 2020, Vol. 40
2020, Vol. 40文章信息
- 李晓丽, 徐满厚, 孟万忠, 刘琦, 刘敏
- LI Xiaoli, XU Manhou, MENG Wanzhong, LIU Qi, LIU Min
- 模拟增温对云顶山亚高山草甸水热因子及群落结构的影响
- Effects of experimental warming on the hydrothermic factor and community structure of subalpine meadow on Yunding Mountain, Shanxi Province, China
- 生态学报. 2020, 40(19): 6885-6896
- Acta Ecologica Sinica. 2020, 40(19): 6885-6896
- http://dx.doi.org/10.5846/stxb201907291600
-
文章历史
- 收稿日期: 2019-07-29
- 网络出版日期: 2020-07-31

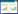




2. 太原师范学院汾河流域科学发展研究中心, 晋中 030619
2. Research center for scientific development in Fenhe River valley, Taiyuan Normal University, Jinzhong 030619, China
由温室效应不断累积所引起的气候变暖对陆地生态系统的结构和功能产生了重要影响[1]。山地空气温度随海拔升高而降低成为普遍事实, 而气候变暖也会影响山地温度的垂直变化[2-3]。2000—2015年全球变暖速率约为上世纪后半叶的一半[4], 到21世纪末全球地表温度将升高1.5—2.0℃[5], 未来40年全球变暖加速的可能性增大[6-7], 升温更为明显[8]。高海拔地区是响应气候变暖最敏感的区域之一, 其升温速率及幅度均远大于低海拔地区[9-10]。气候变暖势必对山地植被产生重要影响, 其中山地草甸植被由于分布位置的特殊性对气候变暖响应更为迅速。
目前, 关于模拟增温对草甸生态系统影响的研究增进了我们对草甸生态系统响应气候变化的理解[1], 但我国关于草甸生态系统增温的研究还相对较少, 特别是对气候变化敏感的高山草甸和亚高山草甸生态系统响应气候变暖的机理研究还很匮乏[11]。亚高山草甸是我国分布范围广泛且以多年生草本植物为主的草甸植被类型, 由于其生态环境脆弱, 极易受到气候变化及人类活动影响, 一些山地亚高山草甸已经出现物种多样性下降的情况[12]。亚高山草甸作为中低纬度、低海拔地区草地的一种组成成分, 由于植被低矮、均匀, 成为模拟增温实验理想的研究平台, 但有关中低纬度、低海拔地区亚高山草甸生态系统响应模拟增温的研究还存在许多不确定性, 制约了对草甸生态系统响应温度升高的过程与机制的理解[11, 13]。
地处我国中纬度地区的山西, 分布的亚高山草甸不仅是优良的天然牧场, 同时还是著名的生态旅游景点, 如被誉为“高原翡翠”的荷叶坪、“华北九寨沟”的舜王坪、“华北屋脊”的五台山等[2-3]。近些年来, 随着旅游业和放牧业的高度发展, 吕梁山、五台山、中条山等山系分布的亚高山草甸受人类破坏逐渐增强, 其生态环境日益敏感、脆弱[14-15]。其中, 吕梁山为山西西部山地, 其自然环境恶劣, 植被稀少, 沟壑纵横, 水土流失严重, 其森林覆盖率仅为18.5%, 是重要的生态功能区。该山系大部分山地均分布有亚高山草甸(云顶山便是其中之一), 其种类组成与周边地区相似性低, 物种多样性遭到破坏后难以恢复[16]。
众多学者从不同角度对山西亚高山草甸进行了相关研究, 主要集中在物种多样性和生物量等方面。如马丽等[17]从经度、纬度、海拔3种地理因子出发, 揭示了山西亚高山草甸植被群落地上、地下生物量及根冠比的水平、垂直空间分布特征;张世雄等[16]按照纬度梯度选取吕梁山系的马伦草原、荷叶坪和云顶山作为研究区域, 对吕梁山亚高山草甸物种多样性时空变化格局进行了研究;王誉陶等[18]以山西北部的五台山、中部的吕梁山、南部的中条山为研究区, 探讨了山西亚高山草甸物种多样性随经度、纬度、海拔的变异规律。然而, 对山西亚高山草甸进行模拟增温实验, 探讨中纬度、低海拔地区亚高山草甸生态系统对增温的响应却未见报道。唯一相似的研究为杨晓艳等[19]在吕梁山进行的模拟增温实验, 他们以吕梁山森林群落草本层植物为研究对象, 分析了草本层物种多样性的空间分布格局及其对模拟增温的响应。研究中纬度、低海拔山地亚高山草甸在模拟增温处理下的动态变化规律, 有助于深刻理解气候变暖背景下山地植被的响应机制, 具有重要理论意义。因此, 需要加强模拟增温对中纬度、低海拔山地亚高山草甸影响的研究。
云顶山作为山西低海拔地区吕梁山的中部山脉, 生境条件复杂, 孕育者大面积的亚高山草甸, 属于典型的中纬度、低海拔亚高山草甸。为防止草甸退化, 山西省人民政府于2002年6月批准建立云顶山自然保护区。但关于云顶山的研究资料较少, 仅见上官铁梁[20-21]对虎榛子灌丛、植被垂直分布, 张峰[22-23]对黄刺玫群落、华北落叶松群落, 刘明光等[24]对植物群落分类与排序等的研究报道;而关于云顶山亚高山草甸的研究, 仅见李素清等[25-26]对其种群格局和优势种群生态位的研究;对云顶山亚高山草甸进行模拟增温响应的研究却未见报道。因此, 通过在云顶山设置模拟增温实验, 分析模拟增温对草甸水热因子、植被群落特征的影响, 探究中纬度、低海拔山地亚高寒草甸对气候变暖的响应, 对该地区草甸植被的有效保护、治理恢复以及合理开发具有积极作用和重要意义。
