 2020, Vol. 40
2020, Vol. 40文章信息
- 李玉凤, 黄婧, 马姜明, 莫燕华
- LI Yufeng, HUANG Jing, MA Jiangming, MO Yanhua
- 桂林喀斯特石山50种常见植物叶片光合特性
- The leaf photosynthetic characteristics of 50 common plants on karst rocky mountain in Guilin
- 生态学报. 2020, 40(23): 8649-8659
- Acta Ecologica Sinica. 2020, 40(23): 8649-8659
- http://dx.doi.org/10.5846/stxb201909302058
-
文章历史
- 收稿日期: 2019-09-30
- 网络出版日期: 2020-10-29





2. 珍稀濒危动植物生态与环境保护教育部重点实验室, 桂林 541006
2. Key Laboratory of Ecology of Rare and Endangered Species and Environmental Protection, Ministry of Education, Guilin 541006, China
植物生理生态特性与叶片特征相联系, 主要体现于叶片的光合作用[1]。光合作用是植物生长发育的基础, 光合指标可反映植物的生理特性[2]。Aarea, Amass, Gs, WUE, Ci/Ca和Tr是植物叶片的主要光合生理特性因子, 综合反映植被生长发育、物质生产与水分消耗关系[3]。单位面积最大净光合速率(Aarea)和单位质量最大净光合速率(Amass)可用来表示叶片光合能力的强弱[4]。植物气孔导度(Gs)可有效表示植物生理状况以及对水分的敏感性, 其对水分胁迫的响应规律对于衡量植物抗旱性具有重要指示作用[5]。水分利用效率(WUE)是评价缺水条件下植物生长适宜程度的综合指标[6], 用于表征树种抗旱性能和水分利用特征[7]。胞间CO2浓度(Ci)是反映大气输入、光合利用能力及光呼吸的CO2动态平衡的瞬间浓度, 一般与Gs及Ca相关, 表示植物对CO2的利用情况[8]。蒸腾速率(Tr)可间接反映植物水分利用效率[9], 其强弱在一定程度上体现了植物适应逆境及调节水分平衡的能力[10]。植物叶片的光合生理特性体现了植物的生长策略和资源利用方式, 植物与环境长期相互作用, 不断适应外界环境[11]。目前, 有关光合特性的研究主要包括:光合特性与环境因子的关系[12], 光合特性的影响因素, 包括干旱胁迫[13]、遮阴[14]、水分胁迫[15], 高温胁迫[16]等, 对于桂林喀斯特地区植物叶片光合特性等方面的研究比较少。
广西喀斯特地区石漠化土地总面积为1.53万km2, 占全区国土面积的6.5%, 桂林岩溶区石漠化问题凸显, 由碳酸盐岩溶蚀形成的岩溶平原(残山平原)、峰林平原、峰丛洼(谷)地为主的典型岩溶地貌区[17], 生态系统较为脆弱, 植被恢复在岩溶区石漠化治理和生态重建中发挥关键作用[18]。本研究对桂林喀斯特石山生境中常见的50种植物叶片光合特性指标进行分析, 旨在进一步阐明喀斯特石山50种常见植物光合特性的差异以及光合特性之间的相互关系, 有助于进一步理解物种的资源利用方式和对喀斯特石山生境的适应策略。以期为加快植被的恢复演替进程提供相应的理论基础, 最终为该地区的植被恢复与重建过程中的物种选择提供参考[19]。
1 研究地区与方法 1.1 研究地概况研究地区位于广西壮族自治区桂林市喀斯特石山, 地处广西东北部, 109°36′—111°29′ E, 24°15′—26°23′ N。属于山地丘陵地区, 为典型的“喀斯特”地貌, 气候属于中亚热带湿润季风气候, 海拔100—500 m。年平均气温18.9℃, 年平均日照时数1670 h。最冷的1月份平均气温15.6℃, 最热的8月份平均气温23℃;全年无霜期300d;年平均降雨量为1949.5 mm, 降雨量年分配不均, 秋、冬季干燥少雨;年平均蒸发量为1490—1905 mm。
1.2 研究方法于生长季6—9月开展试验, 在野外天气晴朗的9:00—11:00时段进行。采用便携式光合仪(LI-6400, LI-COR, Nebraska, USA), 选择生长健壮、长势一致、同一叶位的成熟叶片, 且具有代表性的植株, 取枝条生长良好的向阳中上部叶片进行光合指标的测定。每株测定3片叶片, 每片叶片重复测定3次, 结果取平均值进行分析。测定时, 首先进行充分的光合诱导, 设置光强为1500 μmol m-2 s-1, 用小钢瓶控制叶室CO2浓度在380 μmol/mol测定前让叶片充分照光。测定期间相对湿度60%—65%, 叶片温度28—30℃, 叶片到空气的水气压差为1.0—1.2 kPa。待数据稳定后记录Aarea、Gs、Tr和Ci。计算方法如下:
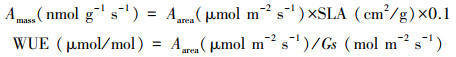
|
采用SPSS 23.0软件处理叶片光合特性指标的基本描述统计量, Pearson相关分析和主成分分析以及聚类分析, R进行主成分排序。
2 结果与分析 2.1 叶片光合特性数量特征由表 1可知, 桂林喀斯特石山生境50种常见植物叶片Aarea含量在(1.97±0.17)—(17.43±0.21) μmol m-2 s-1, 平均值为8.35 μmol m-2 s-1, 密花树(Myrsine seguinii)叶片Aarea含量显著小于其他49种植物, 铜钱树(Paliurus hemsleyanus)叶片Aarea含量显著大于其他49种植物。叶片Amass含量在(11.46±0.96)—(370.13±50.73) nmol g-1 s-1, 平均值为110.98 nmol g-1 s-1, 密花树叶片Amass含量显著小于其他49种植物, 薄叶鼠李(Rhamnus leptophylla)叶片Amass含量显著大于其他49种植物。叶片Gs含量在(0.01±0.01)—(0.24±0.02) mol m-2 s-1, 平均值为0.10 mol m-2 s-1, 密花树叶片Gs含量显著小于其他49种植物, 紫薇叶片Gs含量显著大于其他49种植物。叶片WUE含量在(58.84±1.64)—(167.03±11.35) μmol/mol, 平均值为94.84 μmol/mol, 桂花(Osmanthus fragrans)叶片WUE含量显著小于其他49种植物, 菜豆树(Radermachera sinica)叶片WUE含量显著大于其他49种植物。叶片Ci/Ca含量在(0.31±0.04)—(0.72±0.01), 平均值为0.57, 菜豆树叶片Ci/Ca含量显著小于其他49种植物, 桂花叶片Ci/Ca含量显著大于其他49种植物。叶片Tr含量在(0.23±0.02)—(6.46±0.60) mmol m-2 s-1, 平均值为2.37 mmol m-2 s-1, 密花树叶片Tr含量显著小于其他49种植物, 马甲子(Paliurus ramosissimus)叶片Tr含量显著大于其他49种植物。方差分析表明, 不同物种之间在Aarea, Amass, Gs, WUE, Ci/Ca和Tr之间存在显著差异。
