 2020, Vol. 40
2020, Vol. 40文章信息
- 张全军, 张广帅, 于秀波, 刘宇, 夏少霞, 孟竹剑, 许策, 胡斌华, 万松贤
- ZHANG Quanjun, ZHANG Guangshuai, YU Xiubo, LIU Yu, XIA Shaoxia, MENG Zhujian, XU Ce, HU Binhua, WAN Songxian
- 鄱阳湖湿地优势植物枯落物的分解速率及碳、氮、磷释放动态特征
- Dynamic characteristics of the decomposition rate and carbon, nitrogen and phosphorus release of the dominant plants in Poyang Lake Wetland
- 生态学报. 2020, 40(24): 8905-8916
- Acta Ecologica Sinica. 2020, 40(24): 8905-8916
- http://dx.doi.org/10.5846/stxb201909161925
-
文章历史
- 收稿日期: 2019-09-16
- 修订日期: 2020-03-10





2. 国家海洋环境监测中心, 大连 116023;
3. 江西鄱阳湖南矶湿地国家级自然保护区管理局, 南昌 330038;
4. 中国科学院大学, 北京 100049
2. National Marine Environmental Monitoring Center, Dalian 116023, China;
3. Nanji Wetland National Nature Reserve Agency, Nanchang 330038, China;
4. University of Chinese Academy of Sciences, Beijing 100049, China
植物枯落物分解是生态系统功能的基本过程, 对生态系统碳通量和养分循环都有至关重要的作用, 能推动养分从植物枯落物返回到环境中, 直接决定着植物吸收养分的有效性, 对植被生产力起着关键的控制作用[1-2]。有研究表明, 枯落物分解速率的变化往往会导致当地、区域甚至全球范围内碳库和氮库的巨大变化[3-4]。湿地生态系统仅覆盖全球面积的4%—9%, 但是它们初级生产力高, 碳储存量却占整个陆地生态系统碳储量的37%左右[5-6]。湿地植物枯落物分解会直接影响湿地生态系统中碳、氮、磷等生物必须营养元素的归还与积累[4, 7-8], 是维持湿地生态系统能量流动、物质循环和养分平衡的关键生态过程[9-10]。
植物枯落物分解是一个非常复杂的过程, 这一过程主要由3个相互作用的因素决定, 即化学(枯落物理化特性)、物理(气候和环境)以及生物(参与枯落物分解的微生物和无脊椎动物)因素[4, 11-12]。在温度和湿度不受限制的生态系统中, 分解速率最重要的决定因素是枯落物的理化性质以及分解发生环境中的养分供应条件和分解者活动情况(即立地环境质量)[13-15]。枯落物的理化特性包括其表面性质、组织结构、营养元素及有机化合物的种类和含量等, Swift等[16]将枯落物的化学属性称之为“基质质量(substrate quality)”, 定义为枯落物的相对可分解性。基质质量的描述指标主要有氮含量、磷含量、木质素含量、纤维素和半纤维素含量、酚类含量、以及C:N、C:P、N:P和木质素:N等比例[11]。基质质量对分解速率有很强的控制作用, 能决定枯落物被破碎分解的难易程度, 是制约枯落物分解的内在因素[17]。在有些情况下从枯落物初始的C:N或C:P就能预测一些湿地枯落物的分解速率[18]。然而, 目前在气候相似但植被类型不同的局部地区枯落物分解的变异性究竟有多大, 还有许多不清楚的地方[19]。研究湿地枯落物分解及其影响机制有助于加深对全球尺度碳、氮生物地球化学循环的认识, 也更加有助于湿地的保护和管理。
鄱阳湖是我国第一大淡水湖泊, 受流域来水和长江水位双重影响, 年内丰水期和枯水期时间界限明显, 丰枯期水位差可达十几米之多[20], 这种独特的水文节律使得鄱阳湖湿地成为典型的季节性洪泛平原湿地, 具有非常大面积干湿交替的洲滩湿地。这些洲滩上主要分布着大量的如芦苇(Phragmites), 南荻(Triarrhena lutarioriparia)、薹草(Carex)等挺水植物和湿生植物[21-24], 这些植物为湿地提供了大量的枯落物, 其分解过程为湿地碳、氮和磷等养分元素的归还与积累做出了巨大的贡献[7, 25]。通过前期研究发, 洲滩地下水位梯度[7]、湿地土壤养分条件和微生物条件[26]以及湿地越冬水鸟粪便[15]等因素对湿地薹草枯落物的分解及养分释放都有非常重要的影响。有研究表明鄱阳湖湿地不同类型的植物枯落物碳、氮和磷化学计量比差异非常明显[27-29], 分解速率可能也有所差异。然而迄今为止, 涉及到鄱阳湖湿地不同类型湿地植物枯落物分解过程及养分元素释放特征的野外长期定位观测实验研究尚少。因此, 本研究通过野外定位观测模拟实验, 研究了鄱阳湖湿地优势植物芦苇、南荻和薹草枯落物分解速率及碳、氮和磷元素释放动态特的征差异性及其形成原因。以期为鄱阳湖湿地碳、氮和磷的生物地球化学循环提供更新的认识, 为鄱阳湖湿地的科学管理、保护与恢复提供科学依据。
