定CO2浓度作为初始值, 然后将培养瓶放回培养箱8 h后用Li—840再次测定CO2浓度。
采用底物诱导呼吸法(SIR)测定土壤微生物生物量碳[25]。称取相当于干重10 g的鲜土, 放置于培养瓶中, 用保鲜膜封口并扎孔便于培养瓶内土壤透气。将培养瓶放置于22℃的无照明培养箱中培养一周, 在培养至第4天时, 对培养瓶进行洗气。培养结束后, 对培养瓶进行洗气, 然后加入800 μg/g的葡萄糖溶液并保证培养瓶内土壤水量不超过田间持水量, 之后将培养瓶放置于培养箱1 h使土壤呼吸稳定后用Li-840测定CO2浓度作为初始值, 然后将培养瓶放回培养箱培养两个小时后再次测定CO2浓度。
1.5 数据处理与分析土壤微生物呼吸速率计算公式[26]:
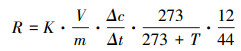
|
(2) |
式中, R为土壤微生物CO2呼吸速率(μg C g-1 h-1);K为气体转换常数(g/m3), Δc/Δt为单位时间内CO2浓度的变化量(μmol mol-1 h-1);V为培养瓶内总气体(m3);m为干土重(g);T为培养温度(℃);12/44为CO2中C的占比。
土壤微生物生物量碳(MBC)含量计算公式:
土壤MBC的计算以SIR方法为理论基础[27], 采用公式计算土壤MBC(mg/kg):
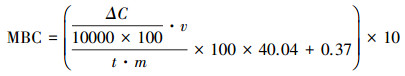
|
(3) |
式中, ΔC表示土壤培养前后CO2浓度变化量(μmol mol-1 h-1);V表示培养瓶内总气体体积(m3);t表示培养时间(h);m表示干土重(g)。
土壤微生物熵(mg/g)计算公式[11]:
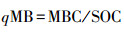
|
(4) |
土壤代谢熵(h-1)计算公式[11]:

|
(5) |
式中, CO2-C为培养8 h内微生物呼吸排放出的碳(μmol/mol)。
采用SPSS 20.0软件进行统计分析。采用单因素方差分析(one-way ANOVA)进行显著性分析, 并通过最小显著差异法(LSD)比较不同数据组间的差异(α=0.05)。
2 结果与分析 2.1 森林转换对土壤微生物呼吸的影响各林分土壤微生物呼吸速率均随土壤深度的增加而下降(图 1)。森林转换后, 马尾松人工林土壤微生物呼吸速率比米槠天然林高32%(P<0.05), 而杉木人工林与米槠天然林之间没有显著差异。与米槠天然林相比, 马尾松人工林和杉木人工林10—20 cm土壤微生物呼吸速率分别下降26%和24%(P<0.05)。但在20—40 cm和40—60 cm土层, 杉木人工林与米槠天然林的土壤微生物呼吸速率无显著差异, 而马尾松人工林土壤微生物呼吸速率显著低于米槠天然林。

|
| 图 1 不同林分不同土层土壤微生物呼吸速率 Fig. 1 Soil microbial respiration rate of different soil layers in different stands (平均值±标准误), n=3;不同大写字母表示同一土层不同林分土壤微生物呼吸速率差异显著(P<0.05);不同小写字母表示同一林分不同土层土壤微生物呼吸速率差异显著(P<0.05) |
各林分土壤MBC随土壤深度增加而降低(图 2)。马尾松人工林和杉木人工林0—10 cm土壤MBC分别比米槠天然林下降19%和40%(P<0.05)。杉木人工林10—20 cm土层MBC含量与米槠天然林没有显著差异, 而马尾松人工林10—20 cm土壤MBC含量比米槠天然林高29%(P<0.05)。但是, 各林分间20—60 cm土壤MBC的变化并不显著。

|
| 图 2 不同林分不同土层土壤微生物生物量碳 Fig. 2 Soil microbial biomass carbon in different soil layers of different forest stands (平均值±标准误), n=3;不同大写字母表示同一土层不同林分土壤MBC差异显著(P<0.05);不同小写字母表示同一林分不同土层土壤MBC差异显著(P<0.05) |
米槠天然林与杉木人工林土壤微生物熵随土壤深度的增加而增加, 马尾松人工林土壤微生物熵随土层增加而降低(图 3)。森林转换并没有显著影响0—10 cm土壤微生物熵, 土壤微生物熵的变化主要发生在深层土壤。在20—40 cm与40—60 cm土层, 杉木人工林土壤微生物熵与米槠天然林相比分别下降了51%、52%, 而马尾松人工林分别下降71%、66%(P<0.05)。但是, 在10—20 cm土层, 马尾松人工林土壤微生物熵比米槠天然林高71%(P<0.05)。

|
| 图 3 不同林分不同土层土壤微生物熵 Fig. 3 Microbial quotient of soil in different soil layers of different forest stands (平均值±标准误), n=3;不同大写字母表示同一土层不同林分土壤微生物熵差异显著(P<0.05);不同小写字母表示同一林分不同土层土壤微生物熵差异显著(P<0.05) |