1 研究方法 1.1 研究区概况云顶山在山西吕梁山脉中部, 位于111°35′—111°41′ E、37°46′—37°51′ N, 面积2386.3 hm2, 最高峰海拔2720 m。云顶山地势高峻, 山体由北向南倾斜, 山峰顶部呈平台状, 生境条件复杂, 属于暖温带半湿润大陆性季风气候, 夏季凉爽多雨, 冬季寒冷干燥, 年平均气温3—4℃, 7月均温16.1℃, 1月均温-10.6℃, 年降水量600—800 mm, 雨量集中在6—9月, 相对湿度70.9%, 无霜期100 d, ≥10℃年积温2100℃[27]。云顶山土壤类型从低海拔到高海拔依次为褐土、山地褐土、山地淋溶褐土、山地棕壤、亚高山草甸土。在植被区划上, 云顶山位于暖温带落叶阔叶林区, 由于地势较高, 植被的垂直分异明显, 从山麓到山顶依次为1200—1750 m落叶阔叶林带、1750—2200 m针阔叶混交林带、2200—2600 m寒温性针叶林带、2600—2720 m亚高山灌丛草甸带[20]。
1.2 样地设置于2016年8月8—10日, 在云顶山亚高山草甸选择地势平坦、植被分布均匀、人为干扰少的地段, 进行模拟增温样地布设, 样地面积70 m2, 外围由铁丝网围栏以防动物进入破坏。增温装置选择适合山地研究、无需电力维持的开顶式增温箱(Open-top chambers, OTCs)[28], 其上端开口, 箱体为正六边形, 由透明PC板材料(透光率可达95%)建构, 斜边与地面夹角保持60°(图 1)。OTC使空气温度升高, 是由于空气由其上端口进入后受到箱体的阻挡, 使水平运动减弱, 但光照可以透过箱体, 因此内部温度增加。控制箱体与地面的夹角, 保持顶部面积不变(保证进入箱体内部的光照、降水量等相等), 通过改变箱体高度和底部面积, 就可以设置内部温度。基于此, 样地设置对照(CK)、小幅度增温(OTC1)和大幅度增温(OTC2)3种实验处理[1], 每种处理3个重复, 共计9个增温样方。各样方随机布设, 间距3—5 m。CK样方:不进行增温处理, 保持植被自然状态, 取样面积1 m×1 m;OTC1样方:高度40 cm, 顶面边长25 cm, 底面边长50 cm;OTC2样方:高度80 cm, 顶面边长25 cm, 底面边长75 cm。OTC2较OTC1增大了箱体的高度和底部面积, 使其具有更高的温度, 从而实现增温梯度的设置。

|
| 图 1 增温样地设置 Fig. 1 Design of warming plots |
于2017年7月14—15日(此时已进行1年的增温实验), 对云顶山亚高山草甸进行植物群落结构特征调查, 统计草甸植物的种类、多度、高度、频度、盖度和地上生物量, 计算每个物种的重要值。采用0.5 m×0.5 m的样方框进行测定, 根据样方框大小定制网格尺寸为0.1 m×0.1 m的铁丝网, 共计25个网格, 测量时将铁丝网平铺于样方框内, 按物种进行统计。其中, 频度为物种在网格中出现的次数, 盖度为物种在网格中所占面积, 高度为物种处于自然状态下卷尺所测最高值。地上生物量采用刈割法, 将收获的植物样品按种类分别放进烘箱中在80°高温下烘干至恒重。物种学名及其分类学信息由《中国植物志》电子版(http://frps.iplant.cn/)查阅得出, 最后将所调查到的所有植物物种按照功能型划分为禾草(禾本科物种)、莎草(莎草科物种)和杂草(除禾本科和莎草科外的所有其他科物种)3种类型。
1.4 环境因子测定植被群落特征调查完毕, 于7月16—18日测量增温样方中空气-土壤水热环境因子, 所使用仪器均来自美国ONSET计算机公司生产的HOBO系列产品。空气温度和空气湿度由架设在距地面20 cm高度处的温湿度传感器(S-THB-M008)测定;土壤温度用温度传感器(S-TMB-M006)分5个深度进行测定:10、20、30、40、50 cm。所有传感器均连接到数据记录仪(H21-USB), 以30 s为时间间隔存储数据, 取1 min内的数据平均值进行分析。由于实验样地无人看守, 实验仪器不能长时间放置野外, 故空气温度、水分数据只测得调查期内的日变化数据, 即在每天的7: 40—18: 00(白天时间)进行测量, 连续观测3天, 取3天相同时刻的平均值进行空气温湿度日变化分析。土壤水分采用烘干法进行分层测定:用内径为3 cm的土钻钻取5层土样(0—10、10—20、20—30、30—40、40—50 cm), 将土样用铝盒密封, 放进105℃的烘箱中烘干至恒重, 称量后计算土壤水分。
1.5 数据分析(1) 土壤水分计算[29]
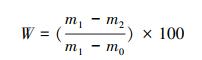
|
(1) |
式中, W为所测样品的土壤水分/%, m0为空铝盒质量/g, m1为烘干前铝盒和土样的质量/g, m2为烘干后铝盒和土样的质量/g。
(2) 物种多样性指数计算
采用α多样性指数对云顶山亚高山草甸不同增温处理下的植被物种多样性进行分析, 其指标包括:Richness指数、Simpson指数、Shannon指数、Pielou指数。计算公式如下[30]:
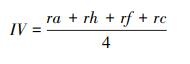
|
(2) |
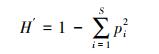
|
(3) |
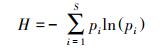