| 物种 Plant species |
单位面积最大净光合速率 The greatest photosynthetic rate per unit areaAarea/(μmol m-2 s-1) |
单位质量最大净光合速率 The most photosynthetic rate per unit massAmass/(nmol g-1 s-1) |
气孔导度 Stomatal conductanceGs/(mol m-2 s-1) |
水分利用效率 Water utilization efficiencyWUE/(μmol/mol) |
Ci/Ca Intercellular/environmental CO2 concentration | 蒸腾速率 Transpiration rateTr/(mmol m-2 s-1) |
| 1.火棘 Pyracantha fortuneana | 11.24±0.71mnopqr | 116.79±8.71ijkl | 0.19±0.01mno | 59.87±3.23a | 0.68±0.01nopqr | 4.17±0.20ij |
| 2.蔓胡颓子 Elaeagnus glabra | 8.81±0.79fghijklmn | 97.80±7.56ghijkl | 0.05±0.01abcd | 161.60±8.15no | 0.45±0.01bcd | 0.82±0.05ab |
| 3.紫薇 Lagerstroemia indica | 17.11±2.23s | 204.51±27.61pq | 0.24±0.02p | 69.53±3.88abcd | 0.62±0.02ijklmnopqr | 4.86±0.33jk |
| 4.矮小天仙果 Ficus erecta | 4.02±0.41abcd | 128.02±12.78jklmn | 0.03±0.01abc | 145.67±11.53mno | 0.34±0.05a | 0.93±0.14ab |
| 5.了哥王 Wikstroemia indica | 4.91±1.42abcde | 53.98±15.53abcdefg | 0.06±0.02abcde | 82.67±7.85abcdefghij | 0.67±0.03mnopqr | 0.92±0.30ab |
| 6.小叶女贞 Ligustrum quihoui | 6.23±0.61cdefgh | 147.13±14.77lmno | 0.09±0.01defghi | 71.07±0.79abcdef | 0.69±0.01opqr | 1.33±0.10bc |
| 7.小花扁担杆 Grewia bilobavar. parviflora | 11.56±0.88mnopqr | 172.93±10.72mnop | 0.17±0.02klmn | 70.46±5.12abcde | 0.64±0.02lmnopqr | 3.49±0.32efghi |
| 8.红背山麻杆 Alchornea trewioides | 10.88±1.88lmnopqr | 146.85±35.99lmno | 0.12±0.01fghijk | 89.50±5.02bcdefghij | 0.59±0.03ghijklmnop | 2.09±0.25cd |
| 9.白皮乌口树 Tarenna depauperata | 9.89±1.07ijklmnopq | 73.31±8.96bcdefghij | 0.12±0.01ghijk | 82.59±8.72abcdefghij | 0.57±0.04efghijklmn | 3.80±0.24hi |
| 10.黄荆 Vitex negundo | 12.95±1.33qr | 237.97±5.62q | 0.14±0.01jklm | 89.89±2.65bcdefghij | 0.58±0.01efghijklmn | 2.88±0.18defgh |
| 11.石岩枫 Mallotus repandus | 10.62±3.24klmnopqr | 125.01±20.78jklm | 0.13±0.03ijkl | 83.82±6.47abcdefghij | 0.60±0.02ghijklmnop | 2.88±0.62defgh |
| 12.斜叶榕 Ficus tinctoriasubsp. gibbosa | 11.05±0.60mnopqr | 88.18±5.97defghijk | 0.11±0.01efghij | 98.77±4.81fghij | 0.50±0.02cdefgh | 3.65±0.21ghi |
| 13.密花树 Myrsine seguinii | 1.97±0.17a | 11.46±0.96a | 0.01±0.01a | 139.13±0.09mn | 0.45±0.01bcd | 0.23±0.02a |
| 14.紫凌木 Decaspermum esquirolii | 2.43±0.38ab | 25.70±4.47abc | 0.04±0.01abcd | 62.40±4.36ab | 0.69±0.02opqr | 1.17±0.17abc |
| 15.白马骨 Serissa serissoides | 4.95±0.49abcde | 180.46±21.03nop | 0.04±0.01abcd | 126.81±6.18klm | 0.48±0.03bcdef | 0.77±0.07ab |
