 2020, Vol. 40
2020, Vol. 40文章信息
- 叶柳欣, 库伟鹏, 刘军, 徐旻昱, 孟方荣, 傅伟军, 刘娟, 金锦, 吴家森
- YE Liuxin, KU Weipeng, LIU Jun, XU Minyu, MENG Fangrong, FU Weijun, LIU Juan, JIN Jin, WU Jiasen
- 封育年限对毛竹林群落结构和林下植物多样性的影响
- Effects of exclosure term on community structure and undergrowth diversity of Phyllostachys pubescens
- 生态学报. 2020, 40(3): 921-930
- Acta Ecologica Sinica. 2020, 40(3): 921-930
- http://dx.doi.org/10.5846/stxb201809282120
-
文章历史
- 收稿日期: 2018-09-28
- 网络出版日期: 2019-11-20






2. 浙江农林大学环境与资源学院, 临安 311300;
3. 杭州市余杭区林业工作站, 余杭 311100;
4. 杭州格真农产品专业合作社, 余杭 311118
2. School of Environment and Resource, Zhejiang Agriculture and Forestry University, Lin'an 311300, China;
3. Forestry Workstations in Yuhang District of Hangzhou, Yuhang 311100, China;
4. Ge Zhen Professional Cooperative of Agricultural Products in Hangzhou, Yuhang 311118, China
森林群落的林分密度、乔木树种的胸径分布和林下植物多样性反映了森林群落的形成、稳定性及演替规律, 影响着森林生态功能的发挥和立地生产力的可持续性[1-3]。不同林龄人工林林下植物种类多样性预示着生态系统的稳定性, 标志着人工林演替的方向和进程[4]。
毛竹是我国重要的森林资源, 广泛分布于我国南方的浙江、福建、江西、湖南等省份, 面积达443×104 hm2, 具有生长快、产量高、用途广、可持续性强等特点[5-6], 2016年, 浙江省毛竹林面积80.71×104 hm2, 总株数297888万株[7]。毛竹林群落结构及其稳定性受到人为活动的强烈支配和制约, 表现为不同干扰程度的毛竹林乔木层结构和林下植物多样性有显著差异。研究表明, 材用毛竹林胸径大于同等管理措施下的笋用林[8];长期经营后毛竹林群落结构简化, 林下灌木多样性降低[9-10];随着人为经营强度的增大, 毛竹林下植物的多样性、丰富度指数均随之减少[11-12];也有研究认为人为经营后毛竹林下灌木、草本的物种多样性上升, 草本层多样性指数、均匀度指数及物种数均以重度干扰的毛竹林为最高[13]。
随着我国社会、经济的发展, 毛竹价格逐年下降[14], 劳动力成本持续上升[15], 导致毛竹砍伐人工成本大于竹材销售收入, 造成了浙江省较大面积的集约经营毛竹林疏于管理, 处于自然封育状态。此外, 由于经济发展过程中的人口城市化迁移, 大量青壮年人口涌入经济发达地区, 致使欠发达山区的青壮年劳动力水平下降, 加剧了农村人口老龄化问题[16], 在一定程度上也会造成毛竹林的荒芜。自然封育后毛竹林的群落结构和林下植物的物种组成、多样性如何演变, 至今还未见报道。鉴于此, 本研究以浙江省典型毛竹纯林为研究对象, 采用时间代替空间的方法, 通过建立标准地全面调查和研究了不同封育年限毛竹林的胸径分布和林下植被多样性, 为毛竹林的群落演替和生态功能的评价提供基础和理论依据。
1 材料与方法 1.1 研究区概况研究区位于有“中国竹子之乡”之称的浙江省杭州市余杭区境内, 地理坐标为E119°42′2.71″—119°50′35.22″、N30°25′28.1″—30°33′34″。属于北亚热带南缘季风气候区, 冬夏长春秋短, 温暖湿润, 四季分明, 光照充足, 雨量充沛, 年平均气温15.3—16.2℃, 年平均降雨量1391.8mm, 年平均相对湿度76%, 无霜期199—328d。
1.2 试验设计与研究方法2017年7月, 通过查阅杭州市余杭区森林资源经营档案和全面踏查的基础上, 选择余杭西部的百丈镇和鸬鸟镇封育年限分别为10年、20年和30年的毛竹纯林作为研究对象, 以常规经营的竹林为对照(0年), 设置不同封育年限样地各9个, 共计36个样地, 样地面积为30m×30m。封育毛竹林常年没有人为活动干扰, 没有挖笋、清理老竹等活动, 完全处于自然封育状态。常规经营毛竹林每年5月上、中旬施肥1次, 施用600 kg/hm2的复合肥(m(N):m(P2O5):m(K2O)为15:15:15), 肥料地表撒施, 结合松土垦复15 cm左右。常年进行挖笋活动(春笋、鞭笋、冬笋), 毛竹砍伐集中在小年。样地基本情况如表 1所示。
