 2020, Vol. 40
2020, Vol. 40文章信息
- 刘若莎, 王冬梅, 李平, 靳永庆, 梁士楚
- LIU Ruosha, WANG Dongmei, LI Ping, JIN Yongqing, LIANG Shichu
- 青海高寒区典型人工林植物多样性、地上生物量特征及其相关性
- Plant diversity, ground biomass characteristics and their relationships of typical plantations in the alpine region of Qinghai
- 生态学报. 2020, 40(2): 692-700
- Acta Ecologica Sinica. 2020, 40(2): 692-700
- http://dx.doi.org/10.5846/stxb201901160133
-
文章历史
- 收稿日期: 2019-01-16
- 网络出版日期: 2019-01-04





2. 大通县林业站, 西宁 810100;
3. 广西师范大学生命科学学院, 桂林 541006
2. Forest Station of Datong County, Xining 810100, China;
3. College of Life Sciences, Guangxi Normal University, Guilin 541006, China
植物多样性反映了群落的稳定程度和结构类型, 其对生态系统功能的影响一直是生态学领域研究的热点[1-2]。生物量指区域范围内所有物种的总干重, 是生态系统功能的重要表现形式[3-4]。因此, 研究植物多样性与生物量的关系有助于阐明植物多样性在生态系统功能中的作用, 还可以对植物多样性及生物量进行相互预测, 并为当地生态系统的结构调整及可持续发展提供科学依据[5-6]。
在自然群落中影响植物多样性与生物量关系的有生物、环境等诸多因素, 这往往会导致相应的生态学过程有所不同, 两者的关系也会表现出多种形式[7-8]。目前许多研究表明, 两者关系的表现形式有单峰关系、正线性关系、负线性关系和不相关[9-10]。近年来随着研究的不断深入, 对高寒区植物多样性与生物量的关系也进行了许多研究[11]。由于高寒区具有海拔较高, 气温较低, 空气稀薄, 土层浅薄等特征, 植被多以草甸形式存在, 因此研究多集中在高寒草地、高寒灌丛等方向。例如朱桂丽等[11]对西藏高寒草地进行了研究, 得出高寒草地植物丰富度与地上生物量间存在显著的幂指数关系。武彦朋等[9]研究了在群落梯度上高寒湿地植物多样性与生产力的关系, 结果表明无论是在群落尺度上还是在区域尺度上植物多样性与地上生物量呈显著负相关。牛钰杰等[5]探讨了放牧干扰下高寒草甸植物多样性与生物量的关系, 得出了在低放牧强度下植物丰富度与生物量呈显著负相关。杨路存等[1]对青海省高寒灌丛的植物多样性与生物量关系进行了调查研究, 得出草本生物量与植物多样性呈负相关, 而灌木植物多样性与其生物量无显著相关性。在青藏高原的高寒草原生态系统中人工林生态系统是植被恢复的主要方式[12]。但目前对高寒区人工林生态系统的植物多样性与生物量关系研究较少, 在高寒区特殊条件下, 人工林植物多样性与生物量是否还呈现明显显著关系尚不清楚。
青海高寒区的人工林树种主要是青海云杉、白桦、华北落叶松、青杨、祁连圆柏等[13-15], 因此, 本文选用青海云杉、白桦、华北落叶松和青杨人工林作为高寒区人工林的典型代表进行研究。通过对人工林植被群落的植物多样性、地上生物量特征及其相互关系进行调查分析, 以期阐明青海高寒区人工林群落植物多样性与生物量的关系。其结果不仅为高寒区人工林植物多样性与生物量的研究提供参考依据, 还对人工林恢复与重建具有重要意义。
1 研究区概况研究区位于青海省东北部大通县的闇门滩、塔尔沟两个小流域, 地理坐标分别为36°54′—36°56′N, 101°40′—101°41′E;37°1′—37°2′N, 101°40′—101°42′E;海拔分别在2460—2650 m, 2840—2970 m之间;塔尔沟位于闇门滩正北方向, 两地相距12500 m, 均属半干旱半湿润的大陆性气候。由于海拔的不同, 闇门滩与塔尔沟年均气温分别为2.8℃、2.4℃, 年内无霜期分别在70—130天, 60—120天之间。两流域土壤类型主要为山地棕褐土, 年降水量在450—820 mm之间[16]。通过对当地护林员进行采访得知, 塔尔沟人工林于1984年实施退耕还林, 到目前为止人工林有35年的林龄, 显著影响土壤发育, 形成的土壤较肥沃;闇门滩于2000年进行退耕还林, 到目前为止人工林有20年的林龄, 对土壤发育影响稍弱。通过野外调查得到塔尔沟流域人工林均种植在阴坡坡向, 而闇门滩流域人工林在阴、阳坡均有种植。本文选择两流域阴坡坡向的人工林进行研究。两流域的植被类型基本无差异主要是寒温性常绿针叶林及落叶阔叶林类, 主要树种有青海云杉(Picea crassifolia)、华北落叶松(Larix principis-rupprechtii)、青杨(Populus cathayana)、白桦(Betula platyphylla)等, 主要灌木种有沙棘(Hippophae rhamnoides)、柠条锦鸡儿(Caragana korshinskii)、枸杞(Lycium chinense)、黄芦木(Berberis amurensis)高山绣线菊(Spiraea alpina)、金露梅(Potentilla fruticosa)等, 主要草本物种有紫花苜蓿(Medicago sativa)、垂穗披碱草(Elymus nutans)、问荆(Equisetum arvense)、珠牙蓼(Polygonum viviparum)等[17]。