| 16.麻叶绣线菊 Spiraea cantoniensis | 12.57±1.84opqr | 316.60±21.64r | 0.18±0.02lmno | 72.23±8.55abcdef | 0.62±0.04ijklmnopqr | 4.08±0.41ij |
| 17.竹叶花椒 Zanthoxylum armatum | 7.29±2.63defghijk | 80.02±17.29cdefghij | 0.05±0.02abcd | 143.22±11.61mno | 0.40±0.04ab | 1.23±0.31abc |
| 18.一叶萩 Flueggea suffruticosa | 9.23±0.52hijklmnop | 64.98±3.00abcdefghi | 0.13±0.01ijkl | 69.62±7.50abcd | 0.62±0.03jklmnopqr | 4.14±0.21ij |
| 19.钩齿鼠李 Rhamnus lamprophylla | 7.24±0.70defghijk | 107.02±15.02hijkl | 0.07±0.01bcdefg | 98.22±4.90efghij | 0.52±0.02defghi | 2.47±0.27d |
| 20.马甲子 Paliurus ramosissimus | 13.67±0.51r | 197.33±9.02opq | 0.22±0.03op | 63.40±4.82ab | 0.63±0.02jklmnopqr | 6.46±0.60m |
| 21.细梗女贞 Ligustrum tenuipes | 7.41±0.94defghijk | 49.87±6.49abcdefg | 0.07±0.01bcdefg | 100.99±10.01ghij | 0.59±0.04ghijklmnop | 1.26±0.07bc |
| 22.广西鼠李 Rhamnus kwangsiensis | 7.24±1.36defghijk | 36.60±6.45abcde | 0.09±0.01defghi | 82.97±4.88abcdefghij | 0.60±0.02ghijklmnopq | 2.96±0.38defgh |
| 23.檵木 Loropetalum chinense | 7.55±1.52efghijkl | 73.59±11.40bcdefghij | 0.12±0.02fghijk | 63.75±2.23ab | 0.69±0.01pqr | 2.57±0.42de |
| 24.薄叶鼠李 Rhamnus leptophylla | 12.66±1.24pqr | 370.13±50.73s | 0.21±0.03nop | 61.44±3.20a | 0.68±0.01nopqr | 4.30±0.49ij |
| 25.青冈栎 Cyclobalanopsis glauca | 3.29±0.83abc | 43.90±11.63abcdefg | 0.03±0.01ab | 130.76±10.75lm | 0.48±0.04bcde | 0.47±0.15ab |
| 26.化香树 Platycarya strobilacea | 7.04±1.34defghij | 141.76±24.9klmn | 0.08±0.02cdefgh | 93.00±6.77cdefghij | 0.61±0.02hijklmnopq | 1.20±0.29abc |
| 27.刺叶冬青 Ilex bioritsensis | 4.67±0.68abcde | 28.34±4.38abc | 0.05±0.01abcd | 108.73±11.15jkl | 0.55±0.04defghijkl | 0.82±0.18ab |
| 28.阴香 Cinnamomum burmannii | 10.89±0.12lmnopqr | 139.09±2.70klmn | 0.13±0.01ghijk | 86.59±0.62abcdefghij | 0.58±0.01fghijklmno | 2.72±0.02defg |
| 29.桂花 Osmanthus fragrans | 11.62±0.55mnopqr | 96.86±7.80ghijkl | 0.20±0.01no | 58.84±1.64a | 0.72±0.01r | 2.66±0.01def |
| 30.光皮梾木 Cornus wilsoniana | 5.53±0.13bcdef | 87.03±3.52defghijk | 0.07±0.01bcdefg | 74.47±4.39abcdefgh | 0.71±0.01qr | 1.18±0.04abc |
| 31.南岭柞木 Xylosma controversa | 6.50±0.81cdefghi | 94.29±48.12fghijkl | 0.09±0.01defghi | 73.08±4.32abcdef | 0.66±0.02mnopqr | 2.15±0.15cd |
| 32.菜豆树 Radermachera sinica | 5.05±0.62abcde | 91.99±12.33efghijkl | 0.03±0.01abc | 167.03±11.35o | 0.31±0.04a | 0.80±0.13ab |
| 33.紫弹树 Celtis biondii | 11.34±1.08mnopqr | 236.95±25.81q | 0.11±0.02efghij | 101.52±5.82hij | 0.50±0.02bcdefgh | 3.55±0.39fghi |
| 34.铜钱树 Paliurus hemsleyanus | 17.43±0.21s | 288.17±4.24r | 0.18±0.01mno | 95.96±3.32defghij | 0.52±0.01defghij | 3.91±0.12i |
| 35.扁片海桐 Pittosporum planilobum | 7.39±1.52defghijk | 34.71±7.35abcd | 0.08±0.02cdefgh | 97.16±8.32defghij | 0.52±0.03defghij | 2.55±0.61de |