米槠天然林、杉木人工林和马尾松人工林表层土壤(0—10 cm)微生物代谢熵均显著高于深层土壤(20—60 cm)(P<0.05)(图 4)。森林转换后, 马尾松人工林和杉木人工林0—10 cm土壤微生物代谢熵比米槠天然林分别增加了38%和29%(P<0.05)。但是, 在10—20 cm土层, 马尾松人工林土壤微生物代谢熵比米槠天然林下降50%(P<0.05), 而各林分在20—40 cm和40—60 cm的土壤微生物代谢熵均没有显著差异。

|
| 图 4 不同林分不同土层微生物代谢熵 Fig. 4 Metabolic entropy of microorganism in different soil layers of different forest stands (平均值±标准误), n=3;不同大写字母表示同一土层不同林分土壤微生物代谢熵差异显著(P<0.05);不同小写字母表示同一林分不同土层土壤微生物代谢熵差异显著(P<0.05) |
土壤微生物呼吸是土壤呼吸的重要组分, 占土壤总呼吸的50%左右, 主要反映了土壤微生物对土壤有机质的利用, 代表着土壤微生物活性和土壤物质代谢的强度[28-29]。森林转换后, 人工林0—10 cm土壤微生物呼吸比天然林显著上升(图 1)。可能的原因是人工林表层土壤经过整地、除草等营林措施, 受人为干扰程度大, 土壤团聚体结构受到破坏, 加剧了微生物对有机质的分解[30]。李英等[31]在黄淮海平原的也研究发现, 长期受到人为干扰的耕地与未受干扰的林地、草地相比表现出更高的微生物呼吸速率。另一方面可能是因为森林转换后, 人工林林冠疏开, 阳光直射使表层土壤温度升高, 提高土壤微生物活性并促进有机质的分解[26]。多数研究也表明, 土壤微生物呼吸与土壤温度呈指数相关, 且针叶林土壤呼吸的温度敏感性会显著高于阔叶林[32]。有研究表明土壤微生物呼吸速率随着土壤有机质稳定性的增加而减少[33]。天然林表层土壤有机碳含量高而呼吸速率小, 人工林有机碳含量低而呼吸速率高这表明在天然林被改为人工林的过程中, 土壤有机碳库的固碳能力在减小[34]。另外, 也有研究表明, 天然林转换为人工林后, 活细根和死细根生物量、细根周转速率、年均净生产量均呈现下降趋势[35], 这可能是造成10—60 cm人工林土壤微生物呼吸速率降低的原因(图 1)。3片林分土壤微生物呼吸速率均随土壤深度增加而降低, 这与土壤有机碳储量随土壤深度降低相符合(表 1), 土壤深度增加, 土壤有机碳含量下降, 微生物可利用底物减少, 微生物呼吸速率随之降低。
| 林型 Stand type |
土壤深度 Soil depth/cm |
总有机碳 Soil organic C/(g/kg) |
总氮 Total N/ (g/kg) |
细根生物量 Fine root biomass/ (g/m2) |
可溶性有机碳 Dissolved organic carbon/ (mg/kg) |
可溶性有机氮 Dissolved organic Nitrogen/ (mg/kg) |
含水率 Moisture content/% |
酸碱度 pH |
| 米槠天然林 | 0—10 | 50.10±2.44Aa | 2.74±0.10Aa | 191.05±23.52Aa | 164.00±34.28Aa | 3.61±1.09Ab | 42.00±2.71Aa | 4.08±0.1Ab |
| Castanopsis carlessi | 10—20 | 26.97±2.36Ba | 1.47±0.04Ba | 63.30±7.36Ba | 24.00±2.95Ba | 0.72±0.12Ba | 30.00±1.02Ba | 4.10±0.08Ba |
| natural forest | 20—40 | 15.58±1.83Ca | 1.54±0.05Ba | 44.27±5.65Ca | 20.00±3.2Ba | 0.49±0.07Ca | 28.00±0.81Ba | 3.78±0.06Ba |
| 40—60 | 8.38±0.90Da | 1.22±0.10Ba | 29.51±2.79Ca | 18.00±4.63Ba | 0.28±0.06Da | 12.00±1.95Ca | 3.65±0.08Bb | |
| 杉木人工林 | 0—10 | 20.86±1.28Ab | 1.28±0.07Ab | 122.71±15.31Ac | 122.96±18.25Ab | 3.70±0.21Aa | 32.00±0.39Ab | 4.52±0.08Aa |
| Cunninghamia lanceolata | 10—20 | 11.70±1.21Bb | 0.85±0.09Bb | 103.68±12.13Bb | 28.00±2.55Ba | 0.72±0.16Ba | 25.00±1.13Bb | 4.35±0.04Aa |