| 封育年限 Number of years of exclosure/a |
土壤类型 Soil type |
海拔 Altitude/m |
坡度 Slope/(°) |
毛竹密度 Density/ (株/hm2) |
平均胸径 Diameter at breast height/cm |
平均株高 Average plant height/m |
| 0 | 红壤 | 275±71 | 15.3±7.5 | 2345±257 | 10.2±0.66 | 13.2±1.0 |
| 10 | 红壤 | 397±75 | 20.2±8.7 | 3773±554 | 10.1±0.79 | 13.5±1.2 |
| 20 | 红壤 | 497±80 | 22.7±7.7 | 5173±459 | 9.6±0.61 | 13.5±1.4 |
| 30 | 红壤 | 509±87 | 19.9±9.8 | 5210±574 | 7.8±0.71 | 13.5±1.5 |
全面测量样地中不同林龄(1度竹、2度以上活立竹)毛竹和枯立竹的胸径, 调查林下灌木、草本植物的种类、株数、高度、冠幅及盖度[17-18]。
1.3 数据处理根据样地调查结果, 计算各封育年限不同林龄毛竹林密度、胸径;同时计算灌、草重要值、Shannon-Wiener指数、Simpson指数、Pielou指数[19]。
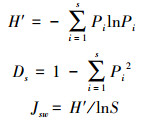
|
H′为Shannon-Wiener指数, Ds为Simpson指数, Jsw为Pielou指数, 式中Pi = ni/N, Pi为第i种植物在样地中出现的概率, N为样地中植物总个体数, S为总种数, ni为第i种植物个体数。
本文数据整理使用Excel 97-03, 显著性差异分析使用SPSS 18.0, 制图使用Origin 8.5。
2 结果与分析 2.1 毛竹林群落结构毛竹林中不同林龄毛竹和枯立竹的密度, 一定程度上反映了毛竹林的结构。由图 1可知, 不同封育年限下毛竹林1度竹的平均密度为727—824株/hm2, 它们之间没有显著差异(P>0.05);封育条件(10年、20年、30年)下毛竹林中2度以上活立竹的平均密度显著高于常规经营(0年), 封育为20、30年的竹林密度又显著高于10年(P < 0.05)。枯立竹的密度随着封育年限的延长而显著增加(P < 0.05)。

|
| 图 1 不同封育年限下毛竹密度和平均胸径 Fig. 1 Density and diameter at breast height of bamboo in different natural exclosure years |
胸径是竹林群落结构的重要指标, 是毛竹的生长状况及土壤养分供给的表现。由表 1和图 1可知, 随着自然封育年限的延长, 毛竹胸径呈现下降的趋势。封育30年的1度竹的平均胸径显著低于其他年限(P < 0.05)。2度以上活立竹则表现为0年、10年显著大于20年(P < 0.05), 同时也显著高于30年(P < 0.05)。枯立竹则表现为10年>20年>30年, 不同封育年限的差异达显著水平(P < 0.05)。
不同林龄毛竹和枯立竹在各封育年限下胸径的分布如图 2所示, 从图 2中可知, 封育年限为0—20年的1度竹胸径分布服从负偏态分布, 峰值向右偏移, 封育30年的1度竹胸径分布则大体服从正态分布, 峰值居中。2度以上活立竹的胸径分布则表现为封育年限为0年和10年呈现负偏态分布, 封育20年表现为正态分布, 封育30年服从正偏态分布(图 2)。枯立竹的胸径分布则表现为封育10年和20年的竹林呈现正态分布, 封育30年表现为正偏态分布。随着封育年限的增加, 胸径分布的峰值会从右往左移动, 即小胸径枯立竹所占比例增加(图 2)。

|
| 图 2 不同封育年龄下毛竹胸径分布情况 Fig. 2 Distribution of diameter at breast height of bamboo in different natural exclosure years |
植物的重要值可以用综合数值来表现该植物物种在群落中的相对重要性, 是评估物种多样性的重要指标。不同封育年限毛竹林下主要灌木植物(重要值≥1)物种组成及重要值如表 2所示, 所有毛竹林中均出现的灌木植物有山莓、络石、紫金牛、山橿、青灰叶下珠和小果菝葜。封育年限为0年时, 重要值排名前5的灌木植物依次为山莓、茶、寒莓、络石、紫金牛, 其中山莓(12.777%)和茶(12.029%)是占优势地位的灌木物种。封育年限为10年的竹林下则依次为络石、山橿、茶、毛花连蕊茶、紫金牛, 其中络石(22.829%)和山橿(19.775%)的重要值明显高于其他灌木植物。20年依次为紫金牛、牛鼻栓、山橿、马银花、异叶爬山虎, 其中紫金牛(10.880%)、牛鼻栓(10.457%)和山橿(9.851%)是毛竹林下主要的优势灌木。30年依次为山橿、牛鼻栓、茶、蓬莱葛、寒莓, 其中山橿(11.702%)的优势度明显。