1 材料和方法 1.1 研究区域概况本研究区域位于鄱阳湖南矶山湿地国家级自然保护区内典型的碟形湖—白沙湖的洲滩上(图 1)。该保护区位于鄱阳湖主湖区南部, 地处赣江北支、中支和南支汇入鄱阳湖开放水域冲积形成的三角洲前缘, 海拔在12—16 m(吴淞)[30]。属亚热带暖湿型季风气候, 夏季炎热多雨, 冬季低温少雨[31]。受鄱阳湖季节性水文节律的影响, 保护区湿地有明显的丰水期和枯水期, 丰水期(4月—9月)内保护区除总面积不足4 km2的南山岛和矶山岛两个人居岛屿外其他草洲均被洪水淹没, 处于典型的湖相水文状态。枯水期内(10月—次年3月)湖水消退归入河道和一些碟形洼地, 区内大小河流纵横, 碟形湖泊星罗棋布, 不同高程洲滩相继出露, 整个三角洲地区呈现河、湖、洲交错的景观[30]。这种特殊的水文节律使得保护区内形成了大量土壤肥沃、水热条件好的洲滩和浅滩, 生长了丰富的湿生植物和水生植物, 多样性极其丰富, 其中以芦苇(Phragmites australis)、南荻(Triarrhena lutarioriparia)、薹草(Carex cinerascens)等为优势种[22, 32]。

|
| 图 1 样点在鄱阳湖湿地中的位置 Fig. 1 The position of sampling area in Poyang Lake Wetland |
本研究始于2016年11月中下旬。在调查鄱阳湖湿地植物分布格局的基础上, 在白沙湖内选择人为干扰相对较少、植物群落发育良好、离湖心水线大约500 m左右的洲滩上建立5块固定的试验样地, 样地之间大约间隔50 m左右(图 1)。
样地内的优势植物为芦苇、南荻、薹草等。在每个样地内选取一个点作为本次分解实验的5个重复样点(图 1)。分解模拟实验采用分解袋法。选择100目(孔径为0.15 mm)、规格为15 cm×20 cm的白色网孔的尼龙网袋作为分解袋。这种网袋既能够避免分解袋中分解残体的非分解损失, 也能保证分解作用不限制。
用于分解模拟实验的装置如图 2所示。将直径0.4 m, 高1 m的PVC管固定在每个实验监测样点, 埋入土壤中的深度为0.6 m, 并在PVC管内插入1 m高的竹竿, 用于固定分解袋。

|
| 图 2 植物分解监测实验装置 Fig. 2 Experimental device to perform the decomposition process monitoring |
2016年10月上旬在固定样方周围洲滩的上采集芦苇、南荻和薹草成熟叶片。将采集好的植物叶片带回实验室用去离子水冲洗干净, 剪成10 cm长小段后混合均匀(消除尺寸对分解的影响)装入牛皮纸信封内, 置于烘箱120℃杀青1 h后再60℃烘干72小时。每种植物分别取5.00 g烘干的叶片5个重复测其初始养分含量(表 2)。
| 芦苇Phragmites australis | 南荻Triarrhena lutarioriparia | 薹草Carex cinerascens | ||||||
| 平均值 | 标准差 | 平均值 | 标准差 | 平均值 | 标准差 | |||
| 总碳Total carbon/% | 43.843a | 1.258 | 43.078b | 2.308 | 42.825c | 1.286 | ||
| 总氮Total nitrogen/% | 3.893a | 0.564 | 1.763b | 0.174 | 2.211a | 0.162 | ||
| 总磷Total phosphorus/‰ | 0.062a | 0.009 | 0.051b | 0.008 | 0.073a | 0.002 | ||
| 碳氮比C/N ratio | 11.262a | 0.803 | 24.434b | 1.041 | 19.369a | 1.041 | ||
| 碳磷比C/P ratio | 711.676a | 10.185 | 845.904b | 11.823 | 590.209a | 20.378 | ||
| 氮磷比N/P ratio | 63.193a | 5.423 | 34.619b | 2.458 | 30.472a | 1.257 | ||
| 差异显著性使用Tukey检验。均值后的不同字母表示样本类型之间的显著差异(n=5, P < 0.05) | ||||||||
| Olson时间衰减模型 Olson decay Model |
k | R2 | T0.5/d | T0.95/d | |
| 芦苇Phragmites australis | R=98.09441e-0.00377t | 0.00377 | 0.84731 | 183.86 | 794.62 |
| 南荻Triarrhena lutarioriparia | R=94.99457e-0.00278t | 0.00278 | 0.87883 | 249.33 | 1077.60 |
| 薹草Carex cinerascens | R=98.83907e-0.001330t | 0.00330 | 0.89201 | 210.05 | 907.80 |
| 样品分解完成50%所需时间T0.5=-ln(0.5)/k, 完成95%分解所需时间T0.95=-ln(0.05)/k, k是分解速率常数 | |||||
再将剩下的叶片分别装入尼龙网袋中制作成分解袋。分解袋内的样品有3种形式:第一种是内装5.00 g的芦苇叶片(以下简称LW), 第二种内装5.00 g的南荻叶片(以下简称ND), 第三种是内装5.00 g的薹草叶片(以下简称TC)。将制作好的分解袋带到样地固定在预先设置的5套装置内。将样品袋用短绳绑在在插入泥土中的竹竿管底部(不破坏地表原有枯落物结构), 保证每个样品袋都充分地接触地表且相互不挤压。分别于实验开始后第15天、30天、60天、90天、120天和150天从各装置内取回分解袋。试验终止于4月中旬, 因为洪水淹没样地改变了分解环境的水文条件。
将取回的分解袋带回实验室, 除去泥土、藻类以及吸附物等杂物, 然后将所有样品装到有编号的牛皮信封中, 置于60℃烘箱中烘干至恒重, 称量其干物质残余量后再测定其中的总碳、总氮和总磷的含量。样品全碳和全氮含量使用元素分析仪(Vario Max CN Analyzer, Elementar Analysensysteme GmbH, Germany)测定;全磷使用电感耦合等离子体发射光谱仪(Inductively Coupled Plasma Optical Emission Spectrometer(ICP-OES), Optima 5300DV, Perkin-Elmer, America)测定。