| Lamb. plantation | 20—40 | 9.67±0.45Bb | 0.63±0.02Bb | 98.25±10.47Bb | 22.00±5.62Ba | 0.35±0.09CB | 23.00±2.86Bb | 4.00±0.1Aa |
| 40—60 | 6.23±1.10Ca | 0.62±0.06Bb | 68.73±8.19Cb | 15.00±2.11Cb | 0.20±0.05Db | 12.00±1.90Ca | 3.60±0.08Bb | |
| 马尾松人工林 | 0—10 | 15.51±1.30Ac | 1.13±0.09Ab | 150.09±17.35Ab | 34.34±2.98Ab | 1.18±0.14Ba | 35.00±2.92Ab | 4.47±0.04Aa |
| Pinusmassoniana | 10—20 | 7.84±0.71Bc | 0.75±0.20Bb | 41.68±5.22Ba | 10.78±0.85Bb | 0.51±0.09Bb | 31.00±1.39Aa | 4.39±0.05Aa |
| Lamb. plantation | 20—40 | 5.50±0.68Bc | 1.33±0.05Aa | 25.24±3.10Cc | 8.40±0.03Bb | 0.19±0.03Cb | 18.00±0.95Bb | 3.92±0.04Ba |
| 40—60 | 4.02±0.22Bb | 0.98±0.03Aa | 20.97±5.30Ca | 7.90±0.46Bc | 0.12±0.04Cb | 12.00±1.10Ba | 3.85±0.06Ba | |
| 注:表中数据为平均值±标准误; 不同大写字母表示同林分不同土层差异显著(P<0.05), 不同小写字母表示同土层不同林分差异显著(P<0.05) | ||||||||
MBC在土壤全碳中占比很小, 但其周转时间快, 是土壤有机质中的活性部分, 对环境变化十分敏感[36-37]。本研究中, 马尾松人工林和杉木人工林0—10 cm与20—60 cm土壤MBC显著低于米槠天然林(图 2), 可能是由于森林转换导致凋落物数量和输入量减少的原因。凋落物分解是森林生态系统中养分归还的主要途径, 是土壤微生物的重要养分来源[38]。天然林凋落物含量为8899.1 kg hm-2 a-1, 分别比马尾松人工林和杉木人工林高43%和67%。另一方面可能是因为马尾松人工林和杉木人工林凋落物多为针叶, 相比天然林, 针叶林凋落物分解速率低, 进而影响微生物的生长与繁殖[39]。相关研究表明MBC与土壤含水量呈显著正相关[40-41]。本研究中, 森林转换后, 马尾松人工林10—20 cm土壤MBC显著高于米槠天然林和杉木人工林(图 2), 可能与马尾松人工林该土层土壤含水率较高有关(表 1)。本研究区地处中亚热带地区, 季风盛行, 湿润多雨, 年平均降雨量达1750 mm, 且强降雨较多[42], 森林转换后, 马尾松人工林凋落物含量少, 表层土壤覆盖少, 0—10 cm土壤养分易随雨水下渗至10—20 cm土层, 促进了微生物的生长和繁殖。
本研究中, 土壤微生物熵的变化比MBC更明显, 表明微生物熵对森林土壤质量变化更为敏感, 这与Saggar等[43]对新西兰耕地的研究结果相一致。微生物熵代表着土壤有机碳可以利用性, 可以表征外界环境变化对土壤有机碳矿化分解的影响[44-45], 是指示土壤质量的重要指标之一。本研究中, 森林转换后, 3种林分0—20 cm土壤微生物熵没有显著差异, 而人工林20—60 cm土壤微生物熵显著低于天然林。这表明森林转换导致人工林深层土壤可利用有机碳含量减少, 土壤质量下降。另外, 米槠天然林与杉木人工林土壤微生物熵均表现为深层土壤显著高于表层土壤, 而马尾松人工林各土层微生物熵无明显规律。这与张于光等[46]在川西亚高山森林对原始冷杉林转换为云杉人工林及农业用地的研究发现类似。不同于马尾松人工林, 杉木人工林微生物熵与米槠天然林呈现相同的变化趋势, 表明杉木人工林土壤有机碳库可利用性逐渐恢复到造林前水平, 王彦梅等[47]在长白山对30年生、45年生红松人工林的研究也表明, 造林45年后人工林土壤有机储量达到36.11 g/kg, 与天然林无显著差异, 土壤质量恢复到造林前水平。
微生物代谢熵反映了土壤微生物群落维持生存所需能量的大小及其对土壤基质的利用效率[48], 它将土壤微生物生物量碳的含量与微生物群落整体活性有机地结合起来[49-50]。微生物代谢熵较小, 表明微生物维持相同的生物量所消耗的能量较少, 释放CO2少, 微生物体周转率快[44]。本研究中, 在0—10 cm土壤, 在森林转换后人工林微生物代谢熵显著高于天然林(图 4)。表明人工林土壤微生物呼吸消耗的碳占微生物总碳的比例大[51], 代谢效率降低。这可能是由于森林转换后, 人工林表层土壤细菌含量升高, 细菌产生较少的生物量, 表现出更高的呼吸速率, 引起代谢熵的上升[52]。另外, 本研究中, 马尾松人工林10—20 cm土壤代谢熵显著低于天然林和杉木人工林, 主要是由于马尾松人工林该土层微生物生物量碳含量较高, 该土层养分充足, 微生物生存环境较好, 碳利用效率高。除此之外, 3种林分深层土壤代谢熵没有显著差异, 但各林分深层土壤代谢熵均显著低于表层土壤, 这可能是由于深层土壤可利用有机质较少, 微生物需保持较高的碳利用效率维持自身生存[51]。