| 植物类型 Plant species |
植物生活型 Plant life form |
植物重要值The important value of plants | |||
| 0年 | 10年 | 20年 | 30年 | ||
| 山莓Rubus corchorifolius | 高位芽 | 12.777 | 2.809 | 1.807 | 3.134 |
| 寒莓Rubus buergeri | 地上芽 | 8.710 | — | 1.316 | 3.815 |
| 络石Trachelospermum jasminoides | 地上芽 | 6.990 | 22.829 | 3.057 | 2.534 |
| 紫金牛Ardisia japonica | 地上芽 | 5.384 | 3.864 | 10.880 | 3.524 |
| 蓬蘽Rubus hirsutus | 地上芽 | 3.718 | — | — | — |
| 山橿Lindera reflexa | 高位芽 | 3.284 | 19.775 | 9.851 | 11.702 |
| 茶Camellia sinensis | 高位芽 | 12.0289 | 10.4810 | 0 | 5.7528 |
| 青灰叶下珠Phyllanthus glaucus | 高位芽 | 2.523 | 2.595 | 1.568 | 1.221 |
| 算盘子Glochidion puberum | 高位芽 | 2.462 | — | — | — |
| 山鸡椒Litsea cubeba | 高位芽 | 2.301 | — | — | — |
| 刺楸Kalopanax septemlobus | 高位芽 | 2.223 | — | — | — |
| 紫藤Wisteria sinensis | 高位芽 | 2.213 | — | 2.194 | — |
| 小果菝葜Smilax davidiana | 地上芽 | 2.202 | 1.088 | 2.876 | 2.960 |
| 野鸦椿Euscaphis japonica | 高位芽 | 2.136 | — | — | — |
| 藤葡蟠Broussonetia kaempferi var. australis | 高位芽 | 1.377 | — | — | — |
| 檵木Loropetalum chinense | 高位芽 | 1.366 | — | — | — |
| 白马骨Serissa serissoides | 地上芽 | 1.259 | — | — | — |
| 太平莓Rubus pacificus | 地上芽 | 1.259 | — | 1.783 | — |
| 黄檀Dalbergia hupeana | 高位芽 | 1.097 | — | 1.552 | 1.294 |
| 毛花连蕊茶Camellia fraterna | 高位芽 | — | 6.849 | 2.562 | 1.251 |
| 茶荚蒾Viburnum setigerum | 高位芽 | — | 3.114 | — | 1.380 |
| 山胡椒Lindera glauca | 高位芽 | — | 2.619 | — | 1.735 |
| 豹皮樟Litsea coreana var. sinensis | 高位芽 | — | 2.284 | — | 1.451 |
| 异叶爬山虎Parthenocissus dalzielii | 地上芽 | — | 2.248 | 3.845 | — |
| 薜荔Ficus pumila | 地上芽 | — | 1.717 | — | 3.795 |
| 木蜡树Toxicodendron sylvestre | 高位芽 | — | 1.587 | — | — |
| 南天竹Nandina domestica | 高位芽 | — | 1.443 | — | — |
| 红楠Machilus thunbergii | 高位芽 | — | 1.307 | — | — |
| 中华常春藤Hedera nepalensis var. sinensis | 地上芽 | — | 1.136 | 1.043 | 1.815 |
| 豆腐柴Premna microphylla | 高位芽 | — | 1.115 | — | — |
| 鹰爪枫Holboellia coriacea | 高位芽 | — | 1.049 | — | — |
| 牛鼻栓Fortunearia sinensis | 高位芽 | — | — | 10.457 | 6.891 |
| 马银花Rhododendron ovatum | 高位芽 | — | — | 5.305 | — |
| 格药柃Eurya muricata | 高位芽 | — | — | 3.412 | 2.586 |
| 杉木Cunninghamia lanceolata | 高位芽 | — | — | 2.523 | 2.546 |
| 红枝柴Meliosma oldhamii | 高位芽 | — | — | 2.394 | 1.192 |