| 36.齿叶黄皮 Clausena dunniana | 8.15±0.66efghijklm | 87.91±7.10defghijk | 0.09±0.01defghi | 94.63±7.48defghij | 0.59±0.02ghijklmnop | 1.36±0.20bc |
| 37.铁榄 Sinosideroxylon pedunculatum | 11.51±0.81mnopqr | 60.80±4.11abcdefghi | 0.19±0.02no | 59.68±3.63a | 0.68±0.01nopqr | 5.49±0.36kl |
| 38.柱果铁线莲 Clematis uncinata | 7.41±0.45defghijk | 41.51±3.84abcdefg | 0.11±0.01efghij | 66.36±1.37abc | 0.68±0.01nopqr | 2.51±0.12d |
| 39.山木通 Clematis finetiana | 12.54±0.38opqr | 94.53±2.73fghijkl | 0.20±0.01nop | 62.47±2.13ab | 0.64±0.01klmnopqr | 6.23±0.08lm |
| 40.青江藤 Celastrus hindsii | 5.40±0.83bcdef | 145.55±19.63lmn | 0.07±0.02bcdef | 89.30±19.94bcdefghij | 0.62±0.07ijklmnopqr | 0.98±0.26ab |
| 41.络石 Trachelospermum jasminoides | 6.46±1.02cdefghi | 92.01±10.34efghijkl | 0.05±0.01abcd | 137.60±11.66mn | 0.40±0.04abc | 1.31±0.29bc |
| 42.灰毛崖豆藤 Millettia cinerea | 3.48±0.85abc | 41.84±10.60abcdefg | 0.05±0.01abcd | 73.75±4.41abcdefg | 0.66±0.02mnopqr | 1.20±0.21abc |
| 43.无柄五层龙 Salacia sessiliflora | 2.31±0.57ab | 19.77±4.98ab | 0.03±0.01ab | 80.18±5.27abcdefghi | 0.67±0.02mnopqr | 0.47±0.09ab |
| 44.小果蔷薇 Rosa cymosa | 9.10±0.55ghijklmno | 113.15±7.97ijkl | 0.09±0.01defghi | 107.19±2.01ijkl | 0.53±0.01defghijk | 1.47±0.10bc |
| 45.吊山桃 Secamone sinica | 4.69±3.08abcde | 45.95±17.60abcdefg | 0.03±0.01abc | 130.67±21.29lm | 0.45±0.09bcd | 0.85±0.26ab |
| 46.疏叶崖豆 Millettia pulchravar. laxior | 10.35±0.82jklmnopqr | 87.13±6.67defghijk | 0.12±0.01fghij | 90.08±5.76bcdefghij | 0.56±0.03efghijklm | 3.72±0.25hi |
| 47.龙须藤 Bauhinia championii | 6.85±1.23fghijklmn | 112.30±8.41ijkl | 0.06±0.01abcd | 161.60±8.15no | 0.46±0.02bcd | 1.31±0.17ab |
| 48.皱叶雀梅藤 Sageretia rugosa | 7.08±0.70defghij | 64.53±5.35abcdefghi | 0.05±0.01abcd | 129.24±2.65lm | 0.50±0.05bcdefg | 1.41±0.13bc |
| 49.亮叶素馨 Jasminum seguinii | 5.71±1.04bcdefg | 39.02±6.87abcdef | 0.08±0.02cdefgh | 79.95±9.65abcdefghi | 0.60±0.03ghijklmnop | 2.64±0.74def |
| 50.皱果崖豆藤 Millettia oosperma | 12.27±0.86nopqr | 113.84±9.26ijkl | 0.13±0.03hijkl | 102.45±17.76ijk | 0.56±0.06efghijklm | 2.00±0.38cd |
| 方差分析 One-way ANOVA | F=12.85, P=0.00 | F=22.48, P=0.00 | F=15.95, P=0.00 | F=13.85, P=0.00 | F=10.17, P=0.00 | F=28.28, P=0.00 |
| 同一列不同字母表示差异显著(P < 0.05) | ||||||
对桂林喀斯特石山生境50种常见植物叶片的6个光合特性指标值分别进行Pearson相关性分析表明(表 2), Aarea与Gs, Amass和Tr之间分别均呈极显著正相关关系, 与WUE则相反, Aarea与Ci/Ca之间呈显著的正相关;Gs与Amass, Ci/Ca和Tr之间分别均呈极显著正相关, 与WUE则相反;Amass与Tr之间呈极显著的正相关, 与Ci/Ca之间呈显著的正相关, 与WUE则相反;Ci/Ca与Tr之间呈极显著的正相关, 与WUE则相反;Tr与WUE之间呈极显著的负相关。
| 叶片光合特性指标 Photosynthetic characteristics of leaves |
Aarea | Gs | Amass | Ci/Ca | Tr |
| Gs | 0.887** | ||||
| Amass | 0.676** | 0.592** | |||
| Ci/Ca | 0.169* | 0.480** | 0.049 | ||
| Tr | 0.785** | 0.899** | 0.442** | 0.373** | |
| WUE | -0.317** | -0.628** | -0.156 | -0.929** | -0.591** |
| **P<0.01, *:P<0.05. | |||||