|
(4) |
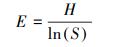
|
(5) |
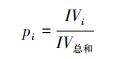
|
(6) |
式中, IV为重要值, ra为相对多度, rh为相对高度, rf为相对频度, rc为相对盖度;H′为Simpson指数, H为Shannon指数, E为Pielou指数;i 为样方框内的植物物种i, pi为第i个物种的相对重要值, S为样方框内所有植物种类的总和。
(3)统计分析
在不同时空尺度上, 用OriginPro 9.1软件分析不同增温处理下云顶山亚高山草甸空气-土壤水热因子和植被群落结构特征的变化;同时采用IBM SPSS Statistics 22.0软件中的GLM(General Linear Model)模型, 分别对草甸土壤水热因子和植被群落结构特征在不同处理、深度、功能型间的差异进行双因素方差分析。对于草甸不同增温处理下温度、水分、植被关系, 首先利用Canoco for Windows 4.5软件对植被群落结构特征数据进行DCA(Detrended Correspondence Analysis)分析, 确定用线性模型进行直接梯度分析(4个排序轴的最大梯度长度小于3), 再选择其中的RDA(Redundancy Analysis)模型进行约束性排序分析。
2 结果与分析 2.1 空气温湿度对增温的响应空气温度随实验时间延长呈减小趋势, 随增温幅度升高而增大(图 2)。不同增温处理下空气温度差异显著(P < 0.05), 在OTC1处理下显著增加3.57℃, 在OTC2处理下显著增加5.04℃(表 1)。空气湿度的变化趋势与空气温度相反, 即随实验时间延长呈增加趋势, 随增温幅度升高而减小(图 2)。空气湿度在OTC1处理下显著减小7.36%, 在OTC2处理下显著减小5.23%(P < 0.05)(表 1)。因此, 增温处理增加空气温度, 减小空气湿度, 使云顶山亚高山草甸气候呈现暖干化。

|
| 图 2 不同增温处理下空气温湿度的日变化 Fig. 2 Trends of air temperature and humidity with daytime in different warming treatments CK:对照, Control;OTC1:小幅度增温, Small-warming amplitude;OTC2:大幅度增温, Large-warming amplitude |
| 水热因子Hydrothermal factor | CK | OTC1 | OTC2 |
| 空气温度Air temperature | 16.18±0.02c | 19.75±0.10b | 21.22±0.04a |
| 空气湿度Air humidity | 77.04±0.09a | 69.68±0.19c | 71.81±0.18b |
| 土壤温度Soil temperature | 13.38±0.54b | 13.33±0.67b | 13.64±0.75a |
| 土壤水分Soil moisture | 30.26±0.98a | 30.06±1.19a | 30.88±1.55a |
| CK:对照, Control;OTC1:小幅度增温, Small-warming amplitude;OTC2:大幅度增温, Large-warming amplitude;字母相同:方差分析结果不显著(P>0.05);字母不同:方差分析结果显著(P < 0.05) | |||
另外, 在一天内的不同时刻, 空气湿度在8:40之后呈增大趋势, 同时该时刻的空气温度开始出现下降, 对照当时的天气情况, 是由于此时开始降雨所致。空气温湿度在12:00和15:00再次出现波动, 是由于降雨在12:00突然变小, 甚至暂停, 而后在15:00再次变大。这表明云顶山亚高山草甸由于受海拔影响, 其气候日变化波动较大。
2.2 土壤温湿度对增温的响应土壤温度随深度增加而减小(P < 0.001), 随增温幅度先减小后增大(P=0.006), 其变化趋势均达到极显著水平(P < 0.01);不同深度的土壤温度在处理间也达到显著差异(P=0.028), 即增温能够显著影响0—50 cm深度范围内的土壤温度(图 3、表 2)。总体而言, OTC1处理下, 土壤温度减小0.05℃;OTC2处理下, 土壤温度增加0.26℃;OTC2处理下的土壤温度均显著高于对照和OTC1处理(表 1)。

|
| 图 3 不同增温处理下土壤温湿度随深度的变化趋势 Fig. 3 Trends of soil temperature and moisture with depth in different warming treatments |
| 因素Factor | 土壤温度Soil temperature | 土壤水分Soil moisture |
| 处理Treatment | 0.006 | 0.285 |
| 深度Depth | 0.000 | 0.000 |
| 深度×处理Depth×Treatment | 0.028 | 0.413 |
土壤水分随深度增加极显著减小(P < 0.001), 随增温幅度未达到显著差异(P=0.285), 且不同深度的土壤水分在处理间的差异也未达到显著水平(P=0.413), 表明土壤水分对增温响应不敏感(图 3、表 2)。在不同增温处理下, 土壤水分具有先减小后增加的变化趋势, 即在OTC1处理下减小0.20%, 在OTC2处理下增加0.62%(表 1)。
