 2020, Vol. 40
2020, Vol. 40文章信息
- 张尹, 于志国, 金彪
- ZHANG Yin, YU Zhiguo, JIN Biao
- 典型温带雨养泥炭沼泽湿地地下部二氧化碳和甲烷浓度变化规律及其影响因素
- The variation of CO2 and CH4 concentrations and their influencing factors in the underground of typical temperate ombrotrophic bogs
- 生态学报. 2020, 40(24): 8936-8947
- Acta Ecologica Sinica. 2020, 40(24): 8936-8947
- http://dx.doi.org/10.5846/stxb202002250337
-
文章历史
- 收稿日期: 2020-02-25
- 网络出版日期: 2020-11-05

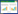




2. 水利部珠江水利委员会水文水资源局, 广州 510611;
3. 中国科学院广州地球化学研究所, 广州 510640
2. Bureau of Hydrology and Water Resources, Pearl River Water Resources Commission of the Ministry of Water Resources, Guangzhou 510611, China;
3. Guangzhou Institute of Geochemistry, Chinese Academy of Sciences, Guangzhou 510640, China
泥炭沼泽湿地(简称“泥炭地”)面积虽只占全球陆地表面积的3%, 其碳储存却占陆地生态系统碳库的三分之一(390-455 Pg)[1-3]。雨养型泥炭地是一种主要依靠大气降水来实现养分和水分补给的泥炭地, 其主要分布在北温带至北极地区[1, 3]。泥炭地中的植物通过光合作用将大气中的碳固定在泥炭土壤中, 由于泥炭土壤中的有机质分解缓慢, 泥炭地便成为重要的碳汇[1]。然而泥炭地并不是稳定的碳汇[4], 前期观测结果以及地球系统模式预测均显示:气候变化的影响下, 其碳汇的能力有减弱趋势[5], 当泥炭地中有机质分解速率加大, 排放到大气中的碳量大于固定的碳量时, 泥炭地便由碳汇转变为碳源。
研究表明, 二氧化碳(CO2)和甲烷(CH4)是泥炭沼泽湿地排放的两种最重要的温室气体, 它们的排放与土壤温度、湿度、pH值、地上生物量、生态系统呼吸等因素有关[6-9]。宋长春等[6]以及郝庆菊等[10]对三江平原泥炭沼泽湿地研究发现, 泥炭地的CO2和CH4排放随温度升高而增加。Laiho[9]研究发现, 原本处于淹水状态的湿地在地下水位下降时可加快有机质分解速率, 促进CO2的排放;反之, 当水位上升, 则可能使CH4排放增加, 而CO2排放减少。Bergman等[11]研究发现, pH值变化会直接影响微生物活性和群落分布, 从而影响其分解有机质的效率。室内模拟实验和模式评估方式普遍认为, 环境因素是直接影响土壤碳库有机碳含量的主要因素[12], 然而Doetterl等[13]研究发现, 在长时间尺度(十年至上百年)内, 环境因素是通过影响土壤风化, 间接影响土壤中的地球化学过程, 从而影响土壤有机质的含量及温室气体的排放。当前关于泥炭地碳排放的研究主要是以地表生态系统排放通量观测为主[14-15], 但是碳通量观测容易受到周围环境的影响, 如风速、地表植被演替等, 直接导致观测结果准确度的不确定性, 而地下部泥炭土壤的生物地球化学过程与碳排放密切相关, 但当前探讨地下部碳排放过程的相关研究较少, 因此本文旨在通过原位不同深度尺度被动式采样方式, 探讨高纬地区的地下部泥炭土壤中CO2和CH4的浓度变化特征及其关键影响因素, 以期从根本上提升碳排放估算的准确性, 为后续碳模型改进提供数据支持, 为未来气候变化趋势研究提供参考依据。
1 材料与方法 1.1 采样点概况本文的两个采样点贝尔山湿地(BBM)和舒特兹山湿地(SBM), 位于德国中部图林根州的图林根森林地区, 均为典型的雨养型泥炭地, 其养分和水分主要来源于大气降水, 无机盐含量低(微摩尔浓度级), 孔隙水导电率极低(10-15 S/m)[4]。BBM位于贝尔山北坡(50°39′34″N, 10°44′34″E), 海拔高度为945-983 m, 年平均降水量为1300 mm, 年平均气温为4 ℃[16]。BBM泥炭土层厚度为3-4 m, 主要植被类型为灌木丛和泥炭藓。SBM位于海拔高度884-893 m的山脊上(50°41′44″N, 10°41′58″E), 年平均降水量为1230 mm, 年平均气温为5 ℃, 泥炭土层厚度约2.5 m, 主要植被类型为灌木丛和泥炭藓[17]。研究所选采样点均位于图林根森林的自然保护区内, 泥炭发育较为完整且受人类活动影响较少, 因而具有较好的代表性及重要的研究价值[18]。