2 研究方法 2.1 样地设置与植被调查本研究于2018年7至8月, 在青海省大通县闇门滩、塔尔沟两个小流域进行, 共选取12种典型人工林, 27个标准地, 对乔木、灌木、草本进行分层调查;闇门滩流域因无混交林, 选择4种纯林共11个标准地进行研究;塔尔沟流域选择4种混交林与4种纯林共16个标准地进行研究。标准地面积设置为20 m×20 m, 运用生长锥、GPS定位等, 测定标准地的海拔、坡度、坡向、坡位、林龄等基本信息, 并对乔木的种类、高度、胸径、冠幅、数量等进行调查与记录;在标准地对角线方向上均匀设置3个5 m×5 m的灌木样方, 共计81个样方, 对灌木种类、高度、数量、地径、盖度等进行调查并记录;在标准地内按“米”字形均匀设置9个1 m×1 m的草本样方, 共计243个样方, 对草本种类、高度、数量和盖度等进行调查并记录(表 1)。
| 分区 Partition |
林分代号 Code |
林分类型 Forest types |
标准地数量/个 Number of standard plots |
海拔/m Altitude | 坡度/(°) Slope degree |
坡位 Slope position |
密度/(株/m2) Density |
| 闇门滩 | A1 | 华北落叶松纯林 Larix principis-rupprechtii |
3 | 2517 | 23° | 中下坡 | 0.2020 |
| A2 | 青杨纯林Populus cathayana | 3 | 2558 | 32° | 中上坡 | 0.1380 | |
| A3 | 白桦纯林Betula platyphylla | 2 | 2557 | 7° | 中上坡 | 0.119 | |
| A4 | 青海云杉纯林 Picea crassifolia |
3 | 2523 | 12° | 中坡 | 0.1590 | |
| 塔尔沟 | T1 | 华北落叶松纯林 Larix principis-rupprechtii |
2 | 2875 | 14° | 中坡 | 0.1700 |
| T2 | 青杨纯林Populus cathayana | 2 | 2906 | 23° | 中坡 | 0.1630 | |
| T3 | 白桦纯林Betula platyphylla | 2 | 2944 | 18° | 中上坡 | 0.1680 | |
| T4 | 青海云杉纯林 Picea crassifolia |
2 | 2954 | 20° | 中上坡 | 0.3130 | |
| T5 | 青海云杉-华北落叶松混交林 Picea crassifolia-Larix principis-rupprechtii |
2 | 2917 | 20° | 中坡 | 0.3050 | |
| T6 | 白桦-青杨混交林 Betula platyphylla-Populus cathayana |
2 | 2869 | 17° | 中下坡 | 0.1630 | |
| T7 | 青海云杉-青杨混交林 Picea crassifolia-Populus cathayana |
2 | 2909 | 13° | 中坡 | 0.240 | |
| T8 | 白桦-青海云杉混交林 Betula platyphylla-Picea crassifolia |
2 | 2948 | 20° | 中上坡 | 0.2810 |
通过样地调查, 根据如下公式分别计算不同人工林植被群落的丰富度指数、多样性指数、Simpson指数和均匀度指数[18]。
丰富度指数:
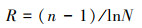
|
Shannon-Wiener指数:

|
Simpson指数:
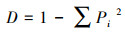
|
Pielou均匀度指数:
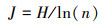
|
式中, 计算乔木、灌木、草本层多样性指数时, n为物种总数, N为所有物种个体数之和, Pi为种i的物种个体数占群落个体数的比例, 即Pi=ni/N, ni为种i个体数;计算群落多样性指数时, Pi为种i的相对重要值, 即Pi=Ni/N, Ni为第i个物种的重要值, N为所有重要值之和, 重要值IV=(相对频度+相对盖度+相对高度)/3[3]。
2.3 地上生物量数据获取乔木层地上部分生物量通过样方调查测定树高、胸径, 根据《中国立木材积表》[19]对样地树木进行立木材积估算, 通过蓄积-生物量模型[20]计算单位面积乔木层地上部分的生物量。
灌木层和草本层的地上部分生物量通过收获法获取, 灌木层的收获样方设在每个5 m×5 m灌木样方的中间, 草本层的收获样方选择标准地内呈对角线方向的3个植被调查样方。在设置的收获样方内对灌、草层地上部分进行收割, 称重, 测定灌、草层地上部分鲜重;同时将称过鲜重的灌、草地上部分取样带回实验室(各取样100 g左右), 用烘干法在85℃条件下烘干至恒重, 根据样品的干鲜重比值计算不同样方内灌木和草本的地上生物量;通过3个样方生物量的平均值, 计算单位面积灌木层、草本层地上生物量[21]。