| 迎春樱Cerasus discoidea | 高位芽 | — | — | 2.343 | — |
| 朱砂根Ardisia crenata | 地上芽 | — | — | 1.452 | — |
| 大芽南蛇藤Celastrus gemmatus | 地上芽 | — | — | 1.395 | — |
| 大青Clerodendrum cyrtophyllum | 高位芽 | — | — | 1.378 | — |
| 蓬莱葛Gardneria multiflora | 地上芽 | — | — | 1.274 | 5.154 |
| 大果山胡椒Lindera praecox | 高位芽 | — | — | 1.137 | — |
| 钝药野木瓜Stauntonia leucantha | 高位芽 | — | — | 1.087 | — |
| 四照花Dendrobenthamia japonica var.chinensis | 高位芽 | — | — | 1.085 | — |
| 五加Acanthopanax gracilistylus | 高位芽 | — | — | — | 2.686 |
| 凹叶厚朴Magnolia officinalis ssp. biloba | 高位芽 | — | — | — | 2.361 |
| 枫香Liquidambar formosana | 高位芽 | — | — | — | 1.489 |
| 华东椴Tilia japonica | 高位芽 | — | — | — | 1.242 |
| 木犀Osmanthus fragrans | 高位芽 | — | — | — | 1.127 |
| 南五味子Kadsura longipedunculata | 地上芽 | — | — | — | 1.023 |
| 细叶青冈Cyclobalanopsis myrsinifolia | 高位芽 | — | — | — | 1.004 |
不同封育年限毛竹林下主要草本植物(重要值≥1)物种及重要值如表 3所示, 所有封育年限下均有分布的草本植物有褐果薹草、求米草、华东蹄盖蕨、玉竹、阔叶箬竹和金星蕨。封育年限为0年时, 重要值前5的草本依次为金毛耳草、疏羽凸轴蕨、淡竹叶、褐果薹草、求米草;10年时依次为麦冬、褐果薹草、华东蹄盖蕨、条穗薹草、紫萁;20年时则为求米草、阔叶箬竹、华东蹄盖蕨、条穗薹草、麦冬;30年依次为阔叶箬竹、绵穗苏、玉竹、山麦冬、华东蹄盖蕨。常规经营条件下(0年), 金毛耳草(11.999%)和疏羽凸轴蕨(9.252%)的重要值明显大于其他草本。封育为10年时, 麦冬(15.029%)、褐果薹草(10.931%)、华东蹄盖蕨(10.785%)和条穗薹草(10.649%)在竹林下都占据一定重要值。随着封育的进行, 20年时求米草(20.159%)、阔叶箬竹(16.065%)和华东蹄盖蕨(11.234%)的重要值增大到一定水平。封育为30年时, 阔叶箬竹(37.224%)成为林下绝对优势草本。
| 植物类型 Plant species |
植物生活型 Plant life form |
植物重要值The important value of plants | |||
| 0年 | 10年 | 20年 | 30年 | ||
| 金毛耳草Hedyotis chrysotricha | 地面芽 | 11.999 | 3.238 | — | — |
| 疏羽凸轴蕨Metathelypteris laxa | 地面芽 | 9.252 | 7.534 | — | 3.941 |
| 淡竹叶Lophatherum gracile | 地面芽 | 6.692 | — | — | — |
| 褐果薹草Carex brunnea | 地面芽 | 6.460 | 10.931 | 1.256 | 4.179 |
| 求米草Oplismenus undulatifolius | 一年生 | 5.810 | 6.900 | 20.159 | 1.247 |
| 青绿薹草Carex breviculmis | 地面芽 | 3.841 | — | — | 2.489 |
| 华东蹄盖蕨Athyrium niponicum | 地上芽 | 3.659 | 10.785 | 11.234 | 4.280 |
| 山麦冬Liriope spicata | 地面芽 | 2.962 | — | 1.691 | 4.659 |
| 南山堇菜Viola chaerophylloides | 地面芽 | 2.763 | — | — | — |
| 芒萁Dicranopteris pedata | 地上芽 | 2.729 | — | — | — |
| 水蓼Polygonum hydropiper | 一年生 | 2.704 | — | — | — |
| 玉竹Polygonatum odoratum | 隐芽 | 2.594 | 1.597 | 5.375 | 4.896 |
| 三叶委陵菜Potentilla freyniana | 地面芽 | 1.925 | — | — | — |