4 结论我国亚热带森林转换现象普遍, 其对土壤的影响一直是人们的关注热点, 本文旨在揭示土壤微生物对森林转换的响应。(1)森林转换对土壤微生物呼吸及代谢熵产生显著影响。人工林表层土壤微生物呼吸速率降低(P<0.05), 代谢熵增高, 与天然林相比, 森林转换降低了表层土壤微生物碳利用效率, 这加重了人工林表层土壤对大气CO2的贡献量。(2)森林转换后, 人工林深层土壤MBC含量虽无显著变化, 但微生物熵成倍下降(P<0.05), 这表明森林转换使人工林深层土壤相比天然林微生物碳库占比下降, 固碳潜力降低。(3)天然林转换为马尾松人工林与转换为杉木林相比, 土壤质量恢复较慢。因此, 需要深入研究我国亚热带地区森林转换对不同深度森林土壤质量、养分恢复的影响及其微生物响应机制。
| [1] |
Carlson K M, Curran L M, Asner G P, Pittman A M, Trigg S N, Adeney J M. Carbon emissions from forest conversion by Kalimantan oil palm plantations. Nature Climate Change, 2013, 3(3): 283-287. DOI:10.1038/nclimate1702 |
| [2] |
Sharma S, Pandey D, Agrawal M. Global warming potential and sustainable management of three land uses in Varanasi. Management of Environmental Quality:An International Journal, 2016, 27(4): 364-373. DOI:10.1108/MEQ-05-2014-0078 |
| [3] |
Lü M K, Xie J S, Wang C, Guo J F, Wang M H, Liu X F, Chen Y M, Chen G S, Yang Y S. Forest conversion stimulated deep soil C losses and decreased C recalcitrance through priming effect in subtropical China. Biology and Fertility of Soils, 2015, 51(7): 857-867. DOI:10.1007/s00374-015-1035-y |
| [4] |
杨玉盛, 陈光水, 王小国, 谢锦升, 高人, 李震, 金钊. 中国亚热带森林转换对土壤呼吸动态及通量的影响. 生态学报, 2005, 25(7): 1684-1690. DOI:10.3321/j.issn:1000-0933.2005.07.022 |
| [5] |
杨玉盛, 谢锦升, 盛浩, 陈光水, 李旭. 中亚热带山区土地利用变化对土壤有机碳储量和质量的影响. 地理学报, 2007, 62(11): 1123-1131. |
| [6] |
Guo L B, Gifford R M. Soil carbon stocks and land use change:a meta analysis. Global Change Biology, 2002, 8(4): 345-360. DOI:10.1046/j.1354-1013.2002.00486.x |
| [7] |
何芳兰, 徐先英, 尉秋实, 刘虎俊, 贺访印, 马俊梅, 金红喜. 祁连山青海云杉人工林与天然林群落结构特征及物种多样性比较研究. 西北林学院学报, 2016, 31(5): 1-7. DOI:10.3969/j.issn.1001-7461.2016.05.01 |
| [8] |
洪丕征, 刘世荣, 王晖, 于浩龙. 南亚热带红椎和格木人工幼龄林土壤微生物群落结构特征. 生态学报, 2016, 36(14): 4496-4508. |
| [9] |
邱甜甜, 刘国彬, 王国梁, 孙利鹏, 姚旭. 黄土高原不同生长阶段油松人工林土壤微生物生物量碳的变化及其影响因素. 应用生态学报, 2016, 27(3): 681-687. |
| [10] |
杨宁, 邹冬生, 杨满元, 雷玉兰, 林仲桂, 付美云, 宋光桃. 紫色土丘陵坡地植被恢复过程中土壤微生物生物量碳、微生物熵的变化. 水土保持通报, 2014, 34(5): 39-43. |