因此, 云顶山亚高山草甸土壤温湿度对深度响应敏感, 随深度增加均显著减小;但对增温处理响应差异较大, 土壤温度显著增加, 土壤水分变化却不显著。
2.3 植被物种多样性对增温的响应Richness指数、Simpson指数、Shannon指数和Pielou指数随增温幅度均表现为递减趋势, 除Pielou指数在OTC2处理下显著低于对照处理(P < 0.05)外, 其余指数在不同处理间的差异均不显著(P>0.05), 这表明增温处理对物种多样性产生了一定负面影响(图 4)。

|
| 图 4 植被物种多样性对增温的响应 Fig. 4 Responses of plant species diversity to warming |
将不同增温样地中所调查的所有物种分成杂草类、禾草类和莎草类三种植物功能型, 分别测定其物种数、地上生物量和重要值(图 5)。在不同处理间, 地上生物量达到极显著差异(P < 0.01), 且随增温幅度递增, 表明增温处理增加了地上生物量;但物种数和重要值均未达到显著性差异(P>0.05), 表明物种多样性对增温处理响应不敏感(表 3)。

|
| 图 5 不同植物功能型生长特征对增温的响应 Fig. 5 Responses of growth characteristics of different plant functional types to warming |
| 因素Factor | 变量Variable | P | 多重比较Multiple comparison | ||
| 处理 | CK | OTC1 | OTC2 | ||
| Treatment | 物种数 | >0.05 | a | a | a |
| 地上生物量 | < 0.001 | c | b | a | |
| 重要值 | >0.05 | a | a | a | |
| 植物功能型 | 杂草 | 禾草 | 莎草 | ||
| Plant functional type | 物种数 | < 0.001 | a | b | b |
| 地上生物量 | < 0.001 | a | a | b | |
| 重要值 | < 0.001 | a | b | c | |
| 处理×植物功能型 | 物种数 | >0.05 | |||
| Treatment×Plant functional type | 地上生物量 | < 0.001 | |||
| 重要值 | < 0.001 | ||||
在不同植物功能型间, 物种数、地上生物量和重要值均达到极显著差异(P < 0.01), 且杂草的这三个指标最高, 禾草次之, 莎草最低, 这表明草甸群落中以杂草类植物为优势种(表 3)。
杂草、禾草、莎草的物种数在不同处理间差异不显著(P>0.05), 但其地上生物量和重要值在不同处理间均达到极显著差异(P < 0.01), 表明增温处理对不同植物功能型产生了影响(表 3)。在OTC1处理下, 莎草的物种数、地上生物量、重要值均增加;但在OTC2处理下, 已不存在莎草类植物(图 5)。随增温幅度, 杂草的地上生物量逐渐增加, 但其重要值和物种数逐渐降低。禾草的物种数、重要值和地上生物量随增温幅度均逐渐增加。因此, 增温处理促进了群落中禾草类植物的生长, 抑制了杂草类植物的生长。
2.5 植被生长指标与水热因子的关系通过RDA排序分析得到(图 6), CK样地(第三象限)中以杂草居多, 杂草的物种数和重要值与各物种多样性指数、空气湿度具有较强正相关性。在OTC1样地(第二象限), 莎草为优势种, 其物种数和重要值与30—40 cm深度土壤水分具较强正相关性。在OTC2样地(第一象限), 优势种为禾草, 其物种数和重要值与地上生物量、空气温度、10 cm深度土壤温度、0—10 cm深度土壤水分具有较强的正相关性。禾草类植物可导致较高地上生物量, 杂草类植物可导致较高物种多样性。随增温幅度, 群落中不同植物功能型由杂草类向禾草类转化。

|
| 图 6 植被指标与水热因子的RDA排序分析 Fig. 6 RDA ordination between vegetation indices and hydrothermal factors AT:空气温度, Air temperature;ST10:10 cm土壤温度, Soil temperature at 10 cm depth;ST20:20 cm土壤温度, Soil temperature at 20 cm depth;ST30:30 cm土壤温度, Soil temperature at 30 cm depth;ST40:40 cm土壤温度, Soil temperature at 40 cm depth;ST50:50 cm土壤温度, Soil temperature at 50 cm depth;AH:空气湿度, Air humidity;SW0—10:0—10 cm土壤水分, Soil moisture at 0—10 cm depth;SW10—20:10—20 cm土壤水分, Soil moisture at 10—20 cm depth;SW20—30:20—30 cm土壤水分, Soil moisture at 20—30 cm depth;SW30—40:30—40 cm土壤水分, Soil moisture at 30—40 cm depth;SW40—50:40—50 cm土壤水分, Soil moisture at 40—50 