2.4 数据处理与统计分析利用SPSS 24.0进行数据统计分析, 单因素方差分析比较不同人工林群落的差异显著性;采用SPSS 24.0软件对高寒区人工林植物多样性与生物量进行相关性分析及回归分析, 选择其中较为简单并且解释度高的曲线进行拟合作为最终结果;利用Microsoft Excel 2010软件对数据进行处理分析和作图。
3 结果与分析 3.1 不同人工林植物多样性从青海高寒区典型人工林的植物多样性指数可以看出(见表 2), 在坡向、坡位、林龄等因素基本相同的情况下, 林龄为20年的人工林群落中丰富度指数(R)、Shannon-Wiener指数(H)最高的为人工林A1, 最低的为人工林A4, 总体趋势表现为人工林A1>人工林A2>人工林A3>人工林A4, 且人工林A1、A2与人工林A3、A4差异显著;Simpson指数(D)和Pielou均匀度指数(J)均以人工林A1最高, 人工林A3最低。灌木层的丰富度指数(R)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J)最高的均为人工林A2, 其值分别为0.80、0.92、0.55、0.87;人工林A1、A3和A4, 均无值, 说明在人工林A2中灌木物种较丰富。草本层的丰富度指数(R)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J)均以人工林A1最高, 其值分别为4.07、2.63、0.89、0.80, 以人工林A3最低, 其值分别为1.36、0.89、0.25、0.32, 总体趋势表现为人工林A1>人工林A2>人工林A4>人工林A3, 且人工林A1、A2与人工林A3、A4差异显著。通过表 2分析得出林龄为20年的人工林群落与草本层植物多样性表现趋势大致相同, 均以人工林A1最高;灌木层与草本层植物多样性指数对比分析得出人工林下的草本物种较灌木丰富。
| 林分代码 Code |
群落植物多样性指数 Community plant diversity index |
灌木植物多样性指数 Shrub plant diversity index |
草本植物多样性指数 Herb plant diversity index |
|||||||||||
| R | H | D | J | R | H | D | J | R | H | D | J | |||
| A1 | 27.00±5.86a | 2.89±0.15a | 0.93±0.01a | 0.88±0.01a | — | — | — | — | 4.07±0.64a | 2.63±0.10a | 0.89±0.02a | 0.80±0.03a | ||
| A2 | 26.00±3.00a | 2.48±0.17a | 0.87±0.02ab | 0.76±0.03b | 0.80±0.36 | 0.92±0.30 | 0.55±0.13 | 0.87±0.04 | 3.39±0.52a | 2.10±0.19a | 0.80±0.05ab | 0.67±0.05ab | ||
| A3 | 15.00±2.12b | 1.97±0.18b | 0.78±0.04b | 0.74±0.03b | — | — | — | — | 1.36±0.28b | 0.89±047b | 0.25±0.21c | 0.32±0.16c | ||
| A4 | 13.00±1.53b | 1.96±0.31b | 0.79±0.07b | 0.76±0.09b | — | — | — | — | 1.44±0.26b | 1.15±0.66b | 0.51±0.25bc | 0.44±0.23bc | ||
| T1 | 14.00±8.49bc | 1.88±0.67ab | 0.77±0.14bc | 0.73±0.08ab | — | — | — | — | 1.68±1.21a | 1.46±0.66ab | 0.64±0.22a | 0.51±0.09ab | ||
| T2 | 23.00±2.12ab | 2.33±0.10a | 0.84±0.04ab | 0.75±0.06ab | — | — | — | — | 1.47±0.18a | 1.28±0.26bc | 0.52±0.13a | 0.41±0.10bc | ||
| T3 | 10.00±4.24bc | 1.53±0.14b | 0.67±0.02b | 0.69±0.07b | — | — | — | — | 1.02±0.49a | 0.32±0.21c | 0.13±0.11c | 0.16±0.12d | ||
| T4 | 6.00±0.71c | 1.44±0.07b | 0.73±0.01b | 0.74±0.02ab | — | — | — | — | 1.36±0.25a | 1.20±0.08bc | 0.39±0.01ab | 0.31±0.01cd | ||
| T5 | 14.00±2.12bc | 2.04±0.05ab | 0.83±0.01ab | 0.79±0.03a | — | — | — | — | 1.72±0.23a | 1.77±0.25a | 0.77±0.07a | 0.73±0.05a | ||