| [11] |
吴东梅, 郭剑芬, 张政, 李帅军, 杨玉盛. DOM对米槠次生林不同土层土壤微生物呼吸及其熵值的影响. 生态学报, 2018, 38(11): 3806-3815. |
| [12] |
彭艳, 李心清, 程建中, 邢英, 闫慧. 贵阳喀斯特地区植被类型与季节变化对土壤微生物生物量和微生物呼吸的影响. 地球化学, 2010, 39(3): 266-273. |
| [13] |
Pabst H, Gerschlauer F, Kiese R, Kuzyakov Y. Land use and precipitation affect organic and microbial carbon stocks and the specific metabolic quotient in soils of eleven ecosystems of Mt. Kilimanjaro, Tanzania. Land Degradation & Development, 2016, 27(3): 592-602. |
| [14] |
Fang X M, Yu D P, Zhou W M, Zhou L, Dai L M. The effects of forest type on soil microbial activity in Changbai Mountain, Northeast China. Annals of Forest Science, 2016, 73(2): 473-482. DOI:10.1007/s13595-016-0540-y |
| [15] |
冯建新, 高伟民, 邓飞, 胡双成, 陈光水. 亚热带天然常绿阔叶林转变为杉木人工林对土壤微生物呼吸的影响. 亚热带资源与环境学报, 2015, 10(2): 16-24. DOI:10.3969/j.issn.1673-7105.2015.02.003 |
| [16] |
元晓春, 林伟盛, 蒲晓婷, 杨智榕, 郑蔚, 陈岳民, 杨玉盛. 更新方式对亚热带森林土壤溶液可溶性有机质数量及化学结构的影响. 应用生态学报, 2016, 27(6): 1845-1852. |
| [17] |
Yan T M, Yang L Z, Campbell C D. Microbial biomass and metabolic quotient of soils under different land use in the Three Gorges Reservoir area. Geoderma, 2003, 115(1/2): 129-138. |
| [18] |
Fontaine S, Barot S, Barré P, Bdioui N, Mary B, Rumpel C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature, 2007, 450(7167): 277-280. DOI:10.1038/nature06275 |
| [19] |
Bengtson P, Barker J, Grayston S J. Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects. Ecology and Evolution, 2012, 2(8): 1843-1852. DOI:10.1002/ece3.311 |
| [20] |
Uselman S M, Qualls R G, Lilienfein J. Quality of soluble organic C, N, and P produced by different types and species of litter:root litter versus leaf litter. Soil Biology and Biochemistry, 2012, 54: 57-67. DOI:10.1016/j.soilbio.2012.03.021 |
| [21] |
孙长忠, 沈国舫. 我国主要树种人工林生产力现状潜力的调查研究:Ⅰ.杉木、马尾松人工林生产. 林业科学研究, 2000, 13(6): 613-621. DOI:10.3321/j.issn:1001-1498.2000.06.007 |
| [22] |
胡砚秋, 李文斌, 崔佳玉, 苏志尧. 亚热带常绿阔叶林优势种个体及生物量的点格局分析. 生态学报, 2016, 36(4): 1066-1072. |
| [23] |
王巧环, 任玉芬, 孟龄, 李虹, 傅慧敏, 王华锋. 元素分析仪同时测定土壤中全氮和有机碳. 分析试验室, 2013, 32(10): 41-45. |
| [24] |