1.2 采样方法利用原位被动式高分辨率采样器(multilevel piezometer, MLP)采集原位泥炭土壤剖面中不同深度的孔隙水和溶解性气体, 如图 1所示, MLP由多段采集管通过螺纹连接而成, 土壤中的孔隙水和气体可以通过采集管管壁的通孔扩散到采集管中, 其中采集管内的上部为硅胶管, 用于采集气体样品, 下部为样品瓶, 用于采集孔隙水。为研究淹水厌氧条件下地下部温室气体排放规律, 采样时间定为降水量较大的夏季, 为2018年6-7月, 以确保泥炭地处于淹水状态。MLP置于泥炭土壤中进行分段采样, 采样的深度分辨率为:150 cm深度以上每隔10 cm采集一个样品, 150 cm深度以下每隔20 cm采集一个样品。平衡5周后, 将MLP分段逐步取出, 每段采集管采集得到的样品储存于容积为20 mL的顶空样品瓶中, 然后注入超纯水并用0.2 μm渗透膜覆盖。静置一段时间后, 在顶空样品瓶中抽取2 mL气体样品, 注入盛有50 μL 6 mol/L HCl的7.5 mL顶空样品瓶中, 用丁基橡胶塞(Wicom, Heppenheim, Germany)紧塞瓶口, 随后测定溶解性温室气体的浓度。因泥炭浅层生物地球化学过程活跃, 故参考Hesslein[19]的方法, 使用深度分辨率为1 cm的被动式透析采样器(dialysis chamber, DC)(60 cm长)对浅层泥炭土进行采样, 采样平衡时间为5周。将每段采样器中收集到的孔隙水样品注入盛有20 μL 6 mol/L HCl的1.8 mL顶空样品瓶中备用, 随后测定溶解性温室气体的浓度。地下水位利用水位传感器(MX2001-S, ONSET, USA)测定。

|
| 图 1 原位被动式高分辨率采样器采集管示意图 Fig. 1 Schematic design of a multilevel piezometer (MLP) segment |
泥炭土壤样品利用俄罗斯箱式采样器(Peat sampler, Eijkelkamp, Netherlands)采样, 受地形变化差异, 不同采样点泥炭累积厚度略有差异, 其中BBM的采样深度为340 cm, SBM的采样深度为210 cm。对采集到的泥炭芯每隔5-10 cm进行分层, 并置于采样袋冷冻保存以待分析。
由于本研究的采样为被动式扩散采样法, 孔隙水及气体扩散收集到的样品为长期动态平衡的结果, 与自然环境中的实际浓度较为接近, 因此样品的代表性较高。整个采样过程持续5周, 取最后一天累积的样品量作为研究值, BBM和SBM采样点均无作重复采样。
1.3 气体样品、孔隙水样品和土壤样品分析方法CO2和CH4的浓度利用气相色谱仪(SRI 8610 C, SRI Instruments, USA)测定, 该仪器由电子捕获检测器(Electron Capture Detector, ECD)和火焰电离检测器(Flame Ionization Detector, FID)组成, 色谱柱为Haysep-D。FID可检测甲烷(CH4)和二氧化碳(CO2), 两种气体的最低检测限均为0.01 mg/L, 检测时每10分钟向仪器内注入1 mL室内空气样品。CO2和CH4的浓度应用亨利定律计算得到, 亨利常数参考Lide等[20]的方法确定。
对于可溶性有机碳(dissolved organic carbon, DOC)的浓度分析, 先用0.45 μm滤膜(NylonMillipore)过滤孔隙水样品, 然后用总有机碳分析仪(TOC 5050, Shimadzu, Japan)进行分析。pH值利用pH计来测定, MLP和DC的每段采样段均需测量pH值。
碳氮比(C/N)可用于衡量泥炭分解程度以及碳、氮的质量损失随深度的变化情况[21]。总C和总N量利用元素分析仪(NA 1108, CE Instruments, Italy)测定, 13C/12C和15N/14N利用同位素比质谱仪(delta S, Finnigan MAT, Germany)测定。同位素比值用式(1)的δ来表示, 用空气中的N2作为N的标准气体。
利用傅里叶变换红外光谱(Fourier Transform Infrared Spectroscopy, FTIR)可研究泥炭土中有机物和矿物的分子结构及其官能团组成[22], 以及由于腐殖化过程引起的有机质组分的变化, 例如腐殖化过程中脂肪族和芳香族等物质的积累[23]。取200 mg干燥KBr和2 mg泥炭土壤样品利用氧化锆球磨后测定波数范围为4500-300 cm-1的红外光谱。本研究定义以下4个吸收峰与多糖(1090 cm-1)的比值为腐殖化系数:1720/1090、1630/1090、1510/1090和1420/1090。其中, 1720 cm-1代表羧酸和芳香酯, 1630 cm-1代表芳烃和芳香族或脂肪族羧酸盐, 1510 cm-1代表酰胺, 1420 cm-1代表酚醛和脂肪族结构[23-24]。
样品测定过程中, 每个采集到的气体样品、孔隙水样品和土壤样品均做了3次重复测定。
1.4 同位素的计算方法利用稳定同位素比质谱仪(GC-C-IRMS, deltaplus, Thermo, Germany)对MLP采样得到的CO2和CH4进行稳定同位素测量, δ13C的值利用参考气体(CO2, -21.225‰)和2个同位素值标准(CO2 50000 mg/L, -39.386‰; CH4 10000 mg/L, -46.362‰)来修正。多次测量得到的平均标准偏差范围为0.09‰(CH4)-0.24‰(CO2)。同位素的计算公式如下:

|
(1) |
式中, δ表示以V-PDB为标准的样品与标准样品偏离的千分率, Rsample表示待测样品中重碳同位素与轻碳同位素的丰度比, 即13C/12C, Rstandard表示标准物质中重碳同位素与轻碳同位素的丰度比。
通常泥炭土壤中产甲烷方式主要有氢产甲烷过程和乙酸产甲烷过程[25]:

|
(2) |
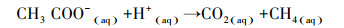
|
(3) |
利用同位素分馏系数αc可衡量泥炭土壤中主要发生的产甲烷方式, 分馏系数αc的计算公式如下:
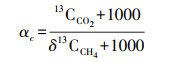
|
(4) |
式中, δ13CCO2和δ13CCH4可由式(1)算得, 当αc < 1.055时, 产甲烷方式以乙酸产甲烷为主, 当αc >1.055时, 产甲烷方式以氢气还原二氧化碳为主[26]。
1.5 数据分析利用Excel 2013和SigmaPlot 14.0软件进行数据处理及分析。采用Pearson相关分析法对碳氮比与FTIR结果及碳、氮同位素比值进行相关性分析。利用SigmaPlot 14.0软件作图, 经计算, 3次重复测定的气体样品、孔隙水样品和土壤样品标准差范围都在1%以内, 因此图表中数据均表示平均值。