cm depth;Richness:Richness指数, Richness index;Simpson:Simpson指数, Simpson index;Shannon:Shannon指数, Shannon index;Pielou:Pielou指数, Pielou index;AGB:地上生物量, Aboveground biomass;HIV:禾草重要值, Importance value of grass;HSN:禾草物种数, Species number of grass;SIV:莎草重要值, Importance value of sedge;SSN:莎草物种数, Species number of sedge;WIV:杂草重要值, Importance value of forb;WSN:杂草物种数, Species number of forb;CK1—3:1—3号CK样地, Control plots from 1 to 3;LW1—3:1—3号OTC1样地, Small-warming amplitude plots from 1 to 3;HW1—3:1—3号OTC2样地, Large-warming amplitude plots from 1 to 3 |
通过相关分析得到(表 4), 空气温度(RAT=-0.857, P < 0.01)、10 cm深度土壤温度(RST10=-0.822, P < 0.01)与Pielou指数呈极显著负相关关系, 与地上生物量呈极显著正相关关系(RAT=0.823, RST10=0.818, P < 0.01), 表明空气、浅层土壤温度能够降低物种多样性, 增加生产力;与禾草物种数(RAT=0.849, RST10=0.801, P < 0.01)、重要值(RAT=0.870, RST10=0.817, P < 0.01)的关系为极显著正相关, 与杂草重要值的关系为显著负相关(RAT=-0.780, RST10=-0.705, P < 0.05), 再次表明空气、浅层土壤温度可促进禾草生长, 抑制杂草生长。30 cm(RST30=-0.685, P < 0.05)、40 cm(RST40=-0.675, P < 0.05)土壤温度与莎草重要值呈显著负相关关系, 表明深层土壤温度能够抑制莎草生长。0—10 cm深度土壤水分与Pielou指数呈显著负相关关系(RSW0—10=-0.703, P < 0.05), 与地上生物量呈显著正相关关系(RSW0-10=0.686, P < 0.05), 表明浅层土壤水分能够降低物种多样性, 增加生产力;与禾草重要值的关系为显著正相关(RSW0-10=0.735, P < 0.05), 表明浅层土壤水分促进禾草生长。
| Richness | Simpson | Shannon | Pielou | AGB | WSN | HSN | SSN | WIV | HIV | SIV | |
| AT | -0.07 | -0.522 | -0.387 | -0.857** | 0.823** | -0.309 | 0.849** | -0.319 | -0.780* | 0.870** | -0.421 |
| AH | -0.063 | 0.353 | 0.208 | 0.588 | -0.414 | 0.243 | -0.607 | -0.118 | 0.594 | -0.548 | 0.101 |
| ST10 | -0.138 | -0.515 | -0.41 | -0.822** | 0.818** | -0.321 | 0.801** | -0.414 | -0.705* | 0.817** | -0.442 |
| ST20 | -0.469 | -0.364 | -0.449 | -0.247 | 0.416 | -0.379 | 0.271 | -0.549 | -0.258 | 0.467 | -0.485 |
| ST30 | 0.126 | 0.295 | 0.216 | 0.127 | 0.175 | 0.208 | 0.398 | -0.628 | 0.069 | 0.298 | -0.685* |
| ST40 | 0.191 | 0.234 | 0.192 | -0.065 | 0.361 | 0.171 | 0.564 | -0.544 | -0.138 | 0.465 | -0.675* |
| ST50 | 0.181 | 0.439 | 0.333 | 0.346 | -0.048 | 0.281 | 0.146 | -0.395 | 0.22 | 0.061 | -0.471 |
| SW0—10 | -0.15 | -0.424 | -0.366 | -0.703* | 0.686* | -0.23 | 0.638 | -0.496 | -0.583 | 0.735* | -0.479 |
| SW10—20 | 0.089 | -0.024 | 0.024 | -0.125 | -0.122 | 0.048 | -0.116 | 0.234 | 0.127 | -0.226 | 0.231 |
| SW20—30 | -0.354 | -0.331 | -0.327 | -0.065 | -0.136 | -0.271 | -0.273 | 0.059 | 0.021 | -0.172 | 0.298 |
| SW30—40 | -0.516 | -0.497 | -0.497 | -0.118 | -0.172 | -0.539 | -0.205 | 0.244 | -0.219 | -0.038 | 0.426 |