| T6 | 30.00±2.83a | 2.54±0.08a | 0.89±0.01a | 0.75±0.04ab | 1.08±0.45 | 1.07±0.33 | 0.60±0.08 | 0.76±0.06 | 1.70±1.29a | 1.48±0.25ab | 0.67±0.01a | 0.61±0.16a | ||
| T7 | 23.00±10.02ab | 2.12±0.23ab | 0.83±0.02ab | 0.70±0.05b | — | — | — | — | 1.69±1.44a | 1.47±0.23ab | 0.65±0.30a | 0.58±0.23ab | ||
| T8 | 14.00±4.95bc | 1.82±0.35ab | 0.78±0.06bc | 0.71±0.03b | — | — | — | — | 1.30±0.59a | 0.63±0.66b | 0.27±0.02bc | 0.24±0.03cd | ||
| 同列不同小写字母表示不同群落间的差异显著(P<0.05); R, 丰富度指数Richness index;H, Shannon-Wiener指数Shannon-Wiener index; D, Simpson指数Simpson index; J, Pielou均匀度指数Pielou evenness index | ||||||||||||||
林龄为35年的人工林群落中丰富度指数(R)、Shannon-Wiener指数(H)最高的为人工林T6, 其值分别为30.00、2.54, 最低的为人工林T4, 其值分别为6.00、1.44, 人工林T6与其他人工林差异显著;Simpson指数(D)和Pielou均匀度指数(J)最高的分别为人工林T6、T5, 最低的均为人工林T3。灌木层的丰富度指数(R)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J)最高的均为人工林T6, 其余7种人工林均无值, 说明在人工林T6中灌木物种较丰富。草本层的丰富度指数(R)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J)均表现为人工林T5最高, 其值分别为1.72、1.77、0.77、0.73, 人工林T3最低, 其值分别为1.02、0.32、0.13、0.16, 总体趋势为人工林T5>人工林T6>人工林T7>人工林T1>人工林T2>人工林T4>人工林T8>人工林T3, 除草本层丰富度指数(R)差异不显著外, 其余三个多样性指标均差异显著, 表明人工林T5的草本层物种最丰富。通过表(2)中灌木层与草本层植物多样性指数的对比分析得出在林龄为35年的人工林群落中草本物种较灌木丰富, 这与20年林龄的人工林得出的结果一致。
3.2 不同人工林地上生物量根据图 1可知在林龄为20年的人工林群落中, 地上部分总生物量最高的为人工林A2, 其值为6314.9671 g/m2, 最低的为人工林A4, 其值为921.9532 g/m2, 且差异显著, 从侧面反映出在20年林龄的人工林群落中人工林A2发挥生态功能的潜能较高。林龄为35年的人工林群落中, 地上总生物量以人工林T5最高, 其值为15181.1201 g/m2, 人工林T8最低, 其值为9563.0731 g/m2, 且差异显著, 由此可看出35年林龄的人工林群落中以人工林T5累积生物量最高, 从侧面反映出其发挥生态功能的潜能较高。

|
| 图 1 不同人工林群落及乔木地上生物量特征 Fig. 1 Characteristics of above-ground biomass in different plantation communities and trees 同列不同小写字母表示不同群落间的差异显著(P<0.05) |
根据图 1可知20年林龄的人工林A1、A2、A3、A4乔木地上部分生物量均比同种林分类型下35年林龄的人工林T1、T2、T3、T4乔木地上生物量低, 说明随着林龄的增长, 乔木地上部分生物量不断增加。图 1中显示不同人工林群落与乔木地上生物量差异较小, 表 3中灌木层与草本层地上部分生物量占地上总生物量的比例较小, 分别在0.77%—1.49%, 0.004%—18.54%之间, 表明群落地上部分总生物量主要取决于乔木层地上生物量。
| 林分代号 Code |
单位面积灌木地上生物量/(g/m2) Shrub biomass per unit area above ground |
单位面积草本地上生物量/(g/m2) Herb biomass per unit area above ground |
灌木生物量占总生物量比值/% Shrub biomass as a percentage of total biomass |
草本生物量占总生物量比值/% Herb biomass as a percentage of total biomass |
| A1 | — | 24.07±20.40c | — | 0.5167 |
| A2 | 93.83±57.66 | 54.75±26.97c | 1.4859 | 0.8669 |
| A3 | — | 234.07±20.62a | — | 15.8819 |
| A4 | — | 170.89±40.35b | — | 18.5357 |
| T1 | — | 25.09±10.97ab | — | 0.2192 |
| T2 | 101.09±32.98 | 83.08±46.83a | 0.7822 | 0.6427 |