2 结果与分析 2.1 地下部温室气体随深度变化特征 2.1.1 地下部CO2和CH4浓度变化特征对于CO2浓度, BBM处除10 cm深度以上浓度较低外(0-2500 μmol/L)(图 2), CO2浓度都随深度在3000 μmol/L左右波动, 10-60 cm处波动幅度较小, 60 cm以下波动幅度较大, 总体呈现随深度减少的趋势。CO2浓度最小值为0.05 μmol/L, 最大值出现在120 cm处, 可达4210.74 μmol/L, 整个垂直采样深度平均值为2568.81 μmol/L。SBM的CO2浓度随深度先增后减, 最小值出现在近地表处, 为573.03 μmol/L, 最大值出现在90 cm, 为4191.94 μmol/L, 平均值为1948.16 μmol/L。0-60 cm处CO2浓度随深度增加缓慢, 在1800 μmol/L左右小幅波动, 60-90 cm随深度急剧增加到最大值, 随后随深度减少, 并且在3000 μmol/L左右波动。

|
| 图 2 BBM和SBM中CO2浓度、CH4浓度和CO2/CH4随深度的变化 Fig. 2 The profiles of concentration of CO2 and CH4 and CO2/CH4 in BBM and SBM |
对于CH4浓度, BBM和SBM处都随深度增加, BBM的甲烷浓度最大值比SBM的大。BBM处CH4浓度为1.88-735.90 μmol/L, 平均值为259.04 μmol/L, 60 cm以上随深度增加幅度较小, 60 cm以下增加幅度较大, 最大值出现在260 cm。SBM的CH4浓度为15.57-543.51 μmol/L, 平均值为301.34 μmol/L, 深度为60 cm以上的CH4浓度都稳定在300 μmol/L左右, 而60 cm以下波动幅度较大, 最大值出现在170 cm。
BBM和SBM的CO2/CH4比率均在近地表处有一峰值。BBM的CO2/CH4比率总体上呈现随深度递减的趋势, 浅层的比率可达16.20, 而150 cm以下的比率仅为4-5。SBM的CO2/CH4比率在60 cm以上变化不大, 60 cm以下处迅速增加到15.14(70 cm)后随深度减少, 并且SBM所有深度中CO2/CH4的比率大多在4-6范围内。
2.1.2 地下部CO2和CH4碳同位素比值变化特征BBM和SBM的δ13CCO2都随深度增加(图 3), 而且值都为-27.40‰--3.13‰。BBM处深度为100 cm以上时, δ13CCO2随深度增加迅速, 100 cm以下随深度增加缓慢。同样地, SBM也出现浅层(80 cm以上)随深度增加迅速, 深层(80 cm以下)随深度增加缓慢的规律。

|
| 图 3 BBM和SBM中δ13CCH4、δ13CCO2和分馏系数αc随深度的变化 Fig. 3 The profiles of δ13CCH4, δ13CCO2 and αc in BBM and SBM |
BBM采样点处δ13CCH4的值(-81.33‰--70.13‰)比SBM的(-70.68‰--60.81‰)小(图 3)。BBM的δ13CCH4在40 cm以上随深度急剧下降, 随后随深度增加, 先减后增是因为在浅层(40 cm以上)12CH4被氧化的量随深度减少, 即12CH4的浓度随深度增加, 根据式(1), δ13CCH4的值在浅层随深度也减少;而SBM的δ13CCH4的值随深度波动下降, 并且波动幅度很大。
与δ13CCO2的变化规律类似, BBM和SBM的分馏系数αc也随深度增加, 并且αc的范围都为1.046-1.077。其中, BBM和SBM分别在20 cm和40 cm深度处αc≈1.055, 由于αc可以判断泥炭沼泽湿地中甲烷产生的主要方式, 因此找到αc为1.055的临界深度即可判断不同深度泥炭土层的产甲烷方式。参考Whiticar[26]定义的分馏系数αc为1.04-1.055时, 泥炭地地下部产甲烷方式以乙酸产甲烷为主, αc为1.055-1.09时, 产甲烷方式以H2还原CO2为主。本研究结果发现, BBM和SBM的αc分别在20 cm和40 cm深度以下为αc>1.055(图 3), 表明BBM和SBM在分别在20 cm和40 cm深度以上是以乙酸产甲烷为主, 20 cm和40 cm以下产甲烷方式都则以H2还原CO2为主, 与前期研究发现的浅层以乙酸产甲烷为主, 深层以H2还原CO2为主的规律一致[27-28]。
2.2 地下部剖面孔隙水中的DOC浓度及pH值随深度变化特征如图 4所示, BBM中的DOC浓度随深度呈逐渐减少趋势, 浓度为14-28 mg/L。SBM中的DOC浓度没有明显的变化规律, 在深度为100 cm以上处波动较大, 100 cm以下波动较小, 其中最大值出现在20 cm处, 为20.81 mg/L, 最小值出现在50 cm处, 为2.22 mg/L。SBM中DOC的浓度(2.22-20.81 mg/L)略少于BBM的DOC浓度(14.75-27.93 mg/L)。

|
| 图 4 BBM和SBM中DOC和pH值分别随深度的变化 Fig. 4 The profiles of concentration of DOC and pH value in BBM and SBM |
BBM和SBM的pH值均随深度升高, SBM随深度变化的幅度比BBM大, BBM的pH值范围为3.49-4.51, SBM的pH值范围为4.00-5.71, 两个采样点pH平均值都为4.3, 符合雨养型泥炭地为酸性的特点[29]。