| 薄叶假耳草Neanotis hirsuta | 地面芽 | 1.897 | — | — | — |
| 阔叶箬竹Indocalamus latifolius | 地上芽 | 1.889 | 4.721 | 16.065 | 37.224 |
| 粗壮小鸢尾Iris proantha var. valida | 地面芽 | 1.778 | — | 3.000 | 2.331 |
| 柔枝莠竹Microstegium vimineum | 一年生 | 1.640 | 1.690 | — | — |
| 鱼腥草Houttuynia cordata | 隐芽 | 1.562 | — | — | — |
| 堇菜Viola arcuata | 地面芽 | 1.548 | — | — | — |
| 金星蕨Parathelypteris glanduligera | 地面芽 | 1.390 | 3.046 | 1.728 | 2.912 |
| 三穗薹草Carex heterostachya | 地面芽 | 1.250 | — | — | — |
| 海金沙Lygodium japonicum | 地上芽 | 1.249 | — | — | 1.511 |
| 泽珍珠菜Lysimachia candida | 地面芽 | 1.183 | — | — | — |
| 山萆薢Dioscorea tokoro | 隐芽 | 1.132 | — | — | 1.026 |
| 疏花野青茅Deyeuxia effusiflora | 地面芽 | 1.116 | — | 1.653 | 1.724 |
| 油点草Tricyrtis chinensis | 地面芽 | 1.106 | — | — | — |
| 微糙三脉紫菀Aster ageratoides var. scaberulus | 地面芽 | 1.037 | — | — | — |
| 鸡矢藤Paederia scandens | 地上芽 | 1.011 | — | — | — |
| 麦冬Ophiopogon japonicus | 地面芽 | — | 15.029 | 5.510 | — |
| 线穗薹草Carex nemostachys | 地面芽 | — | 10.649 | 6.516 | — |
| 紫萁Osmunda japonica | 地上芽 | — | 8.247 | — | 2.516 |
| 水竹Phyllostachys heteroclada | 高位芽 | — | 2.607 | 3.280 | — |
| 狗脊蕨Woodwardia japonica | 地上芽 | — | 1.972 | — | — |
| 阔鳞鳞毛蕨Dryopteris championii | 地上芽 | — | 1.972 | — | — |
| 牛尾菜Smilax riparia | 地上芽 | — | 1.788 | — | — |
| 兔儿伞Syneilesis aconitifolia | 地面芽 | — | 1.717 | — | — |
| 红盖鳞毛蕨Dryopteris erythrosora | 地上芽 | — | — | 4.679 | 2.361 |
| 绵穗苏Comanthosphace ningpoensis | 地面芽 | — | — | 4.067 | 5.258 |
| 苦竹Pleioblastus amarus | 地上芽 | — | — | 2.657 | — |
| 心叶堇菜Viola concordifolia | 地面芽 | — | — | 1.724 | — |
| 虎杖Reynoutria japonica | 地面芽 | — | — | — | 2.783 |
| 剪夏罗Lychnis coronata | 隐芽 | — | — | — | 2.301 |
| 铁灯兔儿风Ainsliaea kawakamii | 地上芽 | — | — | — | 1.289 |
不同封育年限毛竹林下灌木的优势度指数Simpson、多样性指数Shannon-Wiener和均匀度指数Pielou如表 4所示。从表中可知, 封育10年时的毛竹林下灌木的Simpson指数显著低于其他林分(P < 0.05);灌木Shannon-Wiener指数则表现为20年和30年的林分显著高于10年和0年(P < 0.05), Pielou指数则表现为0年显著低于10年、20年和30年的林分(P < 0.05), 10年、20年和30年间数值存在波动, 峰值出现在20年。
| 封育年限 Number of years of exclosure/a |
优势度指数 Simpson dominance index |
多样性指数 Shannon-Wiener diversity index |
均匀度指数 Pielou evenness index |
| 0 | 0.947±0.015a | 2.191±0.155b | 0.519±0.037c |
| 10 | 0.886±0.028b | 2.225±0.143b | 0.626±0.039ab |