| T3 | — | 28.92±13.88ab | — | 0.2796 |
| T4 | — | 0.51±0.23b | — | 0.0041 |
| T5 | — | 13.31±2.41b | — | 0.0877 |
| T6 | 82.16±48.02 | 80.60±37.12a | 0.7776 | 0.7628 |
| T7 | — | 55.91±14.37ab | — | 0.5798 |
| T8 | — | 18.63±23.58b | — | 0.1948 |
| 同列不同小写字母表示不同群落间的差异显著(P<0.05) | ||||
植物多样性与地上生物量的相关分析表明(表 4), 林龄为20年的人工林群落中草本层地上生物量与群落及草本多样性指数呈负相关, 且相关性显著(P<0.05), 说明草本层地上生物量随群落植物多样性增加而减少;灌木层地上生物量与群落及草本植物多样性指数相关关系不显著;乔木层地上生物量、地上总生物量与群落植物丰富度呈显著正相关(P<0.05), 说明人工林地上总生物量与乔木层地上生物量随群落植物丰富度增加而增大。
| 林龄/a Forest age | 地上生物量 Aboveground biomass |
群落植物多样性指数 Community plant diversity index |
草本植物多样性指数 Herb plant diversity index |
|||||||
| R | H | D | J | R | H | D | J | |||
| 20 | 草本层Herbal layer | -0.805** | -0.719** | -0.656* | -0.397 | -0.855** | -0.751** | -0.701* | -0.686* | |
| 灌木层Shrub layer | 0.221 | 0.026 | 0.067 | -0.263 | 0.143 | 0.143 | 0.22 | 0.158 | ||
| 乔木层Arbor layer | 0.618* | 0.35 | 0.361 | -0.087 | 0.532 | 0.425 | 0.469 | 0.392 | ||
| 总生物量Total biomass | 0.613* | 0.343 | 0.355 | -0.093 | 0.525 | -0.751** | 0.464 | 0.386 | ||
| 35 | 草本层Herbal layer | 0.82** | 0.67** | 0.514* | -0.333 | 0.766** | 0.235 | 0.085 | -0.135 | |
| 灌木层Shrub layer | 0.098 | -0.064 | -0.129 | -0.299 | 0.009 | 0.054 | 0.056 | 0.049 | ||
| 乔木层Arbor layer | -0.219 | -0.062 | 0.032 | 0.401 | -0.015 | 0.232 | 0.231 | 0.308 | ||
| 总生物量Total biomass | -0.205 | -0.054 | 0.036 | 0.39 | -0.003 | 0.239 | 0.235 | 0.309 | ||
| R, 丰富度指数Richness index;H, Shannon-Wiener指数Shannon-Wiener index; D, Simpson指数Simpson index; J, Pielou均匀度指数Pielou evenness index; * P < 0.05相关性显著; ** P < 0.01相关性极显著 | ||||||||||
林龄为35年的人工林群落中(表 4), 草本层地上生物量与草本层丰富度指数和群落丰富度指数、Shannon-Wiener指数、Simpson指数均呈显著正相关(P<0.05);灌木层、乔木层地上生物量和地上总生物量与群落及草本植物多样性指数相关关系不显著, 说明草本层地上生物量对群落及草本植物多样性影响较大, 总体表现为草本层地上生物量随群落植物多样性指数的增加而增大。
在植物多样性指数与地上生物量相关性分析的基础上, 对林龄为20年与林龄为35年的人工林地上生物量与植物多样性进行整体回归分析, 研究人工林从林龄20年至35年期间地上生物量与植物多样性关系整体呈现的动态变化, 得到下表 5。由表 5可以看出在回归分析中草本层地上生物量与群落及草本丰富度指数显著相关(P < 0.05), 回归系数分别为0.494, 0.440。通过最优曲线拟合得出草本层地上生物量与群落及草本丰富度回归关系符合三次函数关系, 三次函数方程分别为:
| 地上生物量 Aboveground biomass |
丰富度指数 Richness index |
Shannon-Wiener指数 Shannon-Wiener index |
Simpson指数 Simpson index |
Pielou均匀度指数 Pielou evenness index |
|||||||
| 群落 Community |
草本 Herb |
群落 Community |
草本 Herb |
群落 Community |
草本 Herb |
群落 Community |
草本 Herb |
||||