2.3 泥炭沼泽湿地有机质储量随深度变化特征如图 5所示, BBM和SBM的C/N都是随深度先增后减, 但是BBM中的变化幅度较SBM中的大, BBM和SBM的C/N基本上都大于50, BBM中C/N最大值为206.27(140 cm), SBM的最大值为288.40(100 cm)。

|
| 图 5 BBM和SBM中C/N、δ13C、δ15N随深度的变化 Fig. 5 The profiles of C/N, δ13C and δ15N in BBM and SBM |
BBM和SBM的δ13C都随深度先增大后减少(图 5), 与C/N呈显著正相关(表 1), 并且比值都为-29.17‰--22.63‰。BBM的δ13C最大值为180 cm处的-22.63‰, SBM的δ13C最大值为70 cm处的-24.15‰。
| C/N | |||
| SBM | BBM | ||
| FTIR 1720/1090 | r | -0.518** | -0.194 |
| N | 43 | 23 | |
| FTIR 1630/1090 | r | -0.712** | -0.205 |
| N | 43 | 24 | |
| FTIR 1510/1090 | r | -0.336* | -0.427* |
| N | 43 | 24 | |
| FTIR 1420/1090 | r | -0.533** | -0.379 |
| N | 43 | 24 | |
| δ13C/‰ | r | 0.866** | 0.532** |
| N | 43 | 24 | |
| δ15N/‰ | r | 0.224 | -0.217 |
| N | 43 | 24 | |
| r为皮尔逊相关系数;N为样本数量;*和**分别表示通过0.05和0.01水平的显著性检验; ; FTIR:傅里叶变换红外光谱Fourier transform infrared spectroscopy | |||
BBM和SBM的δ15N随深度变化规律相似, 都是随深度增加。BBM的δ15N(-5.80‰-1.20‰)略小于SBM的δ15N(-2.00‰-2.98‰)。BBM的δ15N随深度波动不大, 除了270-290 cm和330-340 cm处δ15N为正值外, 其余深度均为负值。SBM的δ15N在深度为100 cm以上时在0‰附近波动较大, 100 cm以下波动较小, 且都大于0‰。
BBM中的傅里叶变换红外光谱图中的不同比值(1720/1090、1630/1090、1510/1090、1420/1090)随深度变化都是先减后增(图 6), 变化趋势与BBM中的C/N变化趋势相反(图 6), 且这种负相关关系显著(表 1)。SBM的FTIR图中, 除了1630/1090 50 cm处有一峰值和1720/1090 40 cm处有一最小值外, 不同的比值大致均呈现180 cm以上随深度变化不大, 180 cm以下随深度增加的趋势。

|
| 图 6 BBM和SBM中傅里叶红外变换光谱比值随深度的变化 Fig. 6 The profiles of fourier transform infrared spectroscopy (FTIR) ratio in BBM and SBM |
前期研究发现, 原位泥炭土中的可溶性有机质的FTIR光谱在约1630 cm-1处的吸收峰最强[23-24], 故选取比值为1630/1090作为腐殖化指数。BBM中的腐殖化指数与其C/N呈显著负相关(表 1), 在50 cm以上处腐殖化指数较大, 而对应的C/N较小, 随后腐殖化指数随深度减少至0.4左右(130-150 cm)时, C/N则随深度增加至206.27(140 cm)。在200 cm以下, 腐殖化指数随深度增加至大于0.9, 而C/N随深度减少至50以下。表 1计算结果表明, SBM中的腐殖化指数与C/N也呈负相关关系, 但并不显著。除了50 cm处的腐殖化指数达到1.45外, SBM中的腐殖化指数在190 cm以上处均在0.60-0.85之间, 在190 cm以下处随深度增加至约为1.2。
3 讨论 3.1 原位地下部温室气体浓度影响因素 3.1.1 CO2和CH4气体浓度的影响因素前期研究表明, 地球上高纬度地区(47°-55°)泥炭沼泽湿地中的CO2的浓度范围多为0-6000 μmol/L, 但浓度高的地区, 如英格兰, 可达6000-10000 μmol/L, 而CH4的浓度范围多为0-2000 μmol/L, 并且两种气体的浓度沿土壤剖面变化都是大致随深度增加[6, 28, 30-31]。Beer等[31]和Blodau等[32]分别对加拿大Mer Bleue泥炭地进行原位观测实验发现, CO2的浓度为100-5500 μmol/L, CH4的浓度为0-600 μmol/L, CO2和CH4的浓度均与本研究中两个采样点的结果相近, 因此本研究得到的结果与前期研究结果具有可比性。BBM和SBM的CO2随深度减少, CH4随深度增加, 由于αc可用于判断泥炭地地下部的产甲烷方式(见1.4), 由图 3的αc数据可知, BBM和SBM分别在20 cm和40 cm深度以下存在αc>1.055, 表明在这些深度以下的甲烷产生方式是H2还原CO2为主, CO2被消耗用于产CH4, 因此CO2浓度降低而CH4浓度升高。BBM和SBM的CH4在60cm以下随深度快速增加, 推断是由于随深度增加, 泥炭土壤内部厌氧环境有利于CH4的产生使其积累[27]。BBM和SBM在10cm以上处CO2/CH4的比率都很高, 由图 7可知, 虽然采样时间段内水位一直在地表以上, 但是由于德国的降水量有夏多冬少的特点[33], 因此地下水位会上下波动, 水位波动过程中有氧气的输入, 甲烷被有氧氧化导致其含量极低, 因而导致CO2/CH4的比率极高。10cm以下, BBM和SBM的CO2/CH4比率都随深度增加而减少, 并且比率大多数为4-6, 与前期北半球高纬地区泥炭地研究报道的结果相近[28, 34-35]。

|