| SW40—50 | -0.337 | -0.332 | -0.346 | -0.265 | 0.283 | -0.208 | 0.233 | -0.6 | -0.194 | 0.347 | -0.356 |
| Richness:Richness指数, Richness index;Simpson:Simpson指数, Simpson index;Shannon:Shannon指数, Shannon index;Pielou:Pielou指数, Pielou index;AGB:地上生物量, Aboveground biomass;WSN:杂草物种数, Species number of forb;HSN:禾草物种数, Species number of grass;SSN:莎草物种数, Species number of sedge;WIV:杂草重要值, Importance value of forb;HIV:禾草重要值, Importance value of grass;SIV:莎草重要值, Importance value of sedge;AT:空气温度, Air temperature;AH:空气湿度, Air humidity;ST10:10 cm土壤温度, Soil temperature at 10 cm depth;ST20:20 cm土壤温度, Soil temperature at 20 cm depth;ST30:30 cm土壤温度, Soil temperature at 30 cm depth;ST40:40 cm土壤温度, Soil temperature at 40 cm depth;ST50:50 cm土壤温度, Soil temperature at 50 cm depth;SW0—10:0—10 cm土壤水分, Soil moisture at 0—10 cm depth;SW10—20:10—20 cm土壤水分, Soil moisture at 10—20 cm depth;SW20—30:20—30 cm土壤水分, Soil moisture at 20—30 cm depth;SW30—40:30—40 cm土壤水分, Soil moisture at 30—40 cm depth;SW40—50:40—50 cm土壤水分, Soil moisture at 40—50 cm depth;*:P < 0.05;**:P < 0.01 | |||||||||||
OTC限制了水平方向的空气流动, 一定程度也降低了垂直方向的对流过程, 室内空气湍流作用减弱, 加之太阳辐射很容易透过玻璃纤维, 导致温室内的温度升高[1, 31-33]。在温度升高下, 植物蒸腾增强, 植物生理过程发生改变, 同时温室内的季节冻土层发生消融与融冻等过程, 以及土壤其他物理条件发生改变, 必然对植物的生长和演替产生影响[34]。本研究得到, 随增温幅度变大, 云顶山亚高山草甸物种多样性呈递减趋势, 说明增温对物种多样性产生一定负面影响。这是由于增温使局部小气候出现暖干化, 从而影响了植被的生长。本研究也得到, 云顶山亚高山草甸空气在增温后出现暖干化, 表现为空气温度显著增加, 空气湿度显著减小, 直接影响地上植被的生长, 对群落结构产生负面作用。杨晓艳等[19]在吕梁山进行的模拟增温实验也表明, 适度增温可提高物种多样性, 但持续增温会使多样性受到抑制, 这与徐满厚等[35-37]在青藏高原的研究结果相同, 即高度增温可抑制高寒植被生长。云顶山亚高山草甸由于受到地形和海拔因素影响, 该区表现为严寒微湿气候, 形成与青藏高原高寒草甸相类似的气候。因此, 在增温下云顶山亚高山草甸和青藏高原高寒草甸表现为较为一致的变化趋势。李娜等[1]的研究也表明, OTC1可使高寒草甸杂草分盖度显著增加, OTC2则抑制增加趋势。他们认为, 草甸土壤条件和水热状况不同, 可导致植被物种多样性对增温响应表现不同。因此, 草甸水热因子对群落物种多样性的形成和维持起至关重要的作用。
高寒植被各功能群植物由于生物学特性不同, 其生长特征对增温响应也不同[38-39]。本研究将在云顶山亚高山草甸所调查的物种分成禾草、莎草和杂草2种功能型, 得到增温促进了草甸禾草类植物生长, 抑制了杂草类植物生长, 且随增温幅度变大, 群落中不同植物功能型由杂草类向禾草类转化。周华坤等[31]在海北高寒草甸的研究得出, 增温1年后禾草地上生物量增加了12.30%, 莎草增加了1.18%, 杂草减少了21.13%。石福孙等[32]在川西北亚高山草甸的研究也得到相似的结论, OTC内禾草的地上生物量和盖度显著增大, 杂草的地上生物量和盖度显著减小。同样, 姜炎彬等[34]和李英年等[33]均得到, 增温后禾草物种数和盖度增加, 杂草物种数和盖度减小。但宗宁等[40]在藏北高寒草甸的研究发现, 增温降低了禾草与莎草植物盖度。李娜等[1]在青藏高原的研究也有类似的结论, 增温使高寒草甸禾草和莎草盖度减少, 杂草盖度增加, 而使沼泽草甸中禾草和莎草盖度增加, 杂草盖度减少。分析认为, 增温后直接导致了OTC内土壤含水量的减少, 继而植物种间竞争关系被破坏, 从而引起群落优势种和组成发生改变, 由于土壤条件和水热条件不同, 对增温的响应也表现出不一致[1]。本研究得到, 在OTC1处理下, 莎草的物种数、地上生物量、重要值均增加;但在OTC2处理下, 莎草类植物消失;随增温幅度变大, 杂草的地上生物量逐渐增加, 但其重要值和物种数逐渐降低;禾草的物种数、重要值和地上生物量随增温幅度均逐渐增加。