1.3 数据处理与统计分析植物枯落物分解过程的评估参数用以下公式计算:
实测瞬时残留率(The remaining rate, Rt)计算公式[33]:
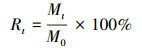
|
(1) |
式中, Rt表示t时刻残留率, Mt表示t时刻重量, M0表示初始重量, t表示分解时间。
瞬时衰减系数(The instantaneous loss rate, k)用Olson负指数衰减模型[34]计算:
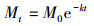
|
(2) |
式中, k表示t时刻瞬时分解速率参数, k值越大分解速度越快。
干物质残余率(y)用负指数衰减模型[35]levenberg Marquardt算法进行模拟:
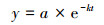
|
(3) |
式中, y为枯落物残留率, a为拟合参数, e为自然底数。
相对养分归还指数(relative return index, RRI)计算公式[33]:
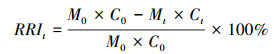
|
(4) |
式中, Ct为t时刻某元素的浓度, C0为某元素的初始浓度。CRRI表示碳的相对归还指数(carbon relative return index), NRRI表示氮积累指数(nitrogen relative return index), PRRI表示磷积累指数(phosphorus relative return index)。当RRI为正数时表示枯落物分解过程中元素发生了净释放, 当RRI为负数时则表示发生了净累积。
数据在Excel 2016中统计分析, 在Origin 9.0软件中制图, 在SPSS 19.0中进行单因素方差分析(ANOVA), Tukey′s honestly差异显著性检验以及Pearson相关性分析, 在Origin 9.0软件中进行单指数衰减模型拟合与制图。
2 结果与分析 2.1 植物枯落物分解速率及残留率的分异特征通过Olson负指数衰减模型Mt=M0e-kt计算出每个分解时间点的枯落物干物质瞬时衰减系数k, 结果表明:分解时间对芦苇、南荻和薹草枯落物k值的影响极其显著(LW: F=371.828, P < 0.0001; ND: F=132.889, P < 0.0001; TC: F=89.047, P < 0.0001)。
三种植物枯落物干物质k值都呈现出快速增大至最大值然后又迅速下降直至稳定的趋势(图 3)。其中芦苇k值在15 d内增加至最大值1.17, 在10 d到90 d内快速下降至0.49左右, 第90天后非常缓慢地变小直至稳定;南荻和薹草的k值在30 d左右增加至最大值, 最大值分别约为0.57和0.92, 在30—90 d内快速下降, 第90天后也非常缓慢地变小直至稳定(图 3)。

|
| 图 3 芦苇、南荻和薹草枯落物干物质分解速率及残留率动态拟合 Fig. 3 Dynamics of decay rate and remaining rate of dry mass of Phragmites australis, Triarrhena lutarioriparia, Carex cinerascens litter |
单因素方差检验发现, 在0—150 d内芦苇、南荻和薹草枯落物干物质k值的差异性极显著(10 d:F=148.642, P < 0.0001; 30 d: F=108.147, P < 0.0001; 60 d: F=9.068, P=0.005; 90 d: F=30.617, P < 0.0001; 120 d: F=22.768, P < 0.0001; 150 d: F=17.857, P < 0.0001)。此外, 在每个分解时间点上k值都是芦苇的最大, 薹草的次之, 南荻最小(图 3)。
在0—150 d内, 芦苇、南荻和薹草枯落物的干物质残留率呈现逐渐减小趋势(图 3)。分解时间对三种植物枯落物的干物质残留率的影响极其显著(LW: F=863.591, P < 0.0001; ND: F=927.086, P < 0.0001; TC: F=75.488, P < 0.0001)。
分解0—150 d内, 三种植物枯落物的干物质残留率也具有极显著的差异性(10 d:F=156.198, P < 0.0001; 30 d: F=117.409, P < 0.0001; 60 d: F=9.573, P=0.004; 90 d: F=29.77, P < 0.0001; 120 d: F=23.302, P < 0.0001; 150 d: F=20.386, P < 0.0001)。此外, 在0—150 d内的干物质残留率总是南荻最大, 薹草次之, 芦苇最低。分解150 d后南荻、薹草和芦苇枯落物的干物质残留率依次约为67.99%、60.88%和56.57%(图 3)。
将干物质残留率用Olson负指数衰减模型的levenberg Marquardt算法进行拟合, 发现拟合效果非常好, 拟合系数R2均在0.84以上, 拟合方程的各参数如图 3与表 2所示。通过拟合方程预测出芦苇、南荻、薹草枯落物分解完成50%所需的时间大约依次为184 d、249 d和210 d, 分解完成95%所需的时间依次为795 d(约2.18 a)、1078 d(约2.95 a)和908 d(约2.49 a), 并且3种样品的平均分解速率差异极显著(P < 0.0001), 分别约为0.00377、0.00278和0.00330.