对桂林喀斯特石山生境50种常见植物叶片的6个光合特性指标值进行主成分分析(表 3), 第一和第二主成分对总方差的贡献率分别为63.34%和24.55%, 两者累积贡献率达87.89%。对第一主成分而言, x3的系数最大为0.97, 表明Gs在第一主成分上具有较大的载荷。第一主成分在综合其他变量反映信息的基础上, 突出地反映了植物叶片Gs的信息。对第二主成分而言, x5的系数绝对值最大为0.74, 表明Ci/Ca比值在第二主成分上具有较大的载荷。第二主成分综合反映了植物叶片Ci/Ca的信息。由此可知, 表征桂林喀斯特石山50种常见植物之间叶片光合特性的主要指标为Gs, 其次为Ci/Ca。
| 主成分 Component |
特征向量 Eigenvector | 特征根 Eigenvalue |
贡献率 Cumulative/% |
|||||
| x1 (Aarea) | x2 (Amass) | x3 (Gs) | x4 (WUE) | x5 (Ci/Ca) | x6 (Tr) | |||
| y1 | 0.85 | 0.60 | 0.97 | -0.77 | 0.62 | 0.89 | 3.80 | 63.34 |
| y2 | 0.44 | 0.57 | 0.12 | 0.61 | -0.74 | 0.11 | 1.47 | 24.55 |
50种常见植物进行光合特性主成分排序(图 1), 3号样本(紫薇)、13号样本(密花树)、20号样本(马甲子)、24号样本(薄叶鼠李)、32号样本(菜豆树)和34号样本(铜钱树)是较为特殊的点, 其他样本(物种)较相似。铜钱树的第二主成分得分最高, 第一主成分得分也偏大, 表明铜钱树叶片Ci/Ca比值最大, 而Gs与其他物种差异偏大。薄叶鼠李的第一主成分得分最高, 第二主成分得分偏小, 表明薄叶鼠李叶片Gs高于其他植物, 而Ci/Ca比值与其他物种差异不大。紫薇和马甲子叶片的Gs仅次于薄叶鼠李。密花树叶片的Gs最小, Ci/Ca比值为均值, 菜豆树叶片的Gs仅次于密花树。

|
| 图 1 50种常见植物光合特性主成分排序图 Fig. 1 The figure of PCA ordination of photosynthetic characteristics of 50 common plants species 1:火棘Pyracantha fortuneana; 2:蔓胡颓子Elaeagnus glabra; 3:紫薇Lagerstroemia indica; 4:矮小天仙果Ficus erecta; 5:了哥王Wikstroemia indica; 6:小叶女贞Ligustrum quihoui; 7:小花扁担杆Grewia bilobavar. parviflora; 8:红背山麻杆Alchornea trewioides; 9:白皮乌口树Tarenna depauperata; 10:黄荆Vitex negundo; 11:石岩枫Mallotus repandus; 12:斜叶榕Ficus tinctoriasubsp. gibbosa; 13:密花树Myrsine seguinii; 14:紫凌木Decaspermum esquirolii; 15:白马骨Serissa serissoides; 16:麻叶绣线菊Spiraea cantoniensis; 17:竹叶花椒Zanthoxylum armatum; 18:一叶萩Flueggea suffruticosa; 19:钩齿鼠李Rhamnus lamprophylla; 20:马甲子Paliurus ramosissimus; 21:细梗女贞Ligustrum tenuipes; 22:广西鼠李Rhamnus kwangsiensis; 23:檵木Loropetalum chinense; 24:薄叶鼠李Rhamnus leptophylla; 25:青冈栎Cyclobalanopsis glauca; 26:化香树Platycarya strobilacea; 27:刺叶冬青Ilex bioritsensis; 28:阴香Cinnamomum burmannii; 29:桂花Osmanthus fragrans; 30:光皮梾木Cornus wilsoniana; 31:南岭柞木Xylosma controversa; 32:菜豆树Radermachera sinica; 33:紫弹树Celtis biondii; 34:铜钱树Paliurus hemsleyanus; 35:扁片海桐Pittosporum planilobum; 36:齿叶黄皮Clausena dunniana; 37:铁榄Sinosideroxylon pedunculatum; 38:柱果铁线莲Clematis uncinata; 39:山木通Clematis finetiana; 40:青江藤Celastrus hindsii; 41:络石Trachelospermum jasminoides; 42:灰毛崖豆藤Millettia cinerea; 43:无柄五层龙Salacia sessiliflora; 44:小果蔷薇Rosa cymosa; 45:吊山桃Secamone sinica; 46:疏叶崖豆Millettia pulchravar. laxior; 47:龙须藤Bauhinia championii; 48:皱叶雀梅藤Sageretia rugosa; 49:亮叶素馨Jasminum seguinii; 50:皱果崖豆藤Millettia oosperma |
基于Gs和Ci/Ca含量值, 通过聚类分析得出将50种常见植物划分为3类(图 2), 第一类(10种):火棘、铁榄、薄叶鼠李、桂花、马甲子、山木通、紫薇、小花扁担杆、麻叶绣线菊和一叶萩, Gs和Ci/Ca均值分别为0.19 mol m-2 s-1和0.65;第二类(29种):灰毛崖豆藤、紫凌木、檵木、柱果铁线莲、小叶女贞、光皮梾木、南岭柞木、了哥王、无柄五层龙、红背山麻杆、白皮乌口树、黄荆、石岩枫、阴香、疏叶崖豆、皱果崖豆藤、细梗女贞、广西鼠李、化香树、齿叶黄皮、青江藤、亮叶素馨、斜叶榕、紫弹树、扁片海桐、钩齿鼠李、刺叶冬青、小果蔷薇和铜钱树, Gs和Ci/Ca均值分别为0.09 mol m-2 s-1和0.60。第三类(11种):蔓胡颓子、龙须藤、密花树、白马骨、吊山桃、青冈栎、皱叶雀梅藤、竹叶花椒、络石、矮小天仙果和菜豆树, Gs和Ci/Ca均值分别为0.04 mol m-2 s-1和0.43。