| 图 7 BBM和SBM中地下水位随时间的变化 Fig. 7 Variation of water table in BBM and SBM |
由于分解过程产生12CO2、12CH4、13CO2和13CH4, 相对分子质量较轻的12CO2和12CH4会优先排放到大气中, 而相对分子质量较重的13CO2和13CH4则留在土壤中, 利用δ13CCO2和δ13CCH4方法可以对地下部温室气体进行路径示踪研究, 了解气体的去向。本研究中CH4和CO2的δ13C分别为-82‰--60‰和-28‰--3‰, 与前期北半球典型泥炭地的研究结果相当[28, 31, 35]。Hornibrook[27]对加拿大Point Pelee Marsh和Sifton Bog两个湿地研究发现, 两个湿地的δ13CCH4都随深度减少, 比值为-72‰--48‰, 变化规律与本研究中的SBM相似, 但比值均比BBM和SBM的δ13CCH4大, 而δ13CCO2都随深度增加, 比值为-20‰--5‰, 变化规律和比值与BBM和SBM均相似, 主要是因为Point Pelee Marsh和Sifton Bog的12CH4被有氧氧化作用消耗或者以冒泡的方式排放到大气中, 导致12CCH4较小, 进而使得δ13CCH4较大。由此可见, 本研究中2个采样点δ13CCH4的比值较小, 表明大部分的12CH4仍被储存在泥炭土壤中, 该地受外界环境的干扰较少。δ13CCO2随深度增大, 表明12CO2随深度消耗增多, 而消耗的CO2用于H2还原CO2的产甲烷过程, 因为图 3 αc数据表明BBM和SBM分别在20 cm和40 cm以下产甲烷方式都以H2还原CO2为主, 另外, 消耗的CO2也有可能用于产乙酸过程[27]。
3.2 原位地下部温室气体浓度变化与泥炭分解的关系利用C/N可以衡量泥炭土中有机质的分解程度, C/N比值越大, 表明泥炭土的分解程度低, 通常情况下, 泥炭土壤的C/N介乎于40-100, 并且当C/N大于50时表明有机质分解程度较低[1, 22, 24, 36]。本研究中, BBM和SBM的C/N的比值基本上都大于50, 并且BBM在深度为100-270 cm处、SBM在70-100 cm处, C/N大于100, 说明在该深度区域的分解程度十分低, 储存着大量有机碳。SBM和BBM都是约200 cm以下C/N较小, 腐殖化指数较大, 表明深层处有机质分解程度较高。欧洲泥炭土平均沉积速率约为2 mm/yr[37], 如此计算, 200 cm以下的泥炭土层可以代表一千年以前的情况。根据IPCC[38]报告可知, 在公元800-1100年北半球的气温比1100年后的略高, 而温度较高有利于加速有机质分解, 所以两个采样点深层处的腐殖化指数较高。
有机质分解过程伴随着同位素分馏的发生, 分解过程中12C会优先富集在产物, 如CO2、CH4中, 并排放到大气中去, 而13C则富集在分解后残余的有机质中, 因此δ13C偏负值时说明有机质分解程度高, 而偏正值时说明分解程度低。BBM和SBM的δ13C在浅层处较大, 主要是由于德国的降水有季节性差异[33], 地下水位波动过程中有氧气的输入, 可以促进有机质分解, 导致浅层处δ13C更偏负值。此外, 也与采样时间处于夏季, 夏季土壤温度随深度变化比较大, 浅层土壤温度对气温的响应较强, 因而地表温度比较高, 有利于有机质的分解有关[39]。
氮同位素可以研究土壤有机质的来源和分布[40], 本研究中两个采样点的δ15N都是随深度趋于正值, 研究结果与前期研究结果吻合[22, 41]。不同植物的稳定氮同位素比率存在差异, 进而会导致不同地区泥炭土壤中的δ15N的值不同[22, 24], 虽然本研究中SBM的δ15N比BBM中的稍大, 但是两个采样点的覆盖植被类型均为泥炭藓和灌木丛, 因而可以排除植被类型的影响导致两地δ15N存在差异。另外, 温度和降水也可以影响δ15N的变化, 但是影响规律是如何, 至今还没有明确的定论, 需进一步实验验证[42]。
DOC的浓度受多种因素的影响, 例如温度、水位、pH、植被类型、泥炭地演替阶段等[43-44], 本研究发现SBM和BBM中DOC浓度比其他学者的研究结果低[24], 是因为研究地区地处中高纬度地区, 温度较低, 凋落物的分解速度较慢, 另外, 2个采样点覆盖的植被类型均为灌木丛和苔藓, 属于森林演替过程中的早期阶段, 凋落物分解释放DOC的速率较低。另外, 前期研究表明, 产甲烷的最适pH值为5-6[45], 甲烷氧化的最适pH值为5-8[46], 而本研究中2个采样点的泥炭土pH值都较低, 基本都为4.3左右, 因此该土壤环境并不十分利于产甲烷及甲烷氧化的发生, 同时pH值低也会限制微生物活性及有效底物活性, 从而限制有机质分解放出二氧化碳[47]。
总的来说, C/N、碳氮同位素和FTIR方法都表明, 本研究中天然条件下原位泥炭土的有机质整体分解程度较低, 但是在浅层和底层分解程度相对较高。浅层分解程度高是因为降水造成的地下水位波动, 有氧气输入, 促进有机质的分解;而底层泥炭土的深度在200 cm以上, 对应的是沉积了1000多年的泥炭土, 对应历史时期的气温略高, 高温可以促进有机质的分解, 表明该地储存着大量的有机碳, 是重要的碳库, 随着气候变化, 该地有可能成为一个重要的碳源。另外, DOC浓度和pH值是影响有机质分解的关键因子, BBM和SBM两个采样点的低DOC浓度和低pH值均限制了有机质的分解。
4 结论(1) BBM采样点地下部CO2浓度变化总体呈随深度减少趋势, SBM的CO2浓度变化随深度先增后减;BBM和SBM地下部CH4浓度均随深度增加, 并且在60cm以下快速增加, CO2和CH4的浓度变化规律与前期研究结果具有可比性。摸清地下部温室气体浓度变化规律对准确核算地表温室气体排放通量具有重要意义。