因此, 短期增温能促进禾草植物生长, 抑制杂草植物生长。究其原因可能是, 禾草在增温条件下分蘖能力的增强和资源分配模式的改变延长了其对土壤元素的获得时期, 使其生长较快;而莎草较禾草耐阴, 当禾草占据群落上层时形成郁闭环境, 莎草便成为群落下层, 同时莎草一般为短根茎地下芽植物, 与禾草竞争吸收土壤氮素的能力不高, 所以莎草生长较慢;杂草则由于禾草和莎草与之过分竞争了光照和养分, 生长受抑制[1, 31-34]。可见, 引起禾草、莎草、杂草在增温下产生分化的主要原因是, 各类功能型植物的生物学特性及其对资源利用的差异性。本研究对云顶山亚高山草甸水热因子与群落结构特征关系进行的RDA排序分析, 验证了这一结论, 得到空气、浅层土壤温度促进禾草生长, 抑制杂草生长;深层土壤温度抑制莎草生长;浅层土壤水分促进禾草生长。因此, 增温改变了云顶山亚高山草甸的水热因子状况, 导致草甸群落结构发生改变, 使之向禾草类植物进行演替。
同样, 国外也有研究发现, 受全球变暖影响, 动植物的体型普遍在“缩水”, 全球平均气温每上升1℃, 植物体型可能缩小3—17%[41]。由温度升高引起的植物早衰现象降低了植物蒸腾速率, 在一定程度上间接增加了土壤湿度[42];但大尺度下气候变暖引起的干旱胁迫通过改变群落组成物种(优势种)的丰富度和稳定性, 直接影响到整个植被群落的稳定性[43]。增温可通过改变土壤水分对群落的结构和组成产生影响, 但不同物种和生活型对增温有着不同的反应[44]。研究表明, 增温改变了亚高山草甸植物群落的物种组成, 增加了禾草和豆科植物在亚高山草甸植物群落中的比例, 却降低了其他阔叶草本植物在群落中的比例[45]。Hoeppner和Dukes[46]认为, 温度增加会显著降低物种丰富度, 显著改变功能群组成, 使C4禾草以及一、二年生植物进入到以多年生C3植物为主导的群落。总之, 温度变化已经显著影响了草地植物群落物种组成和植物多样性, 从而进一步影响植被群落生产力的形成。
4 结论在云顶山亚高山草甸进行模拟增温实验, 温度升高下草甸空气、土壤、植物群落发生较大变化。在不同增温处理下, 空气温度显著增加, 空气湿度显著减小, 气候呈现暖干化;土壤温度显著增加, 土壤水分趋于增加, 土壤趋向暖湿化。草甸植物群落在增温处理下杂草类植物生长受到抑制, 禾草类植物生长得以促进;禾草类植物可导致群落具有较高的地上生物量, 杂草类植物可导致较高的物种多样性。因此, 增温增加了草甸植物群落地上生物量, 但对其物种多样性产生负面影响。随温度持续升高, 草甸中不同植物功能型由杂草类向禾草类转化。这可能是由于在增温处理下, 空气、浅层土壤温度增加促进禾草生长、抑制杂草生长, 深层土壤温度增加抑制莎草生长, 从而导致整个群落向禾草类植物进行演替。
| [1] |
李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895-905. |
| [2] |
Xu M H, Ma L, Jia Y Y, Liu M. Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS One, 2017, 12(3): e0174231. DOI:10.1371/journal.pone.0174231 |
| [3] |
Xu M H, Zhang S X, Wen J, Yang X Y. Multiscale spatial patterns of species diversity and biomass together with their correlations along geographical gradients in subalpine meadows. PLoS One, 2019, 14(2): e0211560. DOI:10.1371/journal.pone.0211560 |
| [4] |
Karl T R, Arguez A, Huang B Y, Lawrimore J H, McMahon J R, Menne M J, Peterson T C, Vose R S, Zhang H M. Possible artifacts of data biases in the recent global surface warming hiatus. Science, 2015, 348(6242): 1469-1472. DOI:10.1126/science.aaa5632 |
| [5] |
Held I M. Climate science:the cause of the pause. Nature, 2013, 501(7467): 318-319. DOI:10.1038/501318a |
| [6] |
Roberts C D, Palmer M D, McNeall D, Collins M. Quantifying the likelihood of a continued hiatus in global warming. Nature Climate Change, 2015, 5(4): 337-342. DOI:10.1038/nclimate2531 |
| [7] |