|
| 图 2 50种常见植物的聚类分析 Fig. 2 Cluster analysis of 50 common plants species |
植物的光合特性往往可表征植物生长和抗逆性强弱[20], Gs是植物耐旱性的重要指标, 在一定程度上反映了同一生长环境下不同植物对环境的适应性, Gs的改变可对光合速率和蒸腾速率进行调节, 以适应外界环境[21]。Ci/Ca是反映植物光合速率大小的重要指标。由表 4可知, 本研究的Gs高于贵州[22]及退化喀斯特森林[23]常见植物, 低于广西典型园林[24]、桂西北[25]以及浑善达克沙地植物[26];Ci/Ca低于中科院研究所[27]及神农架林区[28]几种植物, 高于藏东南地区植物[29]。不同地区植物主要光合特性指标的差异, 可进一步体现植物对生境的适应性。Tr可在一定程度上反映植物调节水分和适应干旱环境的能力[30]。Aarea和Amass是植物生长、生理代谢过程和产量构成的主要因子。桂林喀斯特石山植物Gs, Ci/Ca, Aarea, Amass和Tr均值均低于贵州石漠化区29种优势植物种[31]和云南元江干热河谷24种木本植物[30], 由此可见, 相对于较高海拔的贵州喀斯特高原生态环境以及水分亏缺, 极度干热环境下较高海拔的云南元江干热河谷地区植物而言, 较低海拔的桂林喀斯特山地丘陵区植物光合碳同化能力较低, 净光合速率较小, 光合生产力较弱, 表现出较弱的光合特性以及耐旱性。同时在一定程度上反映植物生长缓慢, 具有保守生存策略[32], 表现出这些植物和较高海拔贵州喀斯特高原生态环境以及云南元江干热河谷地区植物采取的是不同的生长策略。较低海拔桂林喀斯特山地丘陵区植物具备一定的调节水分平衡的能力, 反映了植物对恶劣生境的适应表现出明显的响应特征, 整体上反映了植物对资源的利用能力。WUE是反映植物光合、蒸腾特性, 评价植物生长适应程度的综合指标, 反映植物的水分利用状况和抗旱性能[33]。对生境条件具有一定的响应, 随着生境条件的恶化, 植物的WUE将逐渐增加, 逐步调控植物对逆境的适应能力[34]。本研究中的WUE均值远高于贵州石漠化区29种优势植物种[31]和云南元江干热河谷24种木本植物[30], 这表明相对于较高海拔的贵州喀斯特高原生态环境以及水分亏缺, 极度干热环境下较高海拔的云南元江干热河谷地区植物而言, 较低海拔桂林喀斯特山地丘陵区植物具有较高的水分利用效率, 同时具备适应桂林喀斯特石山干旱的特征或避旱策略, 该地区植物采取保守的水分利用策略, 通过“节流”策略来适应干旱胁迫。
| 项目 Item |
气孔导度Gs Stomatal conductance/(mol m-2 s-1) |
Ci/Ca Intercellular/environmental CO2 concentration |
文献来源 Literature sources |
| 贵州喀斯特7种常见树种 7 common tree species in Guizhou karst |
0.08 | — | [22] |
| 退化喀斯特森林8种常见植物 8 common plants in degraded karst forest |
0.07 | — | [23] |
| 广西15种典型园林植物 15 typical garden plants in Guangxi |
0.34 | 0.21 | [24] |
| 桂西北喀斯特地区典型植物 Typical plants in karst area of northwest Guangxi |
0.12 | — | [25] |
| 浑善达克沙地14种植物 14 species of plants in the Ortindag sandy |
0.21 | — | [26] |
| 中科院研究所5种植物 5 species of plants from the institute of Chinese Academy of Sciences |
— | 0.62 | [27] |
| 神农架林区8种植物 8 plants in Shennongjia national nature reserve |
— | 0.70 | [28] |
| 藏东南15个物种 15 species in southeastern Tibet |
0.1 | 0.24 | [29] |
| 元江干热河谷24种木本植物 24 woody plants in a dry-hot valley of Yuanjiang |
0.23 | 0.66 | [30] |
| 本研究(桂林喀斯特石山常见植物) This study (common plants on karst rock mountain in Guilin) |
0.10 | 0.57 |
总体而言, 桂林喀斯特石山植物在Aarea, Amass, Gs, WUE, Ci/Ca和Tr之间存在明显差异, 说明这些物种虽然生长在非常相似的环境中, 但在光合策略上有着明显差异, 以不同的方式适应相似的生境条件, 体现出植物为了最大限度获取碳而采取的生存适应策略[35]。叶片光合生理指标在不同植物类群间的不同显示了不同物种在资源利用上可能的差异, 而这种差异将有助于植物对环境资源的充分利用, 进而提高整个系统的稳定性[36]。
3.2 叶片光合特性之间的相关性在桂林喀斯特石山生态环境恢复过程中, 植物会不断调整资源配置及生理过程, 以适应生境的变化, 最终在光合特性方面作出响应[17]。本研究表明, 表征50种常见植物叶片光合特性的6个指标相关性除Ci/Ca与Aarea和Amass, WUE与Amass不一致外, 其他指标两两之间相关性均表现为一致性。桂林喀斯特石山植物Tr与WUE之间呈极显著负相关, 这与贵州石漠化区29种优势植物种[31]研究结果相似, 说明石漠化地区植物WUE伴随着蒸腾耗水量的降低而提高, 植物通过“节流”策略来适应干旱胁迫。Tr与Gs呈极显著正相关, 这与金沙江干热河谷29种植被恢复树种[37]的研究结果相似。Aarea和Gs之间呈现极显著正相关, 此结果与云南元江干热河谷24种木本植物[30]研究结果一致。王春雪等[38]对元江干热河谷酸角的光合作用研究中也表明Gs是影响光合作用的初始因素, 气孔的开闭直接影响Tr等, 即影响酸角净光合速率的主要是气孔因素, 气孔的关闭导致了光合参数的降低。