| 20 | 0.952±0.012a | 2.671±0.097a | 0.658±0.024a |
| 30 | 0.964±0.009a | 2.541±0.112a | 0.585±0.026b |
从表 5可知, 随着封育年限增长, 草本植物的Simpson指数呈现下降趋势, 0年的优势度指数显著高于10年和20年(P < 0.05), 同时显著高于30年(P < 0.05)。多样性指数Shannon-Wiener表现为常规经营(0年)的林分显著高于封育20年以后的林分(P < 0.05), 而不同封育年限间的差异不显著。自然封育10年草本的均匀度指数Pielou显著高于其他林分(P < 0.05), 且封育为0年和20年林分中的草本均匀度指数Pielou又显著高于自然封育为30年的林分(P < 0.05)。
| 封育年限 Number of years of exclosure/a |
优势度指数 Simpson dominance index |
多样性指数 Shannon-Wiener diversity index |
均匀度指数 Pielou evenness index |
| 0 | 0.955±0.011a | 2.699±0.106a | 0.649±0.026b |
| 10 | 0.918±0.017b | 2.478±0.114ab | 0.791±0.036a |
| 20 | 0.903±0.022b | 2.277±0.136b | 0.663±0.039b |
| 30 | 0.844±0.027c | 2.149±0.158b | 0.568±0.042c |
毛竹林总密度随着封育年限的延长而增加。在自然封育条件下, 单位面积毛竹林中1度竹株数没有发生明显变化, 2度以上活立竹株数逐渐增加, 封育20年后基本保持稳定。
1度竹株数在不同封育年限间没有显著差异。主要原因是常规经营毛竹林样地的1度竹受人为选择性留养, 一般种竹留养量约800株/hm2。自然封育10年后, 毛竹林密度达3000株/hm2, 由于水、肥、光等因子的限制, 新竹成竹的难度加大, 1度竹的密度也基本保持在800株/hm2, 随着自然封育时间的进一步延长, 老竹逐渐死亡, 枯立竹数量大大增加, 样地内新生毛竹的数量与死亡毛竹数量基本趋于平衡。
2度以上活立竹逐渐增加的主要原因有:常规经营毛竹林经营中一般遵循“砍四留三”的原则, 即砍去4度以上的毛竹, 而保留3度及以下的毛竹, 除1度竹外, 样地中的毛竹为2—3度, 密度大约在1600株/hm2。自然封育初期, 毛竹生长不再受人为干扰的影响, 毛竹林中包括1—6度活立竹, 因此其密度明显提高。随着自然封育年限的进一步延长, 毛竹林内生存资源开始短缺, 造成了2度以上的活立竹增长速率减缓并最终趋于稳定, 每年枯死的毛竹也在逐渐增加。
毛竹林平均胸径随封育时间的延长而逐步下降。较小胸径毛竹的株数占毛竹总株的比例逐渐增大, 毛竹胸径趋于减小。这与王平等人的研究结果相似, 即当立竹密度增加时, 毛竹平均胸径趋于减小, 毛竹的平均胸径服从自疏法则[20]。在空间和资源的限制下, 竞争是导致毛竹平均胸径下降和总密度趋于稳定的主要因素。自然封育20年后毛竹林总密度没有显著差异, 但平均胸径则表现为自然封育30年的林分显著低于20年。
3.2 自然封育对毛竹林下植物种类与生活型的影响毛竹林下植物群落的科属组成复杂, 植物种类丰富。其中主要灌草植物(表 2—3)隶属50科81属, 总共94种, 其中被子植物42科72属84种, 裸子植物1科1属1种, 蕨类植物7科8属9种。封育年限为0, 10, 20, 30年毛竹林下灌木种类分别为68, 35, 58, 77种, 而草本植物种类则分别为64, 23, 31, 44种。灌草物种丰富度表现为常规经营>自然封育(年限小于等于20年), 这与方茹意等人的研究结果相似, 即适度的人为干扰有利于维持林中灌草的物种丰富度[21]。封育条件下灌、草物种丰富度表现为30年>20年>10年, 毛竹林下灌木、草本植物种类随封育时间的延长逐渐增加。可见长期的自然封育也对提高竹林灌、草物种丰富度有一定的帮助。
在封育过程中, 山橿是一种优势灌木, 在各封育年限(10年、20年、30年)下都有较高重要值, 而在常规经营(0年)竹林下, 山橿的重要值却很低。研究表明山橿属于多年生灌木, 生长缓慢, 一般生长在山地树林下, 喜欢土层深厚, 土壤肥沃和半阴凉的环境[22]。毛竹林下草本层中阔叶箬竹随着封育年限的延长, 重要值连续升高, 封育30年后, 其值高达37.224%, 是该阶段的绝对优势草本。阔叶箬竹为灌木状草本, 径0.5—1.5cm, 节间长5—22cm, 高1—1.5m, 最大竿高可达2m, 对土壤要求不严, 较耐寒, 喜湿耐旱, 可在山谷、山麓和山腰地带等迅速生长扩延[23-24], 对土壤、温度、水分的要求不高, 且作为灌木状草本, 其株高、盖度在吸收光线方面占有一定优势, 竞争能力极强。因此阔叶箬竹逐渐淘汰其他草本成为长期封育毛竹林下的绝对优势草本。