2.2 植物枯落物碳、氮、磷释放动态分异特征在整个分解0—150 d内芦苇、南荻和薹草枯落物的碳元素都表现为净释放模式(CRRI>0), 未出现积累的情况(图 4)。不同分解时间段内三种植物枯落物CRRI值差异性极显著(10 d:F=185.613, P < 0.0001; 30 d: F=23.927, P < 0.0001; 60 d: F=13.319, P=0.003; 90 d: F=104.349, P < 0.0001; 120 d: F=53.742, P < 0.0001; 150 d: F=103.095, P < 0.0001)。在每个分解时间都是芦苇的CRRI值最高, 薹草次之, 南荻最小。随着分解的进行三种植物枯落物的CRRI一直在持续增大, 直到到150 d时芦苇、薹草和南荻枯落物的CRRI值依次增大至约为57.44%、41.75%和34.58%(图 4)。

|
| 图 4 枯落物分解过程碳、氮、磷相对归还指数动态 Fig. 4 Dynamics of RRIs of Phragmites australis, Triarrhena lutarioriparia, Carex cinerascens litter |
在整个分解0—150 d内芦苇和薹草枯落物的氮元素都表现为净释放模式(NRRI>0), 未出现积累的情况, 而南荻的氮元素一直表现为净积累模式(NRRI < 0), 未出现释放的情况(图 4)。分解0—150 d 3种植物枯落物的NRRI值差异性极其显著(10 d:F=39.472, P < 0.0001; 30 d: F=38.248, P < 0.0001; 60 d: F=71.223, P < 0.0001; 90 d: F=178.993, P < 0.0001; 120 d: F=47.074, P < 0.0001; 150 d: F=33.614, P < 0.0001)。每个分解时间段的NRRI值均为芦苇最高, 薹草次之, 南荻最小。随着分解的进行NRRI值并不是一直都升高, 而是波动起伏的。芦苇的的NRRI值波幅度很小, 整体呈现出一直增大的趋势。薹草NRRI值在0—10 d内快速升高, 而15—30 d内快速减小, 在30—150 d内一直以非常缓慢的速度增大。南荻的NRRI在0—60 d内迅速下降至最小值约-44.53%, 在60—120 d内缓慢上升至约-18.01%, 在150 d时有减小至约-22.66%, 南荻的NRRI在0—150 d内虽上下波动起伏, 但均为负值(图 4)。
在整个分解0—150 d内芦苇、南荻和薹草枯落物的磷元素都表现为净释放模式(PRRI>0), 未出现积累的情况(图 4)。不同分解时间段内三种植物枯落物PRRI值差异性极显著(10 d:F=25.324, P < 0.0001; 30 d: F=120.071, P < 0.0001; 60 d: F=108.879, P < 0.0001; 90 d: F=339.118, P < 0.0001; 120 d: F=159.296, P < 0.0001; 150 d: F=94.718, P < 0.0001)。3种植物枯落物的PRRI值相差不大, 但是每个分解时间段内PRRI值都是芦苇最高, 薹草次之, 南荻最小。随着分解的进行三种植物枯落物的PRRI值都持续升高, 都在0—30 d内快速上升至最大值后再逐渐缓慢增大, 到150 d时, 芦苇、薹草、南荻枯落物的PRRI值分别依次约为88.91%、85.63%和79.27%(图 4)。
2.3 植物枯落物碳、氮、磷化学计量比分异特征芦苇、南荻和薹草枯落物分解过程中的碳、氮、磷化学计量比动态变化十分明显(图 5)。

|
| 图 5 枯落物分解过程碳、氮、磷化学计量比动态 Fig. 5 Dynamics of stoichiometric ratio of Phragmites australis, Triarrhena lutarioriparia, Carex cinerascens litter |
在分解的起始阶段, C/N比值的大小顺序为:南荻(约24.43)>薹草(约19.37)>芦苇(约11.26)。但是随着分解的进行, 在15—150 d内C/N的大小顺序变为:薹草>南荻>芦苇, 并且芦苇C/N比一直比其他两种低很多, 而薹草和南荻C/N比差值并不大。不同分解时间段内三种植物枯落物C/N比值差异性显著(P < 0.05)。从植物种类来看, 芦苇C/N比在一直在缓慢下降, 在150 d时达到最小值(约9.85), 比初始值低约12.52%;南荻的C/N在0—60 d持续快速降低到最小值(约12.83), 然后缓慢上升, 到150时约为13.05, 比初始值低约46.58%薹草的C/N比在0—60 d内小幅度增大, 在60 d时达到最小值(约14.98), 而后逐渐缓慢增加, 到150 d时约为15.96, 比初始值低约17.60% (图 5)。