3.3 基于Gs和Ci/Ca的植物功能群特征本研究得出表征桂林喀斯特石山50种常见植物叶片光合特性的主要指标为Gs和Ci/Ca, 这与刘泽[39]和付威波[40]等研究结果一致。相对于其他地区植物而言, 本研究中的第一类植物, Gs和Ci/Ca均值分别为0.19 mol m-2 s-1和0.65, 为中等Gs较高Ci/Ca类型, 这表明火棘、铁榄、薄叶鼠李、桂花、马甲子、山木通、紫薇、小花扁担杆、麻叶绣线菊和一叶萩这10个物种在耐旱性方面表现较强, 以应对干旱胁迫, 有利于对生长环境干湿程度的适应和固碳释氧能力提升, 促进植物的光合作用、呼吸作用及蒸腾作用, 对于衡量植物与环境间的水分、CO2平衡以及循环具有重要作用。基于以上光合特性, 第一类植物适应策略突出表现在较好抗旱特性及光合能力, 以抵御外界不利环境, 对喀斯特生境具有较强的适应能力, 为此可考虑第一类植物作为植被恢复重建的首选树种;第二类植物Gs和Ci/Ca均值分别为0.09 mol m-2 s-1和0.60, 为较低Gs较高Ci/Ca类型, 这表明灰毛崖豆藤、紫凌木、檵木、柱果铁线莲、小叶女贞、光皮梾木、南岭柞木、了哥王、无柄五层龙、红背山麻杆、白皮乌口树、黄荆、石岩枫、阴香、疏叶崖豆、皱果崖豆藤、细梗女贞、广西鼠李、化香树、齿叶黄皮、青江藤、亮叶素馨、斜叶榕、紫弹树、扁片海桐、钩齿鼠李、刺叶冬青、小果蔷薇和铜钱树这29个物种整体上表现为具有一定耐旱性, 并伴随较高的光合速率, 有利于光合产物的积累, 一定程度上可较好与喀斯特生境相适应, 应对干旱胁迫, 第二类植物以高光合能力的适应策略, 参与喀斯特生境植被恢复与重建过程中的树种选择;第三类植物Gs和Ci/Ca均值分别为0.04 mol m-2 s-1和0.43, 为较低Gs, Ci/Ca类型, 这表明蔓胡颓子、龙须藤、密花树、白马骨、吊山桃、青冈栎、皱叶雀梅藤、竹叶花椒、络石、矮小天仙果和菜豆树这11个物种, 具有相对较低的光合速率, 同时具有低蒸腾高水分利用的特点, 对于植被恢复重建而言, 第三类植物在水分利用方面表现出独特优势, 相比其他物种更能有效地节约利用土壤水分, 即消耗等量水分可生产更多的干物质, 较好适应喀斯特地区的干旱环境[41]。在植被恢复与重建过程中, 三类植物在不同的光合指标方面展现出不同的优势, 最终表现为耐受高温及干旱胁迫, 有利于植被群落朝着稳定性及抗逆性强的方向发展, 符合可持续的生态恢复理念[42]。今后对于物种选择不仅要重视植被恢复树种的生理生态机制, 同时需结合外部生态环境的变化进行整体评估。
综上所述, 本研究分析了50种常见植物叶片光合特性之间的差异及其内在联系, 结果表明Gs和Ci/Ca可作为反映桂林喀斯特石山植物适应生境的重要光合指标, 主要表征植物耐旱性及光合特性强弱, 用于衡量植物对喀斯特生境的适应特性。由本研究可知, 喀斯特生境植物在生理生态方面所表现出的适应策略主要为对资源利用方式及抵御外界不利环境的适应策略, 这为后续选择物种加速植被恢复演替进程提供了参考。
| [1] |
郭雯, 徐瑞晶, 漆良华, 胡璇, 丁霞, 程昌锦, 张建, 雷刚. 竹类植物光合特性与叶片功能性状研究. 世界林业研究, 2018, 31(4): 29-35. |
| [2] |
Pierangelini M, Stojkovic S, Orr P T, Beardall J. Photosynthetic characteristics of two Cylindrospermopsis raciborskii strains differing in their toxicity. Journal of Phycology, 2014, 50(2): 292-302. |
| [3] |
付忠, 谢世清, 徐文果, 岩所, 陈军文. 不同光照强度下谢君魔芋的光合作用及能量分配特征. 应用生态学报, 2016, 27(4): 1177-1188. |
| [4] |
宋贺, 于鸿莹, 陈莹婷, 许振柱, 周广胜. 北京植物园不同功能型植物叶经济谱. 应用生态学报, 2016, 27(6): 1861-1869. |
| [5] |
陈振, 元慕田, 曹琪琪, 刘涛, 肖辉杰. 土壤含水量对苜蓿和沙棘气孔导度与叶水势的影响. 中国水土保持科学, 2019, 17(2): 37-43. |
| [6] |
林艳华, 梁千慧, 刘锦春. 喀斯特地区适生树种复羽叶栾树幼苗对干旱胁迫下异质生境的生长和光合响应. 西南大学学报:自然科学版, 2019, 41(8): 20-26. |
| [7] |
唐俊煜, 冉丽, 张琦. 不同品种桃叶片光合特性日变化的研究. 北方果树, 2019(3): 5-9. |
| [8] |
郎校安, 杨蕾蕾, 崔铁成, 张寿洲, 戚建华. 红花深山含笑光合速率日变化及其与环境因子的关系. 西南林业大学学报, 2017, 37(4): 22-27. |
| [9] |
何丽娜, 王德炉, 郝家孝, 谢双喜. 4个兔眼蓝莓品种光合特性对季节变化的响应. 经济林研究, 2019, 37(2): 95-103. |
| [10] |
屠凯, 温国胜, 侯平. 红叶石楠绿叶与红叶的光合蒸腾特性比较研究. 中国农学通报, 2019, 35(15): 110-115. |
| [11] |
Niinemets Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology, 2001, 82(2): 453-469. |
| [12] |
郭连金, 杜佳朋, 吴艳萍, 肖志鹏, 廖婷, 邵秀娟. 香果树实生苗的光合特性及其与环境因子的关系. 应用生态学报, 2017, 28(5): 1473-1481. |
| [13] |
贾双杰, 李红伟, 江艳平, 赵国强, 王和洲, 杨慎骄, 杨青华, 郭家萌, 邵瑞鑫. 干旱胁迫对玉米叶片光合特性和穗发育特征的影响. 生态学报, 2020, 40(3): 854-863. |
| [14] |
唐星林, 姜姜, 金洪平, 周晨, 刘光正, 杨桦. 遮阴对闽楠叶绿素含量和光合特性的影响. 应用生态学报, 2019, 30(9): 2941-2948. |
| [15] |
郑鹏丽, 黄晓蓉, 费永俊, 周明芹. 水分胁迫对桢楠幼树光合生理特性的影响. 中南林业科技大学学报, 2019, 39(10): 64-70. |
| [16] |
孙胜楠, 王强, 孙晨晨, 刘丰娇, 毕焕改, 艾希珍. 黄瓜幼苗光合作用对高温胁迫的响应与适应. 应用生态学报, 2017, 28(5): 1603-1610. |