| 草本层Herbal layer | 0.494* | 0.440* | 0.300 | 0.138 | 0.301 | 0.232 | 0.109 | 0.300 | |||
| 乔木层Arbor layer | 0.511 | 0.365 | 0.003 | 0.294 | 0.001 | 0.265 | 0.107 | 0.355 | |||
| 总生物量Total biomass | 0.506 | 0.364 | 0.004 | 0.297 | 0.001 | 0.265 | 0.111 | 0.358 | |||
| * P < 0.05相关性显著;** P < 0.01相关性极显著 | |||||||||||
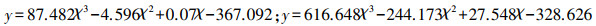
|
y为因变量即草本层地上生物量;χ为自变量为群落及草本物种丰富度。
4 结论与讨论 4.1 讨论植物多样性反映了植被群落的物种丰富程度, 生物量体现了群落的结构与功能, 能反映出植被的生长状况[22-23]。本文通过对高寒区典型人工林植被群落的植物多样性、地上生物量特征及其相互关系进行研究, 阐明了青海高寒区20年林龄人工林群落草本层地上生物量与群落及草本植物多样性指数呈负相关;35年林龄的人工林群落草本层地上生物量与草本层丰富度指数和群落植物多样性指数呈显著正相关;整体动态变化为草本层地上生物量与群落及草本丰富度呈三次函数关系。从人工林植被群落的植物多样性特征研究结果来看, 人工林群落中草本物种较灌木丰富, 且灌木多存在于阔叶人工林群落中, 这与曹广月等[24]研究一致。这可能是由于以下原因造成的:(1)灌木较草本对环境条件要求较高, 萌发所需累积的热量高, 本研究样地均选在了阴坡, 光照时间短, 光强弱, 光热少, 无法满足灌木生长需求, 但能满足草本生长需求;(2)可能是研究区位于高寒区地区, 由于海拔高, 温度低, 生长季短等原因, 灌木无法积累足够的热量支持其生长;(3)阔叶林密度较小, 叶子大而稀疏, 较针叶林林下光热条件好, 更易灌木生长。
从人工林植被群落的地上生物量特征研究结果来看, 林龄为20年的人工林地上生物量介于921.9532 —6314.9671 g/m2之间, 林龄为35年的人工林地上生物量介于9563.0731—15181.1201 g/m2之间, 且相同林龄的不同林分类型之间地上生物量差异显著, 这可能是因为组成人工林群落的树种生理生态特征不同, 导致单位时间内生物量积累速度不同。此外, 研究结果显示青海省高寒区人工林灌木层与草本层地上部分生物量占地上总生物量的比例较小, 乔木层地上生物量在群落地上部分总生物量中起主导地位。这与王丽红等[25]研究一致, 造成这种结果的主要原因是:(1)乔木层生长速度快, 形成遮荫蔽环境, 灌草生长受到抑制, 地上生物量积累受限;(2)在青海高寒区海拔高, 温度低等特殊的环境条件下, 灌、草较乔木抗逆性弱, 单位时间积累生物量少。
从植物多样性与生物量的关系研究来看, 目前许多生态学研究者就生物量与植物多样性两者关系进行了研究, 探求出的结果有正相关关系、负相关关系、单峰函数关系、不相关等[26-28]。然而由于地域不同[8]、植被类型的不同[29]、研究尺度不同[28], 往往会表现出不同的结果, 所以并未形成统一定论。在高寒地区, 主要对草甸、湿地、灌丛等植物多样性与生物量关系进行研究, 对人工林研究较少。杨路存等[1]对高寒灌丛进行了研究, 得出草本生物量与植物多样性呈显著负相关, 而灌木植物多样性与生物量并无显著相关性。武彦朋等[9]对高寒湿地植被群落进行研究, 得出在群落或区域尺度上植物多样性指数与地上生物量均呈显著负相关。牛钰杰等[5]得出在低放牧强度下植物丰富度与生物量呈负相关关系。本文对高寒区人工林植被群落进行了研究, 得出20年林龄人工林草本层地上生物量与群落及草本植物多样性指数呈负相关;35年林龄的人工林草本层地上生物量与草本层丰富度指数和群落植物多样性指数呈显著正相关;整体动态变化为草本层地上生物量与群落及草本丰富度呈三次函数关系。其中杨路存等[1]与本研究中林龄为20年的人工林群落中得出的结果较一致, 但与本文整体得出的三次函数关系不一致。导致这种结果的可能是:(1)在高寒区海拔高、温度低、土壤贫瘠的特殊环境中乔木生长较慢, 20年林龄的人工林形成的植被群落与灌丛群落发挥的生态功能相似, 得到较一致结果;(2)本文根据人工林的时间序列变化得出了草本层地上生物量与群落及草本丰富度呈三次函数关系, 而杨路存等[1]、武彦朋等[9]和牛钰杰等[5]只研究了灌丛群落某一时间的关系, 并未研究两者在时间序列的动态变化关系;(3)也可能与研究对象有关, 本文研究的是人工林, 其他三者研究的为草甸、灌丛。
目前, 许多研究者对人工林植物多样性与生物量的关系也进行了大量研究, 如太立坤等[30]和温远光等[26]研究得出桉树林植物多样性与生物量呈正相关。姚俊宇等[31]和李雁鸣等[32]认为群落层与草本层植物多样性与生物量相关性不显著。刘亚茜等[33]以不同林龄的华北落叶松纯林进行研究, 得出植物多样性和生物量之间表现为幂指数函数关系。而本研究与这些研究结果均不完全一致, 可能有以下原因:(1)本研究在高寒地区, 特殊的立地条件导致了结果差异;(2)可能是本文选择的林龄间隔较大, 且林龄序列较少。
综上所述, 本研究与国内外其他研究发现的规律不是完全相符, 可能是由于群落年龄结构、物种组成、立地条件、土壤质地、植被覆盖度等多种因素导致[25]。此外, 还需对高寒区植物多样性与生物量的关系进行更深入研究, 探明影响两者关系的限制因子及其作用机制, 从而更好地进行区域生态结构优化与调控。