在分解的起始阶段, C/P比值的大小顺序为:南荻(约845.60)>芦苇(约711.68)>薹草(约590.21)。但是随着分解的进行, 在15 d以及90—150 d内C/P的大小顺序变为:芦苇>南荻>薹草, 并且三者之间差值并不大;在30—60 d内C/P的大小顺序变为:芦苇>薹草>南荻, 并且芦苇C/N比一直比其他两种高很多, 而薹草和南荻C/N比差值并不大。不同分解时间段内三种植物枯落物C/N比值差异性显著(P < 0.05)。从植物种类来看, 芦苇C/P比在0—30 d内迅速增大至最大值(约为3755.58), 然后缓慢减小, 直至第150天时仍比初始值高约284.93%;南荻的C/P在0—30 d内快速增加, 在30—120 d内持续缓慢增大到最大值(约3091.29), 然后缓慢下降, 到150 d时仍比初始值高约215.65%;薹草的C/P比在0—30 d内快速增加, 在30—120 d内持续缓慢增大到最大值(约2694.47), 而后逐渐缓慢下降, 到150 d仍比初始值高约316.96% (图 5)。
在分解的起始阶段, N/P比值的大小顺序为:芦苇(约63.19)>南荻(约34.62)>薹草(约30.47)。随着分解的进行, 在0—150 d内N/P的大小顺序没有发生改变。芦苇N/P比一直比其他两种高很多, 而薹草和南荻N/P比差值并不大。不同分解时间段内三种植物枯落物N/P比值差异性显著(P < 0.05)。从植物种类来看, 芦苇N/P比在0—30 d内迅速增大至最大值(约为333.44), 然后缓慢减小, 直至第150天时仍比初始值高约348.55%;南荻的N/P在0—30 d内快速增加, 在30—120 d内持续缓慢增大到最大值(约221.02), 然后缓慢下降, 到150 d时仍比初始值高约419.91%;薹草的N/P比在0—30 d内快速增加, 在30—90 d内持续缓慢增大到最大值(约176.31), 而后逐渐缓慢下降, 到150 d仍比初始值高约428.98% (图 5)。
3 讨论通过在鄱阳湖湿地展开野外定位观测研究, 对湿地不同优势植物芦苇、南荻和薹草枯落物进行150 d的分解模拟实验, 发现3种植物枯落物干物质分解速率k值及残留率都存在极显著的差异性(图 3), 这表明不同物种对枯落物的分解有不同程度的影响。本研究的分解模拟实验是在相同的非生物(光、温、水)条件下、相同的土壤类型和相同的试验持续时间内进行的, 所以植物枯落物分解的差异性主要与不同物种枯落物之间的化学组成与其计量比的差异有关(表 1), 而不是与小气候[11]、水分条件[7]及土壤微生物活性[36]的差异有关。本研究中三种植物枯落物的碳、氮和磷含量及其化学计量比在0—150 d内差异性极显著。我们的研究结果与许多前人在这方面的研究结果是一致的[2, 11-12, 19, 36], 认为在其他条件相同时, 枯落物的质量, 如木质素含量、碳、氮和磷等养分浓度及其化学计量比, 特别是氮和磷的浓度与分解速率有着非常密切的关系[37-39]。在通常情况下, 木质素等不容易分解的成含量高以及低氮、低磷的枯落物分解速率很慢[11, 40], 反之, 较高营养含量和较低木质素含量的枯落物往往能更快更容易地被分解[41]。一般在湿地生态系统中植物叶片C:N含量或C:P含量越高, 其分解速率越慢[42]。本研究中芦苇叶片的碳和氮初始含量最高、薹草次之, 而南荻的碳和氮含量最低, 南荻初始C:N和C:P最高, 而芦苇C:N最低(表 1), 因此, 在每个分解时间点上的分解速率都是芦苇的最大, 薹草的次之, 南荻最小(图 3)。
3种植物枯落物分解速率k都呈现出0—15 d内(芦苇)或0—30 d内(南荻和薹草)快速增大至最大值然后又迅速下降直至90 d以后趋于稳定的趋势(图 3)。这是可能由于鄱阳湖湿地环境中水分比较充足[43], 分解前期植物枯落物中可溶解成分快速淋溶, 水溶成分和非木质素碳水化合物优先分解, 且分解速率非常快[44], 而分解中后期期已木质化的未分解碳水化合物与原有木质素在微生物作用下同时降解, 这个过程就比较缓慢, 直到最后枯落物的木质素含量趋于稳定, 残余物质逐渐腐殖化, 这时候分解速率非常小也很稳定[45]。
3种植物枯落物分解过程中的碳、氮和磷归还指数及其化学计量比都存在极显著的差异性、并且碳、氮和磷归还指数的值都表现为芦苇的最高, 薹草次之, 南荻最小(图 4), 这与分解速率大小规律是高度一致的。这进一步说明枯落物的质量对枯落物分解过程中的养分释放也有十分重要的影响[46], 植物叶片枯落物C:N及C:P对分解过程中碳、氮和磷释放量具有显著的调控作用[39]。本研究中三种植物枯落物的CRRI一直在持续增大, 并且前期增加速度快, 后期增加速度较慢(图 4)。这与前人的研究结果基本一致[33, 47]。一般情况下, CRRI变化整体包含前期快速增加和后期缓慢增加两个过程[48], 在前期淋溶过程中溶解性碳水化合物快速发生淋溶, CRRI会快速的增加, 而后期受外部环境因子的激发作用或者外源营养元素输入等因素的影响[15, 44], 土壤微生物数量变多、活力增强, 分解残留物中剩余木质素、纤维素和单宁等难分解物质开始逐渐的被微生物所利用, 此时CRRI的增加速度开始减缓甚至开始减小[44]。
3种植物枯落物的NRRI表现出不相同的趋势。在整个分解过程芦苇和薹草NRRI都正值, 表现为波浪式缓慢增加, 都是氮的净释放模式。而南荻的NRRI在整个分解过程中均为负值, 呈现先迅速下降再缓慢增加的趋势, 都是氮的净积累模式(图 4)。这是因为淋溶过程氮元素会由于含氮盐的流失而释放, 但是在微生物在分解大分子蛋白质时会因对氮需求的增加而导致氮积累[49], NRRI增加速度会减缓甚至会大幅下降[50], 因此NRRI会表现出波动起伏的情况。另外, 分解过程中氮元素净积累还是净释放还与初始状态下的C:N和氮元素浓度有关[51-52], 当氮浓度高于2.0%或C:N在25—30之间时, 氮元素开始释放, 当氮浓度过低时则会发生氮的积累。本研究中, 南荻叶片氮浓度约为1.763%, 而薹草和芦苇叶片氮浓度均大于2.0%(表 1)。因此, 芦苇和薹草表现为氮释放, 而南荻表现为氮积累。