Nakamoto T, Wakahara S. Development of substrate induced respiration (SIR) method combined with selective inhibition for estimating fungal andbacterial biomass in humic andosols. Plant Production Science, 2004, 7(1): 70-76. DOI:10.1626/pps.7.70 |
| [25] |
Tateishi T. Estimation of fungal to bacterial biomass ratio in a burned pine forest soil by substrate-induced respiration inhibition method and microscopic method. Soil Microorganisms, 2000, 54(1): 1-11. |
| [26] |
林杉, 陈涛, 赵劲松, 向荣彪, 胡荣桂, 张水清, 王米兰, 吕昭琪. 不同培养温度下长期施肥水稻土的有机碳矿化特征. 应用生态学报, 2014, 25(5): 1340-1348. |
| [27] |
Carter M R. Soil Sampling and Methods of Analysis. Boca Raton:Lewis Publishers, 1993, 15-24. |
| [28] |
Wang Q, He N P, Yu G R, Gao Y, Wen X F, Wang R F, Koerner S E, Yu Q. Soil microbial respiration rate and temperature sensitivity along a North-South forest transect in eastern China:patterns and influencing factors. Journal of Geophysical Research:Biogeosciences, 2016, 121(2): 399-410. DOI:10.1002/2015JG003217 |
| [29] |
Yazdanpanah N, Mahmoodabadi M, Cerdà A. The impact of organic amendments on soil hydrology, structure and microbial respiration in semiarid lands. Geoderma, 2016, 266: 58-65. DOI:10.1016/j.geoderma.2015.11.032 |
| [30] |
王小红, 杨智杰, 刘小飞, 杨玉盛, 吴君君, 付林池, 陈坦, 李伟. 天然林转换成人工林对土壤团聚体稳定性及有机碳分布的影响. 水土保持学报, 2014, 28(6): 177-182, 189-189. |
| [31] |
李英, 韩红艳, 王文娟, 杨光菲, 赵灿灿. 黄淮海平原不同土地利用方式对土壤有机碳及微生物呼吸的影响. 生态环境学报, 2017, 26(1): 62-66. |
| [32] |
罗璐, 申国珍, 谢宗强, 周利光. 神农架海拔梯度上4种典型森林的土壤呼吸组分及其对温度的敏感性. 植物生态学报, 2011, 35(7): 722-730. |
| [33] |
Sonnenholzner S, Boyd C E. Vertical gradients of organic matter concentration and respiration rate in pond bottom soils. Journal of the World Aquaculture Society, 2000, 31(3): 376-380. |
| [34] |
戴慧, 王希华, 阎恩荣. 浙江天童土地利用方式对土壤有机碳矿化的影响. 生态学杂志, 2007, 26(7): 1021-1026. DOI:10.3321/j.issn:1000-4890.2007.07.010 |
| [35] |
杨玉盛, 陈光水, 林鹏, 黄荣珍, 陈银秀, 何宗明. 格氏栲天然林与人工林细根生物量、季节动态及净生产力. 生态学报, 2003, 23(9): 1719-1730. DOI:10.3321/j.issn:1000-0933.2003.09.004 |
| [36] |
李国辉, 陈庆芳, 黄懿梅, 安韶山, 郑粉莉, 陈利顶. 黄土高原典型植物根际对土壤微生物生物量碳、氮、磷和基础呼吸的影响. 生态学报, 2010, 30(4): 976-983. |
| [37] |
郭振, 王小利, 徐虎, 段建军, 张雅蓉, 李渝, 蒋太明. 长期施用有机肥增加黄壤稻田土壤微生物量碳氮. 植物营养与肥料学报, 2017, 23(5): 1168-1174. |
| [38] |