4.2 结论本研究得出青海高寒区人工林植物多样性表现为草本层高于灌木层与乔木层, 群落植物多样性受草本层多样性影响较大;灌、草地上生物量占地上总生物量的比例较小, 群落地上部分总生物量主要取决于乔木层地上生物量;青海高寒区人工林群落及草本植物丰富度与草本地上生物量之间呈三次函数关系(P<0.05)。
致谢: 感谢北京林业大学张鹏、彭非、朱柱、黄乾、赵嘉伟、谭继旭、林莎、李远航在野外工作期间给予的帮助。| [1] |
杨路存, 赵玉红, 徐文华, 周国英. 青海省高寒灌丛物种多样性、生物量及其关系. 生态学报, 2018, 38(1): 309-315. |
| [2] |
冶民生, 关文彬, 谭辉, 马克明, 刘国华, 汪西林. 岷江干旱河谷灌丛α多样性分析. 生态学报, 2004, 24(6): 1123-1130. DOI:10.3321/j.issn:1000-0933.2004.06.005 |
| [3] |
王勇军, 黄从德, 张健, 杨万勤, 王宪帅. 岷江干旱河谷灌丛物种多样性、生物量及其关系. 干旱区研究, 2010, 27(4): 567-572. |
| [4] |
刘国华, 马克明, 傅伯杰, 关文彬, 康永祥, 周建云, 刘世梁. 岷江干旱河谷主要灌丛类型地上生物量研究. 生态学报, 2003, 23(9): 1757-1764. DOI:10.3321/j.issn:1000-0933.2003.09.008 |
| [5] |
牛钰杰, 杨思维, 王贵珍, 刘丽, 杜国祯, 花立民. 放牧干扰下高寒草甸物种、生活型和功能群多样性与生物量的关系. 生态学报, 2018, 38(13): 4733-4743. |
| [6] |
Wang C T, Cao G M, Wang Q L, Jing Z C, Ding L M, Long R J. Changes in plant biomass and species composition of alpine Kobresia meadows along altitudinal gradient on the Qinghai-Tibetan Plateau. Science in China Series C:Life Sciences, 2008, 51(1): 86-94. DOI:10.1007/s11427-008-0011-2 |
| [7] |
高艳, 杜峰, 王雁南. 黄土丘陵区撂荒群落地上生物量和物种多样性关系. 水土保持研究, 2017, 24(3): 96-102. |
| [8] |
Ma W H, He J S, Yang Y H, Wang X P, Liang C Z, Anwar M, Zeng H, Fang J Y, Schmid B. Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites. Global Ecology and Biogeography, 2010, 19(2): 233-243. DOI:10.1111/j.1466-8238.2009.00508.x |
| [9] |
武彦朋, 陈克龙, 张斐, 晁增国, 王永生, 陈亮, 杨龙. 青海湖内陆高寒湿地物种多样性和地上生物量的关系. 水土保持通报, 2011, 31(1): 76-80. |
| [10] |
Bai Y F, Wu J G, Pan Q M, Huang J H, Wang Q B, Li F S, Buyantuyev A, Han X G. Positive linear relationship between productivity and diversity:evidence from the Eurasian Steppe. Journal of Applied Ecology, 2007, 44(5): 1023-1034. DOI:10.1111/j.1365-2664.2007.01351.x |
| [11] |
朱桂丽, 李杰, 魏学红, 何念鹏. 青藏高寒草地植被生产力与生物多样性的经度格局. 自然资源学报, 2017, 32(2): 210-222. |
| [12] |
邹扬, 贺康宁, 赵畅, 周建, 赵丹丹, 安国才. 高寒区青海云杉人工林密度与林下植物多样性的关系. 西北植物学报, 2013, 33(12): 2543-2549. DOI:10.7606/j.issn.1000-4025.2013.12.2543 |
| [13] |
曹恒, 杨海龙, 贺康宁, 王伟伟, 高二鹏, 刘慧博, 王冰洁. 青海高寒区不同人工林配置下灌木种群的生态位研究. 西北农林科技大学学报:自然科学版, 2014, 42(5): 84-90. |
| [14] |
陈艳, 贺康宁, 伏凯, 刘静, 邹扬, 王辉. 青海大通不同树种水源涵养林对水质的影响. 水土保持学报, 2015, 29(1): 220-225. |
| [15] |
王世雷, 贺康宁, 刘可暄, 曹广月. 青海高寒区不同人工林下植被的多样性及生态位研究. 西北农林科技大学学报:自然科版, 2013, 41(11): 67-72. |
| [16] |
大通回族土族自治县志编纂委员会. 大通县志. 西安: 陕西人民出版社, 1993: 258-289.