本研究中3种植物枯落物分解过程中PRRI都表现为在0—30 d内快速上升至最大值后再逐渐缓慢增大直至稳定的趋势(图 4)。这与前人的研究结果基本一致。由于植物组织中的磷主要以磷酸根或化合物的形式存在而容易发生淋溶损失的原因[53], 分解初期磷的淋溶作用比较强, 枯落物中磷归还速度的非常快, 当磷释放到一定程度时浓度较低时归还速度就会减缓并趋于稳定[54]。而在Kuehn和Suberkropp[21]的研究发现在灯芯草(Juncus effuses)的分解过程中, 枯落物中磷元素浓度略有增加。这种研究结果的不一致主要是因为本研究所选择三种湿地植物叶片枯落物中磷元素的浓度远远高于灯芯草叶片中磷元素的浓度。在鄱阳湖湿地生态系统中主要优势植物的分解过程并不受到磷元素限制的影响, 植物枯落物残存的磷元素足以供给微生物用于自身的生长, 并且还会把多余的磷元素释放到环境中[7, 15, 29]。
4 结论本研究在江西省鄱阳湖湿地展开野外定位观测实验, 采用分解法对3种优势湿地植物芦苇、南荻和薹草枯落物分解过程进行了研究。表明在气候和立地环境条件相同的情况下, 植物种类及基质物质量对枯落物分解及其养分释放有很强的调控作用。主要结论如下:
(1) 芦苇、南荻和薹草枯落物的分解速率、干物质残留率以及碳、氮和磷的相对归还指数都有极显著的差异性;(2)在0—150 d内分解速率都是芦苇的最大, 薹草的次之, 南荻最小。用Olson负指数衰减模型拟合方程预测芦苇、南荻、薹草枯落物分解完成50%所需的时间大约依次为184 d、249 d和210 d, 分解完成95%所需的时间依次为795 d、1078 d和908 d;(3)三种植物枯落物C和P在分解过程中都表现出净释放模式, 而南荻枯落物的N一直表现为净积累模式。芦苇分解过程中的营养释放作用最强, 而南荻群落对氮的吸收和富集效应最强。
开展枯水期鄱阳湖湿地优势植物分解过程的研究, 不仅有助于深入理解湖泊湿地优势植物枯落物在碳、氮、磷生物地球化学循环过程的作用, 为鄱阳湖水文节律变化背景下草洲营养元素释放过程的预测提供数据支持, 而且有利于完善鄱阳湖生态安全预警机制, 并为草洲管理提供科学认识。今后的研究应考虑不同物种枯落物混合时的分解过程以及分解过程中的微生物因素, 以便能揭示植物群落物种多样性及微生物活动在湿地生物地球化学循环中的调控作用机制。
致谢: 感谢江西鄱阳湖南矶湿地国家级自然保护区管理局及相关工作人员对本研究野外工作的支持;感谢南昌大学生命科学研究院流域生态学研究所的金松斌老师、兰志春博士和沈瑞昌博士在室内试验提供的帮助;感谢南昌大学张欢老师、吴申浩同学和王鑫同学、中国科学院地理科学与资源研究所博士生饶滴滴同学对野外样地布设、取样以及制图的帮助。| [1] |
Schlesinger W H. Biogeochemistry: An Analysis of Global Change. San Diego: Academic Press, 1991.
|
| [2] |
Hoorens B, Aerts R, Stroetenga M. Does initial litter chemistry explain litter mixture effects on decomposition?. Oecologia, 2003, 137(4): 578-586. DOI:10.1007/s00442-003-1365-6 |
| [3] |
Sun Z G, Mou X J, Sun W L. Potential effects of tidal flat variations on decomposition and nutrient dynamics of Phragmites australis, Suaeda salsa and Suaeda glauca litter in newly created marshes of the Yellow River Estuary, China. Ecological Engineering, 2016, 93: 175-186. DOI:10.1016/j.ecoleng.2016.05.024 |
| [4] |
Berg B, Laskowski R. Litter Decomposition: A Guide to Carbon and Nutrient Turnover. London: Academic Press, 2006.
|
| [5] |
Roehm C L. Respiration in wetland ecosystems//Del Giorgio P A, Williams P J L B, eds. Respiration in Aquatic Ecosystems. Oxford: Oxford University Press, 2005.
|
| [6] |
Del Giorgio P A, Le B Williams P J. Respiration in Aquatic Ecosystems. Oxford: Oxford University Press, 2005.