(2) 两个采样点δ13CCH4的值均较小, 表明大部分的12CH4仍被储存在泥炭土壤中, 被氧化或排放到大气中的量较少。BBM和SBM的δ13CCO2和分馏系数αc都随深度增加, BBM和SBM分别在20 cm和40 cm以下有αc >1.055, 表明产甲烷方式在该深度以上为乙酸产甲烷为主, 该深度以下为H2还原CO2为主, 深层消耗的12CO2有一部分用于产甲烷过程。
(3) 碳氮比、碳氮同位素比值、傅里叶红外光谱法都表明, BBM和SBM两个采样点的有机质分解程度较低, 是因为低DOC浓度和低pH值不利于有机质的分解, 由此2个采样点储存大量有机碳。
| [1] |
Berger S, Braeckevelt E, Blodau C, Burger M, Goebel M, Klemm O, Knorr K H, Wagner-Riddle C. A 1-year greenhouse gas budget of a peatland exposed to long-term nutrient infiltration and altered hydrology:high carbon uptake and methane emission. Environmental Monitoring and Assessment, 2019, 191(9): 533. DOI:10.1007/s10661-019-7639-1 |
| [2] |
Bonaiuti S, Blodau C, Knorr K H. Transport, anoxia and end-product accumulation control carbon dioxide and methane production and release in peat soils. Biogeochemistry, 2017, 133(2): 219-239. DOI:10.1007/s10533-017-0328-7 |
| [3] |
Xu J R, Morris P J, Liu J G, Holden J. PEATMAP:Refining estimates of global peatland distribution based on a meta-analysis. CATENA, 2018, 160: 134-140. DOI:10.1016/j.catena.2017.09.010 |
| [4] |
Limpens J, Berendse F, Blodau C, Canadell J G, Freeman C, Holden J, Roulet N, Rydin H, Schaepman-Strub G. Peatlands and the carbon cycle:from local processes to global implications-a synthesis. Biogeosciences, 2008, 5(5): 1475-1491. DOI:10.5194/bg-5-1475-2008 |
| [5] |
Todd-Brown K E O, Randerson J T, Post W M, Hoffman F M, Tarnocai C, Schuur E A G, Allison S D. Causes of variation in soil carbon simulations from CMIP5 Earth system models and comparison with observations. Biogeosciences, 2013, 10(3): 1717-1736. DOI:10.5194/bg-10-1717-2013 |
| [6] |
宋长春, 杨文燕, 徐小锋, 娄彦景, 张金波. 沼泽湿地生态系统土壤CO2和CH4排放动态及影响因素. 环境科学, 2004, 25(4): 1-6. |
| [7] |
沙晨燕, 王敏, 王卿, 陆健健. 湿地碳排放及其影响因素. 生态学杂志, 2011, 30(9): 2072-2079. |
| [8] |
李敬, 黄佳芳, 罗敏, 刘育秀, 谭季, 朱爱菊, 王声钰, 仝川. 淹水增加对闽江河口淡水潮汐湿地孔隙水地球化学特征及CO2和CH4排放通量的影响. 环境科学, 2019, 40(12): 5493-5502. |
| [9] |
Laiho R. Decomposition in peatlands:reconciling seemingly contrasting results on the impacts of lowered water levels. Soil Biology and Biochemistry, 2006, 38(8): 2011-2024. DOI:10.1016/j.soilbio.2006.02.017 |
| [10] |
郝庆菊, 王跃思, 宋长春, 王毅勇, 王明星. 三江平原湿地土壤CO2和CH4排放的初步研究. 农业环境科学学报, 2004, 23(5): 846-851. DOI:10.3321/j.issn:1672-2043.2004.05.003 |
| [11] |
Bergman I, Svensson B H, Nilsson M. Regulation of methane production in a Swedish acid mire by pH, temperature and substrate. Soil Biology and Biochemistry, 1998, 30(6): 729-741. |
| [12] |