| [17] |
莫燕华, 马姜明, 苏静, 秦佳双, 潘小梅, 菅瑞. 桂林岩溶石山檵木群落老龄林植物叶性状. 广西植物, 2019, 39(8): 1059-1068. |
| [18] |
马姜明, 占婷婷, 莫祖英, 梁士楚. 漓江流域岩溶区檵木群落不同恢复阶段主要共有种生态位变化. 西北植物学报, 2012, 32(12): 2530-2536. |
| [19] |
马姜明, 黄婧, 杨栋林, 梅军林. 桂林喀斯特石山50种常见植物叶片光合色素含量及耐荫性定量评价. 林业科学, 2015, 51(10): 67-74. |
| [20] |
Kumari S, Agrawal M. Growth, yield and quality attributes of a tropical potato variety (Solanum tuberosum L. cv Kufri chandramukhi) under ambient and elevated carbon dioxide and ozone and their interactions. Ecotoxicology and Environmental Safety, 2014, 101: 146-156. |
| [21] |
Fischer R A, Turner N C. Plant productivity in the arid and semiarid zones. Annual Review of Plant Physiology, 1978, 29: 227-317. |
| [22] |
姜霞, 张喜. 贵州喀斯特山地几种常见树种的光合特性比较. 林业科技开发, 2011, 25(3): 17-22. |
| [23] |
丁晓雅, 罗绪强, 冯双, 任艳, 莫愁, 陆汉芝, 李艳琴, 秦会斌. 退化喀斯特森林不同植物叶片气孔导度对环境因子的响应. 山东农业科学, 2020, 52(4): 32-37. |
| [24] |
高传友. 广西地区15种典型园林观赏植物的耐阴性及光合特性. 江苏农业科学, 2016, 44(4): 209-214. |
| [25] |
邓彭艳.桂西北喀斯特地区典型植物光合及水分生理特性研究[D].桂林: 广西师范大学, 2010.
|
| [26] |
李清雪.浑善达克沙地植物光合特性及蒸腾特性研究[D].呼和浩特: 内蒙古农业大学, 2011.
|
| [27] |
程建峰, 陈根云, 沈允钢. 植物叶片特征与光合性能的关系. 中国生态农业学报, 2012, 20(4): 466-473. |
| [28] |
程建峰, 陈根云, 沈允钢. 神农架林区不同类型植物的叶片特征与光合性能研究. 生态环境学报, 2010, 19(1): 165-171. |
| [29] |
关志华, 王太航, 王陆州, 赵唯唯, 陈双臣, 王忠红. 藏东南15个辣椒地方品种开花结果期的光合——光响应特性. 高原农业, 2018, 2(2): 118-125, 157-157. |
| [30] |
张教林.元江干热河谷木本植物的物候、叶片结构和功能关系及光合作用的季节变化[D].西双版纳: 中国科学院研究生院(西双版纳热带植物园), 2007.
|
| [31] |
张仕豪, 熊康宁, 张俞, 季传泽, 马学威, 赖佳丽, 闵小莹. 典型石漠化生态系统演替过程优势植物种叶片功能性状特征及影响因素. 生态环境学报, 2019, 28(11): 2165-2175. |
| [32] |
张晶, 赵成章, 李雪萍, 任悦, 雷蕾. 嘉峪关草湖湿地芦苇净光合速率与叶面积和叶厚度的关系. 生态学报, 2018, 38(17): 6084-6091. |
| [33] |
刘娟, 杜曲, 涂淑萍. 圆齿野鸦椿光合特性日变化的比较. 植物学研究, 2019, 8(5): 396-402. |
| [34] |
张瑞文, 赵成义, 王丹丹, 施枫芝, 郑金强. 极端干旱区不同水分条件下胡杨林生态耗水特征. 水土保持学报, 2019, 33(4): 270-278. |
| [35] |
Cordell S, Goldstein G, Meinzer F C, Vitousek P M. Regulation of leaf life-span and nutrient-use efficiency of Metrosideros polymorpha trees at two extremes of a long chronosequence in Hawaii. Oecologia, 2001, 127(2): 198-206. |
| [36] |
Anderson J E, Inouye R S. Landscape-scale changes in plant species abundance and biodiversity of a sagebrush steppe over 45 years. Ecological Monographs, 2001, 71(4): 531-556. |
| [37] |
段爱国.干热河谷主要植被恢复树种蒸腾耗水特性及适应机制评价[D].北京: 中国林业科学研究院, 2008.
|
| [38] |
王春雪, 何光熊, 宋子波, 樊博, 张梦寅, 方海东, 王艳丹, 史亮涛. 元江元谋干热河谷土壤氮磷水平对酸角叶片氮磷含量及光合的影响. 生态学杂志, 2019, 38(3): 710-718. |
| [39] |
刘泽, 郑建伟, 潘景玉, 裴姿琛, 兰济艳, 李保会. 不同杜仲单株光合特性的比较. 中国科技论文, 2019, 14(4): 391-398. |
| [40] |
付威波, 宋敏, 彭晚霞, 杨钙仁, 杜虎, 文丽, 李莎莎. 西南喀斯特典型树种光合特性的季节变化及主要影响因子. 农业现代化研究, 2015, 36(5): 890-894. |
| [41] |
魏媛, 胡刚, 杨萍, 周家春. 喀斯特山地不同景观树种的光合特性及生态适应性. 贵州农业科学, 2013, 41(1): 65-68. |
| [42] |
沈利娜, 蒋忠诚, 覃星铭, 吴孔运, 梁铭忠, 马祖陆, 李晖. 峰丛洼地恢复树种光合生理生态特征日变化研究——以广西马山弄拉峰丛洼地为例. 广西师范大学学报:自然科学版, 2010, 28(1): 47-53. |