|
| [7] |
Zhang Q J, Zhang G S, Yu X B, Liu Y, Xia S X, Ya L, Hu B H, Wan S X. Effect of ground water level on the release of carbon, nitrogen and phosphorus during decomposition of Carex. cinerascens kükenth in the typical seasonal floodplain in dry season. Journal of Freshwater Ecology, 2019, 34(1): 305-322. DOI:10.1080/02705060.2019.1584128 |
| [8] |
Fioretto A, Di Nardo C, Papa S, Fuggi A. Lignin and cellulose degradation and nitrogen dynamics during decomposition of three leaf litter species in a Mediterranean ecosystem. Soil Biology and Biochemistry, 2005, 37(6): 1083-1091. DOI:10.1016/j.soilbio.2004.11.007 |
| [9] |
Serna A, Richards J H, Scinto L J. Plant decomposition in wetlands:effects of hydrologic variation in a re-created everglades. Journal of Environmental Quality, 2013, 42(2): 562-572. DOI:10.2134/jeq2012.0201 |
| [10] |
Mitsch W J, Gosselink J G. Wetlands. 5th ed. Hoboken, New Jersey: John Wiley & Sons, Inc, 2015.
|
| [11] |
Aerts R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems:a triangular relationship. Oikos, 1997, 79(3): 439-449. DOI:10.2307/3546886 |
| [12] |
Vivanco L, Austin A T. Tree species identity alters forest litter decomposition through long-term plant and soil interactions in Patagonia, Argentina. Journal of Ecology, 2008, 96(4): 727-736. |
| [13] |
Vitousek P M. Nutrient cycling and limitation: Hawai'i as a model system. Princeton, NJ: Princeton University Press, 2004.
|
| [14] |
Zhang G S, Yu X B, Xu J, Duan H L, Rafay L, Zhang Q J, Li Y, Liu Y, Xia S X. Effects of environmental variation on stable isotope abundances during typical seasonal floodplain dry season litter decomposition. Science of the Total Environment, 2018, 630(1): 1205-1215. |
| [15] |
张全军, 张广帅, 万松贤, 刘宇, 夏少霞, 李雅, 许策, 于秀波. 鄱阳湖植食越冬候鸟粪便对洲滩湿地薹草枯落物分解过程及碳、氮、磷释放的影响. 湖泊科学, 2019, 31(3): 814-824. |
| [16] |
Swift M J, Heal O W, Anderson J M. Decomposition in Terrestrial Ecosystems. Oxford: Blackwell, 1979.
|
| [17] |
Cornelissen J H C, Pérez-Harguindeguy N, Díaz S, Grime J P, Marzano B, Cabido M, Vendramini F, Cerabolini B. Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents. New Phytologist, 1999, 143(1): 191-200. DOI:10.1046/j.1469-8137.1999.00430.x |
| [18] |
Gessner M O. Breakdown and nutrient dynamics of submerged Phragmites shoots in the littoral zone of a temperate hardwater lake. Aquatic Botany, 2000, 66(1): 9-20. DOI:10.1016/S0304-3770(99)00022-4 |
| [19] |
Petraglia A, Cacciatori C, Chelli S, Fenu G, Calderisi G, Gargano D, Abeli T, Orsenigo S, Carbognani M. Litter decomposition:effects of temperature driven by soil moisture and vegetation type. Plant and Soil, 2019, 435(1/2): 187-200. |
| [20] |
戴雪, 何征, 万荣荣, 杨桂山. 近35年长江中游大型通江湖泊季节性水情变化规律研究. 长江流域资源与环境, 2017, 26(1): 118-125. |
| [21] |
Kuehn K A, Suberkropp K. Decomposition of standing litter of the freshwater emergent macrophyte Juncus effusus. Freshwater Biology, 1998, 40(4): 717-727. DOI:10.1046/j.1365-2427.1998.00374.x |
| [22] |
张全军, 于秀波, 钱建鑫, 熊挺. 鄱阳湖南矶湿地优势植物群落及土壤有机质和营养元素分布特征. 生态学报, 2012, 32(12): 3656-3669. |
| [23] |
葛刚, 赵安娜, 钟义勇, 吴志强. 鄱阳湖洲滩优势植物种群的分布格局. 湿地科学, 2011, 9(1): 19-25. DOI:10.3969/j.issn.1673-3290.2011.01.005 |
| [24] |
李冰, 杨桂山, 王晓龙, 万荣荣. 鄱阳湖典型洲滩植物物种多样性季节动态特征. 土壤, 2016, 48(2): 298-305. |
| [25] |
张广帅, 于秀波, 刘宇, 张欢, 张全军, 李雅, 段后浪. 鄱阳湖碟形湖泊植物分解和水位变化对水体碳、氮浓度的叠加效应. 湖泊科学, 2018, 30(3): 668-679. |
| [26] |
Zhang G S, Yu X B, Gao Y, Li Y, Zhang Q J, Liu Y, Rao D D, Lin Y M, Xia S X. Effects of water table on cellulose and lignin degradation of Carex Cinerascens in a large seasonal floodplain. Journal of Freshwater Ecology, 2018, 33(1): 311-325. DOI:10.1080/02705060.2018.1459324 |
| [27] |
聂兰琴, 吴琴, 尧波, 付姗, 胡启武. 鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征. 生态学报, 2016, 36(7): 1898-1906. |
| [28] |
付姗, 吴琴, 郑艳明, 胡启武. 鄱阳湖沙山植物叶片与土壤C:N、C:P沿沙化梯度分布特征. 长江流域资源与环境, 2015, 24(3): 447-454. DOI:10.11870/cjlyzyyhj201503014 |
| [29] |
郑艳明, 尧波, 吴琴, 胡斌华, 胡启武. 鄱阳湖湿地两种优势植物叶片C、N、P动态特征. 生态学报, 2013, 33(20): 6488-6496. |
| [30] |
刘信中, 胡斌华. 江西南矶山湿地自然保护区综合科学考察. 北京: 中国林业出版社, 2005.