Carvalhais N, Forkel M, Khomik M, Bellarby J, Jung M, Migliavacca M, Mu M Q, Saatchi S, Santoro M, Thurner M, Weber U, Ahrens B, Beer C, Cescatti A, Randerson J T, Reichstein M. Global covariation of carbon turnover times with climate in terrestrial ecosystems. Nature, 2014, 514(7521): 213-217. DOI:10.1038/nature13731 |
| [13] |
Doetterl S, Stevens A, Six J, Merckx R, Van Oost K, Casanova Pinto M, Casanova-Katny A, Muñoz C, Boudin M, Zagal Venegas E, Boeckx P. Soil carbon storage controlled by interactions between geochemistry and climate. Nature Geoscience, 2015, 8(10): 780-783. DOI:10.1038/ngeo2516 |
| [14] |
李永福, 葛继稳, 翁闻畅, 刘明盟, 斯南雍茜, 程腊梅, 张志麒. 神农架大九湖泥炭湿地二氧化碳和甲烷排放化学计量比研究. 安全与环境工程, 2019, 26(4): 21-28. |
| [15] |
侯颖, 李红, 赵敏, 熊俊, 戴圣骐, 赵斌, 胡静, 郭海强. 从湿地到农田:围垦对生态系统碳排放的影响. 应用生态学报, 2017, 28(8): 2517-2526. |
| [16] |
ThüringenForst. Thüringens Wilde Wälder: Beerbergmoor.[2020-04-25]. https://www.thueringenforst.de/fileadmin/user_upload/Wilde_Waelder/A_C/Beerbergmoor-Handout.pdf.
|
| [17] |
Unterwegs Auf Dem Moorsteg: Schützenbergmoor für Besucher Erlebbar Gemacht.[2020-04-25]. https://www.biosphaerenreservat-thueringerwald.de/de/naturselbsterkunden/unterwegs-auf-dem-moorsteg/.
|
| [18] |
Fischer F. Democracy and Expertise:Reorienting Policy Inquiry. Oxford: Oxford University Press, 2009.
|
| [19] |
Hesslein R H. An in situ sampler for close interval pore water studies. Limnology and Oceanography, 1976, 21(6): 912-914. DOI:10.4319/lo.1976.21.6.0912 |
| [20] |
Lide D R, Frederikse H P R. CRC Handbook of Chemistry and Physics. 78th ed. Boca Raton FL: CRE Press, 1997.
|
| [21] |
Biester H, Hermanns Y M, Martinez Cortizas A. The influence of organic matter decay on the distribution of major and trace elements in ombrotrophic mires-a case study from the Harz Mountains. Geochimica et Cosmochimica Acta, 2012, 84: 126-136. DOI:10.1016/j.gca.2012.01.003 |
| [22] |
Drollinger S, Kuzyakov Y, Glatzel S. Effects of peat decomposition on δ13C and δ15N depth profiles of Alpine bogs. CATENA, 2019, 178: 1-10. DOI:10.1016/j.catena.2019.02.027 |
| [23] |
Beer J, Lee K, Whiticar M, Blodau C. Geochemical controls on anaerobic organic matter decomposition in a northern peatland. Limnology and Oceanography, 2008, 53(4): 1393-1407. DOI:10.4319/lo.2008.53.4.1393 |
| [24] |
Broder T, Blodau C, Biester H, Knorr K H. Peat decomposition records in three pristine ombrotrophic bogs in southern Patagonia. Biogeosciences, 2012, 9(4): 1479-1491. DOI:10.5194/bg-9-1479-2012 |
| [25] |
Ehrlich H L, Newman D K. Lithosphere as microbial habitat//Ehrlich H L, ed. Geomicrobiology. 5th ed. London: CRC Press, 2009: 37-55.