张明锦, 陈良华, 张健, 杨万勤, 刘华, 李勋, 张艳. 马尾松人工林林窗内凋落叶微生物生物量碳和氮的动态变化. 应用生态学报, 2016, 27(3): 672-680. |
| [39] |
杨晨, 刘勇, 陈晓, 王巍伟. 油松人工林下真菌群落对凋落物分解的影响. 中南林业科技大学学报, 2016, 36(7): 41-47. |
| [40] |
刘爽, 王传宽. 五种温带森林土壤微生物生物量碳氮的时空格局. 生态学报, 2010, 30(12): 3135-3143. |
| [41] |
赵彤, 闫浩, 蒋跃利, 黄懿梅, 安韶山. 黄土丘陵区植被类型对土壤微生物量碳氮磷的影响. 生态学报, 2013, 33(18): 5615-5622. |
| [42] |
万菁娟, 郭剑芬, 纪淑蓉, 任卫岭, 杨玉盛. 可溶性有机物输入对亚热带森林土壤CO2排放及微生物群落的影响. 林业科学, 2016, 52(2): 106-113. |
| [43] |
Saggar S, Yeates G W, Shepherd T G. Cultivation effects on soil biological properties, microfauna and organic matter dynamics in Eutric Gleysol and Gleyic Luvisol soils in New Zealand. Soil and Tillage Research, 2001, 58(1/2): 55-68. |
| [44] |
刘珊珊, 王芬, 张兴华, 宫渊波, 王燕, 尹艳杰, 李渊, 马金松, 郭挺. 放牧干扰对岷江上游山地森林-干旱河谷交错带土壤微生物量及呼吸熵的影响. 水土保持通报, 2014, 34(2): 63-68. |
| [45] |
立天宇, 康峰峰, 韩海荣, 高晶, 宋小帅. 冀北辽河源油松天然林土壤微生物碳代谢特征研究. 土壤, 2015, 47(3): 550-557. |
| [46] |
张于光, 张小全, 肖烨. 米亚罗林区土地利用变化对土壤有机碳和微生物量碳的影响. 应用生态学报, 2006, 17(11): 2029-2033. DOI:10.3321/j.issn:1001-9332.2006.11.007 |
| [47] |
王彦梅, 王朋, 于立忠. 辽东山区天然次生林转化为人工林对土壤有机碳的影响. 东北林业大学学报, 2010, 38(12): 54-57. DOI:10.3969/j.issn.1000-5382.2010.12.018 |
| [48] |
Babujia L C, Hungria M, Franchini J C, Brookes P C. Microbial biomass and activity at various soil depths in a Brazilian oxisol after two decades of no-tillage and conventional tillage. Soil Biology and Biochemistry, 2010, 42(12): 2174-2181. DOI:10.1016/j.soilbio.2010.08.013 |
| [49] |
Lu S, Wang Q, Katahata S, Naramoto M, Mizunaga H. Soil microbial activities in beech forests under natural incubation conditions as affected by global warming. Pedosphere, 2014, 24(6): 709-721. DOI:10.1016/S1002-0160(14)60058-8 |
| [50] |
张亮亮, 罗明, 韩剑, 何贵伦, 徐金虹. 南疆枣树-棉花间作对土壤微生物区系及代谢熵的影响. 棉花学报, 2016, 28(5): 493-503. |
| [51] |
周焱, 徐宪根, 王丰, 阮宏华, 汪家社, 方燕鸿, 吴焰玉, 徐自坤. 武夷山不同海拔梯度土壤微生物生物量微生物呼吸及其商值(qMB, qCO2). 生态学杂志, 2009, 28(2): 265-269. |
| [52] |
Shihan A, Hättenschwiler S, Milcu A, Joly F X, Santonja M, Fromin N. Changes in soil microbial substrate utilization in response to altered litter diversity and precipitation in a Mediterranean shrubland. Biology and Fertility of Soils, 2017, 53(2): 171-185. DOI:10.1007/s00374-016-1166-9 |