|
| [17] |
周立华. 西宁植物志. 北京: 中国藏学出版社, 1999: 78-128.
|
| [18] |
张建军, 张守红. 水土保持与荒漠化防治实验研究方法. 北京: 中国林业出版社, 2017: 35-76.
|
| [19] |
刘琪璟. 中国立木材积表. 北京: 中国林业出版社, 2017: 1043-1052.
|
| [20] |
王斌, 刘某承, 张彪. 基于森林资源清查资料的森林植被净生产量及其动态变化研究. 林业资源管理, 2009(1): 35-43. DOI:10.3969/j.issn.1002-6622.2009.01.009 |
| [21] |
郝文芳, 陈存根, 梁宗锁, 马丽. 植被生物量的研究进展. 西北农林科技大学学报:自然科学版, 2008, 36(2): 175-182. |
| [22] |
王伟伟, 杨海龙, 贺康宁, 王瑞斌, 王辉, 王晓. 青海高寒区不同人工林配置下草本群落生态位研究. 水土保持研究, 2012, 19(3): 156-160, 165-165. |
| [23] |
郑晓翾, 靳甜甜, 木丽芬, 刘国华. 呼伦贝尔草原物种多样性与生物量、环境因子的关系. 中国草地学报, 2008, 30(6): 74-81. |
| [24] |
曹广月, 贺康宁, 胡兴波, 董梅, 贾瑞燕. 青海大通山区5种不同类型人工林下植被物种多样性比较. 广东农业科学, 2012, 39(11): 164-167. DOI:10.3969/j.issn.1004-874X.2012.11.054 |
| [25] |
王丽红, 辛颖, 邹梦玲, 赵雨森, 刘双江, 任清胜. 大兴安岭火烧迹地植被恢复中植物多样性与生物量分配格局. 北京林业大学学报, 2015, 37(12): 41-47. |
| [26] |
温远光, 陈放, 刘世荣, 梁宏温, 元昌安, 朱宏光. 广西桉树人工林物种多样性与生物量关系. 林业科学, 2008, 44(4): 14-19. |
| [27] |
陈芙蓉, 程积民, 刘伟, 朱仁斌, 杨晓梅, 赵新宇, 苏纪帅. 不同干扰对黄土区典型草原物种多样性和生物量的影响. 生态学报, 2013, 33(9): 2856-2866. |
| [28] |
任引, 彭丹, 潘俊忠, 洪志猛, 叶功富. 不同时空尺度下武夷山甜槠林物种多样性与生物量的动态关系. 林业科学, 2010, 46(8): 33-38. |
| [29] |
Adler P B, Seabloom E W, Borer E T, Hillebrand H, Hautier Y, Hector A, Harpole W S, O'Halloran L R, Grace J B, Anderson T M, Bakker J D, Biederman L A, Brown C S, Buckley Y M, Calabrese L B, Chu C J, Cleland E E, Collins S L, Cottingham K L, Crawley M J, Damschen E I, Davies K F, DeCrappeo N M, Fay P A, Firn J, Frater P, Gasarch E I, Gruner D S, Hagenah N, Hille Ris Lambers J, Humphries H, Jin V L, Kay A D, Kirkman K P, Klein J A, Knops J M H, La Pierre K J, Lambrinos J G, Li W, MacDougall A S, McCulley R L, Melbourne B A, Mitchell C E, Moore J L, Morgan J W, Mortensen B, Orrock J L, Prober S M, Pyke D A, Risch A C, Schuetz M, Smith M D, Stevens C J, Sullivan L L, Wang G, Wragg P D, Wright J P, Yang L H. Productivity is a poor predictor of plant species richness. Science, 2011, 333(6050): 1750-1753. DOI:10.1126/science.1204498 |
| [30] |
太立坤, 余雪标, 时忠杰, 周卫卫, 杨曾奖, 郭俊誉. 琼中桉树人工林植物多样性与生物量关系研究. 广东农业科学, 2009, 36(6): 143-147. DOI:10.3969/j.issn.1004-874X.2009.06.046 |
| [31] |
姚俊宇, 伍炫蓓, 孙千惠, 吴霞, 姚小兰, 郝建锋, 齐锦秋. 林窗大小对川西马尾松人工林林下物种多样性和生物量的影响. 应用与环境生物学报, 2018, 24(2): 214-220. |
| [32] |
李雁鸣, 杨德军, 张劲峰. 西南桦人工林物种多样性与生物量相关性研究. 山东林业科技, 2012, 42(6): 4-7. DOI:10.3969/j.issn.1002-2724.2012.06.002 |
| [33] |
刘亚茜, 剪文灏, 秦琰, 徐学飞, 张玉珍. 华北落叶松林生物量与生物多样性关系研究. 河北林果研究, 2010, 25(3): 223-227. DOI:10.3969/j.issn.1007-4961.2010.03.001 |