如表 2、3所示, 林下主要灌、草植物按照出芽方式分为5种生活型(高位芽、地上芽、地面芽、隐芽和一年生植物), 不同封育年限下各生活型重要值之和的变化一定程度上能表现林下灌、草植被在自然封育条件下的演替方向(图 3)。

|
| 图 3 毛竹林下不同生活型主要灌草的重要值变化情况 Fig. 3 Changes in important values of main shrubs and herbs under different life forms under bamboo forests |
根据图 3可知, 地面芽草本植物在毛竹林中随封育年限的延长有较为明显的衰退, 而地上芽草本植物的重要值则在该过程中显著增长, 此外一年生草本植物在封育为30年时几乎绝迹。灌木植物的重要值变化规律相对不明显, 表现为高位芽灌木重要值小幅上升, 地上芽灌木重要值小幅下降。
植株的出芽高度一定程度上体现了其对光照的竞争能力, 自然封育条件下, 在林分日渐闭郁的竹林中, 出芽较高的植物相对容易接受光照, 这点在生态位处于最下层的草本植物中有更为明显的体现。因此, 长期的自然封育过程中, 由于竹林密度增大, 光照逐渐稀缺, 拥有易受光和耐阴特性的林下植被更容易在竹林下繁荣。
3.3 自然封育对毛竹林下植物多样性指数的影响优势度指数Simpson、多样性指数Shannon-Wiener和均匀度指数Pielou等多样性指数可以表征林分的植物多样性情况, 体现不同封育阶段林分下灌木层和草本层的植物资源丰富程度。封育为10年的毛竹林分下灌木层的优势度指数Simpson显著低于其他林分。主要是该阶段的林分下存在绝对优势的灌木植物, 从表 1可知, 络石和山橿在封育为10年的林下占据极高的重要值, 其值分别为22.829%, 19.775%。灌木多样性指数Shannon-Wiener在封育过程中大体表现为先升高后降低, 其中以封育20年的林分为最大。随着封育时间的进一步延长, 灌木多样性呈现下降趋势, 主要原因是由于毛竹密度增加到一定阈值, 林分闭郁, 林下光线匮乏, 植物间的竞争更为激烈, 抑制了灌木植物的生长[25-26]。灌木均匀度指数Pielou随封育年限的延长先增大而后降低, 0年和封育30年的林下灌木均匀度指数Pielou较低。这可能是由于0年和30年的林下灌木物种丰富度S较高, 致使均匀度指数的计算公式Jsw = H′/InS中的分母InS比较大, 导致Pielou指数值的下降。这与马克平等人的研究结果相似, 即物种的均匀度指数Pielou与多样性指数Shannon-Wiener呈正相关, 与物种丰富度S呈负相关[27]。综上, 在一定时长范围内, 自然封育可以提高毛竹林下灌木植物的多样性, 但长期的自然封育, 灌木群落多样性会呈现为一定程度的衰退。
草本植物的优势度指数Simpson随封育年限的增加而下降, 草本植物阔叶箬竹的优势地位在封育过程中逐渐突出, 封育年限为30年时在林下占有绝对优势。多样性指数Shannon-Wiener也随着封育时间的延长而逐渐下降, 表明草本植物的多样性水平与封育年限呈负相关。均匀度指数Pielou与封育年限大体呈负相关的关系。随着封育的进行, 草本植物群落逐渐衰退, 个别草本的优势地位逐渐突出, 草本植物的物种多样性水平明显下降。这主要是由于在日渐闭郁的林分中, 竹林密度日益增大, 生态位处于最下层的草本植物, 对光照等资源的反应更为敏感, 面临的竞争更为残酷, 草本层植物群落衰退, 只有少数适应能力强的种类可以在林下大量生长, 成为绝对优势的品种[28]。
| [1] |
崔宁洁, 陈小红, 刘洋, 张健, 杨万勤. 不同林龄马尾松人工林林下灌木和草本多样性. 生态学报, 2014, 34(15): 4313-4323. |
| [2] |
闫明, 毕润成. 山西霍山植被分类及不同演替阶段群落物种多样性的比较分析. 植物资源与环境学报, 2009, 18(3): 56-62. DOI:10.3969/j.issn.1674-7895.2009.03.009 |
| [3] |
冯耀宗. 物种多样性与人工生态系统稳定性探讨. 应用生态学报, 2003, 14(6): 853-857. DOI:10.3321/j.issn:1001-9332.2003.06.004 |
| [4] |
吴彦, 刘庆, 何海, 林波. 亚高山针叶林人工恢复过程中物种多样性变化. 应用生态学报, 2004, 15(8): 1301-1306. DOI:10.3321/j.issn:1001-9332.2004.08.001 |
| [5] |
吴伟光, 曹先磊. 毛竹林经营投入产出关系与经营效益分析. 南京林业大学学报:自然科学版, 2016, 40(3): 108-114. |
| [6] |
姚钧能, 吕建华, 俞卫良, 张君波, 雷赵枫, 李全, 宋新章. 氮沉降和经营强度对毛竹林凋落叶生态化学计量特征的影响. 应用生态学报, 2018, 29(2): 467-473. |
| [7] |
浙江省林业厅.浙江省森林资源及其生态功能价值公告.浙江日报, 2018-01-25(12).