|
| [26] |
Whiticar M J, Faber E, Schoell M. Biogenic methane formation in marine and freshwater environments:CO2 reduction vs. acetate fermentation-isotope evidence. Geochimica et Cosmochimica Acta, 1986, 50(5): 693-709. DOI:10.1016/0016-7037(86)90346-7 |
| [27] |
Hornibrook E R C, Longstaffe F J, Fyfe W S. Spatial distribution of microbial methane production pathways in temperate zone wetland soils:Stable carbon and hydrogen isotope evidence. Geochimica et Cosmochimica Acta, 1997, 61(4): 745-753. DOI:10.1016/S0016-7037(96)00368-7 |
| [28] |
Broder T, Blodau C, Biester H, Knorr K H. Sea spray, trace elements, and decomposition patterns as possible constraints on the evolution of CH4 and CO2 concentrations and isotopic signatures in oceanic ombrotrophic bogs. Biogeochemistry, 2015, 122(2/3): 327-342. DOI:10.1007/s10533-014-0044-5 |
| [29] |
Ye R Z, Jin Q S, Bohannan B, Keller J K, McAllister S A, Bridgham S D. pH controls over anaerobic carbon mineralization, the efficiency of methane production, and methanogenic pathways in peatlands across an ombrotrophic-minerotrophic gradient. Soil Biology and Biochemistry, 2012, 54: 36-47. DOI:10.1016/j.soilbio.2012.05.015 |
| [30] |
Clymo R S, Bryant C L. Diffusion and mass flow of dissolved carbon dioxide, methane, and dissolved organic carbon in a 7-m deep raised peat bog. Geochimica et Cosmochimica Acta, 2008, 72(8): 2048-2066. DOI:10.1016/j.gca.2008.01.032 |
| [31] |
Beer J, Blodau C. Transport and thermodynamics constrain belowground carbon turnover in a northern peatland. Geochimica et Cosmochimica Acta, 2007, 71(12): 2989-3002. DOI:10.1016/j.gca.2007.03.010 |
| [32] |
Blodau C, Roulet N T, Heitmann T, Stewart H, Beer J, Lafleur P, Moore T R. Belowground carbon turnover in a temperate ombrotrophic bog. Global Biogeochemical Cycles, 2007, 21(1): GB1021. |
| [33] |
Bohnenstengel S I, Schlünzen K H, Beyrich F. Representativity of in situ precipitation measurements-A case study for the LITFASS area in North-Eastern Germany. Journal of Hydrology, 2011, 400(3/4): 387-395. |
| [34] |
Steinmann P, Eilrich B, Leuenberger M, Burns S J. Stable carbon isotope composition and concentrations of CO2 and CH4 in the deep catotelm of a peat bog. Geochimica et Cosmochimica Acta, 2008, 72(24): 6015-6026. DOI:10.1016/j.gca.2008.09.024 |
| [35] |
Münchberger W, Knorr K H, Blodau C, Pancotto V A, Kleinebecker T. Zero to moderate methane emissions in a densely rooted, pristine Patagonian bog-biogeochemical controls as revealed from isotopic evidence. Biogeosciences, 2019, 16(2): 541-559. DOI:10.5194/bg-16-541-2019 |
| [36] |
Biester H, Martinez-Cortizas A, Birkenstock S, Kilian R. Effect of peat decomposition and mass loss on historic mercury records in peat bogs from Patagonia. Environmental Science & Technology, 2003, 37(1): 32-39. |
| [37] |
Novak M, Brizova E, Adamova M, Erbanova L, Bottrell S H. Accumulation of organic carbon over the past 150 years in five freshwater peatlands in western and central Europe. Science of the Total Environment, 2008, 390(2/3): 425-436. |
| [38] |
Intergovernmental Panel on Climate Change (IPCC). Climate Change 2007: The Physical Science Basis. Contribution of Working Group to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007.
|
| [39] |
宋长春, 王毅勇. 湿地生态系统土壤温度对气温的响应特征及对CO2排放的影响. 应用生态学报, 2006, 17(4): 625-629. DOI:10.3321/j.issn:1001-9332.2006.04.014 |
| [40] |
汪智军, 梁轩, 贺秋芳, 袁道先. 岩溶区不同植被类型下的土壤氮同位素分异特征. 生态学报, 2011, 31(17): 4970-4976. |
| [41] |
熊鑫, 张慧玲, 吴建平, 褚国伟, 周国逸, 张德强. 鼎湖山森林演替序列植物-土壤碳氮同位素特征. 植物生态学报, 2016, 40(6): 533-542. |
| [42] |
刘贤赵, 张勇, 宿庆, 田艳林, 王庆, 全斌. 陆生植物氮同位素组成与气候环境变化研究进展. 地球科学进展, 2014, 29(2): 216-226. |
| [43] |
Broder T, Knorr K H, Biester H. Changes in dissolved organic matter quality in a peatland and forest headwater stream as a function of seasonality and hydrologic conditions. Hydrology and Earth System Sciences, 2017, 21(4): 2035-2051. |
| [44] |
潘思涵, 程宇琪, 杜浩, 杨宇娜, 王雨晴, 张成福. 大兴安岭森林演替过程中凋落物分解与DOC释放研究. 西南林业大学学报, 2019, 39(5): 75-83. |
| [45] |
Dunfield P, Knowles R, Dumont R, Moore T R. Methane production and consumption in temperate and subarctic peat soils:response to temperature and pH. Soil Biology and Biochemistry, 1993, 25(3): 321-326. |
| [46] |
Semrau J D, DiSpirito A A, Yoon S. Methanotrophs and copper. FEMS Microbiology Reviews, 2010, 34(4): 496-531. |
| [47] |
Scanlon D, Moore T. Carbon dioxide production from peatland soil profiles:the influence of temperature, Oxic/Anoxic conditions and substrate. Soil Science, 2000, 165(2): 153-160. |