|
| [31] |
《鄱阳湖研究》编委会.鄱阳湖研究.上海: 上海科学技术出版社, 1988.
|
| [32] |
Wang Y Y, Yu X B, Zhang L, Lei G C. Seasonal variability in baseline δ15N and usage as a nutrient indicator in Lake Poyang, China. Journal of Freshwater Ecology, 2013, 28(3): 365-373. DOI:10.1080/02705060.2013.763296 |
| [33] |
Zhao Q Q, Bai J H, Liu P P, Gao H F, Wang J J. Decomposition and carbon and nitrogen dynamics of Phragmites australis litter as affected by flooding periods in coastal wetlands. Clean:Soil, Air, Water, 2015, 43(3): 441-445. |
| [34] |
Olson J S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 1963, 44(2): 322-331. |
| [35] |
Wider R K, Lang G E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology, 1982, 63(6): 1636-1642. |
| [36] |
Chapman S K, Newman G S. Biodiversity at the plant-soil interface:microbial abundance and community structure respond to litter mixing. Oecologia, 2010, 162(3): 763-769. |
| [37] |
Tessier J T, Dudley D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. Journal of Applied Ecology, 2003, 40(3): 523-534. |
| [38] |
Crawford E R, Day F P, Atkinson R B. Influence of environment and substrate quality on root decomposition in naturally regenerating and restored Atlantic white cedar wetlands. Wetlands, 2007, 27(1): 1-11. |
| [39] |
Enríquez S, Duarte C M, Sand-Jensen K. Patterns in decomposition rates among photosynthetic organisms:the importance of detritus C:N:P content. Oecologia, 1993, 94(4): 457-471. |
| [40] |
Day Jr F P. Litter decomposition rates in the seasonally flooded great dismal swamp. Ecology, 1982, 63(3): 670-978. |
| [41] |
dos Santos Fonseca A L, Bianchini Jr I, Pimenta C M M, Soares C B P, Mangiavacchi N. The flow velocity as driving force for decomposition of leaves and twigs. Hydrobiologia, 2013, 703(1): 59-67. |
| [42] |
Liao C Z, Luo Y Q, Fang C M, Chen J K, Li B. Litter pool sizes, decomposition, and nitrogen dynamics in Spartina alterniflora-invaded and native coastal marshlands of the Yangtze Estuary. Oecologia, 2008, 156(3): 589-600. |
| [43] |
张广帅, 于秀波, 张全军, 李雅, 刘宇, 段后浪. 鄱阳湖湿地土壤微生物群落结构沿地下水位梯度分异特征. 生态学报, 2018, 38(11): 3825-3837. |
| [44] |
Davis III S E, Corronado-Molina C, Childers D L, Day Jr J W. Temporally dependent C, N, and P dynamics associated with the decay of Rhizophora mangle L. leaf litter in oligotrophic mangrove wetlands of the southern Everglades. Aquatic Botany, 2003, 75(3): 199-215. |
| [45] |
Kalbitz K, Kaiser K, Bargholz J, Dardenne P. Lignin degradation controls the production of dissolved organic matter in decomposing foliar litter. European Journal of Soil Science, 2006, 57(4): 504-516. |
| [46] |
Grosso F, Bååth E, De Nicola F. Bacterial and fungal growth on different plant litter in Mediterranean soils:effects of C/N ratio and soil pH. Applied Soil Ecology, 2016, 108: 1-7. |
| [47] |
Debusk W F, Reddy K R. Litter decomposition and nutrient dynamics in a phosphorus enriched everglades marsh. Biogeochemistry, 2005, 75(2): 217-240. |
| [48] |
史学军, 潘剑君, 陈锦盈, 杨志强, 张黎明, 孙波, 李忠佩. 不同类型凋落物对土壤有机碳矿化的影响. 环境科学, 2009, 30(6): 1832-1837. |
| [49] |
Aber J D, Melillo J M. Litter decomposition:measuring relative contributions of organic matter and nitrogen to forest soils. Canadian Journal of Botany, 1980, 58(4): 416-421. |
| [50] |
Wu H T, Lu X G, Yang Q, Jiang M, Tong S Z. Early-stage litter decomposition and its influencing factors in the wetland of the Sanjiang Plain, China. Acta Ecologica Sinica, 2007, 27(10): 4027-4035. |
| [51] |
Kröger R, Holland M M, Moore M T, Cooper C M. Plant senescence:a mechanism for nutrient release in temperate agricultural wetlands. Environmental Pollution, 2007, 146(1): 114-119. |
| [52] |
Jacob M, Weland N, Platner C, Schaefer M, Leuschner C, Thomas F M. Nutrient release from decomposing leaf litter of temperate deciduous forest trees along a gradient of increasing tree species diversity. Soil Biology and Biochemistry, 2009, 41(10): 2122-2130. |
| [53] |
温达志, 魏平, 张佑昌, 孔国辉. 鼎湖山南亚热带森林细根分解干物质损失和元素动态. 生态学杂志, 1998(2): 1-6, 31-31. |
| [54] |
Wrubleski D A, Murkin H R, Van Der Valk A G, Nelson J W. Decomposition of emergent macrophyte roots and rhizomes in a northern prairie marsh. Aquatic Botany, 1997, 58(2): 121-134. |