|
| [8] |
胡可易, 柳丽娜, 李雪涛, 金爱武. 经营干扰对毛竹林土壤肥力及胸径分布的影响. 浙江林业科技, 2013, 33(5): 52-56. DOI:10.3969/j.issn.1001-3776.2013.05.011 |
| [9] |
张刚华, 萧江华, 聂洁珠, 陈双林, 郭子武. 不同类型毛竹林植物物种多样性研究. 林业科学研究, 2007, 20(5): 615-621. DOI:10.3321/j.issn:1001-1498.2007.05.005 |
| [10] |
张飞萍.毛竹林节肢动物多样性及主要叶部害虫发生机理[D].福州: 福建农林大学, 2003.
|
| [11] |
何艺玲.不同类型毛竹林林下植被的发育状况及其与土壤养分关系的研究[D].北京: 中国林业科学研究院, 2000.
|
| [12] |
郭文霞, 牛树奎, 张彦龙, 李艳霞, 笪志祥. 经营强度对毛竹林植物多样性的影响. 河北农业大学学报, 2009, 32(4): 46-52. DOI:10.3969/j.issn.1000-1573.2009.04.010 |
| [13] |
郑成洋, 何建源, 罗春茂, 方燕鸿. 不同经营强度条件下毛竹林植物物种多样性的变化. 生态学杂志, 2003, 22(6): 1-6. DOI:10.3321/j.issn:1000-4890.2003.06.001 |
| [14] |
黄继育, 黄勇, 张健. 浙江省安吉县毛竹笋竹两用林土壤养分分析. 世界竹藤通讯, 2017, 15(2): 10-14. |
| [15] |
李海宾. 我国劳动力价格变化与经济增长的实证分析. 中国劳动, 2015(14): 4-9. |
| [16] |
王桂新. 高度重视农村人口过快老龄化问题. 探索与争鸣, 2015(12): 28-30. DOI:10.3969/j.issn.1004-2229.2015.12.008 |
| [17] |
曹志华, 张文斌, 吴中能, 刘俊龙, 苗婷婷, 孙慧. 垦复与砍灌措施对毛竹林生长和竹林结构的影响. 世界竹藤通讯, 2018, 16(2): 7-10. |
| [18] |
侯琳, 雷瑞德, 康博文, 王得祥. 黄龙山林区油松林封育过程中植物物种多样性特征. 西北植物学报, 2004, 24(7): 1165-1172. DOI:10.3321/j.issn:1000-4025.2004.07.001 |
| [19] |
刘晓琴, 张翔, 张立锋, 李英年, 赵亮, 徐世晓, 李红琴, 马荣荣, 牛犇, 高玉葆, 古松. 封育年限对高寒草甸群落组分和物种多样性的影响. 生态学报, 2016, 36(16): 5150-5162. |
| [20] |
王平, 林树燕, 范婷婷, 张玉, 李芳, 凡美玲, 方水元, 王福升, 董丽娜, 时培建. 南京紫金山毛竹平均胸径与立株度之间的关系. 安徽农业科学, 2018, 46(7): 106-108. DOI:10.3969/j.issn.0517-6611.2018.07.033 |
| [21] |
方茹意, 吴炜, 鲁翔, 周柳江, 钟意, 桂仁意. 不同强度人为干扰对毛竹林植物多样性的影响. 世界竹藤通讯, 2017, 15(6): 1-5. |
| [22] |
陈随清, 张峰, 王三姓, 卢玉兵, 潘永峰. 山橿生物特性的初步研究. 中国野生植物资源, 2009, 28(2): 46-49. DOI:10.3969/j.issn.1006-9690.2009.02.013 |
| [23] |
吴丽萍. 云和阔叶箬竹资源调查与发展思考. 吉林农业, 2010(5): 95-96. |
| [24] |
肖朵, 张奇奇, 王艺循. 浅谈阔叶箬竹的引种及栽培技术. 现代种业, 2011(2): 26-27. |
| [25] |
巢林, 洪滔, 李键, 陈灿, 洪伟, 吴承祯. 中亚热带不同林龄杉木人工林径级结构与林下物种多样性分析. 植物资源与环境学报, 2015, 24(2): 88-96. DOI:10.3969/j.issn.1674-7895.2015.02.13 |
| [26] |
黄新荣, 黄承标, 岑国峰, 邓荣艳, 朱木花. 不同林龄马尾松人工林林下植物多样性研究. 农业研究与应用, 2013(4): 1-6. DOI:10.3969/j.issn.2095-0764.2013.04.001 |
| [27] |
马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268-277. DOI:10.3321/j.issn:1000-0933.1995.03.006 |
| [28] |
朱锦懋, 江训强, 黄儒珠, 姜志林. 毛竹林物种多样性的初步分析. 福建林学院学报, 1996, 16(1): 5-8. |