Smith S J, Edmonds J, Hartin C A, Mundra A, Calvin K. Near-term acceleration in the rate of temperature change. Nature Climate Change, 2015, 5(4): 333-336. DOI:10.1038/nclimate2552 |
| [8] |
Marotzke J, Forster P M. Forcing, feedback and internal variability in global temperature trends. Nature, 2015, 517(7536): 565-570. DOI:10.1038/nature14117 |
| [9] |
Thomas C D, Cameron A, Green R E, Bakkenes M, Beaumont L J, Collingham Y C, Erasmus B F N, de Siqueira M F, Grainger A, Hannah L, Hughes L, Huntley B, van Jaarsveld A S, Midgley G F, Miles L, Ortega-Huerta M A, Townsend Peterson A, Phillips O L, Williams S E. Extinction risk from climate change. Nature, 2004, 427(6970): 145-148. DOI:10.1038/nature02121 |
| [10] |
Peñuelas J, Sardans J, Estiarte M, Ogaya R, Carnicer J, Coll M, Barbeta A, Rivas-Ubach A, Llusià J, Garbulsky M, Filella I, Jump A S. Evidence of current impact of climate change on life:A walk from genes to the biosphere. Global Change Biology, 2013, 19(8): 2303-2338. DOI:10.1111/gcb.12143 |
| [11] |
侯彦会, 周广胜, 许振柱. 基于红外增温的草地生态系统响应全球变暖的研究进展. 植物生态学报, 2013, 37(12): 1153-1167. |
| [12] |
白家烨, 刘卫华, 赵冰清, 张青, 郭东罡. 芦芽山荷叶坪亚高山草甸生物多样性. 应用生态学报, 2018, 29(2): 389-396. |
| [13] |
叶旺敏, 熊德成, 杨智杰, 朱益广, 张秋芳, 刘小飞, 林伟盛, 胥超, 张景, 杨玉盛. 模拟增温对杉木幼树生长和光合特性的影响. 生态学报, 2019, 39(7): 2501-2509. |
| [14] |
卢爱英, 张先平, 王世裕, 王孟本. 干扰对云顶山亚高山草甸群落物种多样性的影响. 植物研究, 2011, 31(1): 73-78. |
| [15] |
章异平, 江源, 刘全儒, 任斐鹏. 放牧对五台山高山、亚高山草甸牧草品质的影响. 生态学报, 2011, 31(13): 3659-3667. |
| [16] |
张世雄, 杨晓艳, 温静, 徐满厚. 山西吕梁山亚高山草甸物种多样性的时空变化格局. 生态学报, 2018, 38(18): 6685-6693. |
| [17] |
马丽, 徐满厚, 周华坤, 贾燕燕, 王孝贤, 席文涛. 山西亚高山草甸植被生物量的地理空间分布. 生态学杂志, 2018, 37(8): 2244-2253. |
| [18] |
王誉陶, 毕玉婷, 王倩, 郝海超, 徐满厚. 山西亚高山草甸植物群落物种多样性的空间分异. 中国农学通报, 2018, 34(18): 77-83. |
| [19] |
杨晓艳, 张世雄, 温静, 徐满厚. 吕梁山森林群落草本层植物物种多样性的空间格局及其对模拟增温的响应. 生态学报, 2018, 38(18): 6642-6654. |
| [20] |
上官铁梁, 张峰. 云顶山植被及其垂直分布研究. 山地研究, 1991, 9(1): 26-29. |
| [21] |
上官铁梁, 张峰. 云顶山虎榛子灌丛群落学特性及生物量. 山西大学学报:自然科学版, 1989, 12(3): 347-352. |
| [22] |
张峰, 上官铁梁. 关帝山黄刺玫灌丛群落结构与生物量的研究. 武汉植物学研究, 1991, 9(3): 247-252. |
| [23] |
张峰, 上官铁梁. 关帝山华北落叶松林的群落学特征和生物量. 山西大学学报:自然科学版, 1992, 15(1): 72-77. |
| [24] |
刘明光, 刘莹, 张峰, 梁小明. 云顶山自然保护区植物群落的分类与排序. 林业资源管理, 2011(4): 82-88. |
| [25] |
李素清, 张金屯. 山西云顶山亚高山草甸群落生态分析. 地理研究, 2007, 26(1): 83-90. |
| [26] |
李素清, 杨斌胜, 张金屯. 山西云顶山亚高山草甸优势种群和群落的格局分析. 应用与环境生物学报, 2007, 13(1): 9-13. |
| [27] |
李斌, 李素清, 张金屯. 云顶山亚高山草甸优势种群生态位研究. 草业学报, 2010, 19(1): 6-13. |
| [28] |
Wang J F, Wu Q B. Impact of experimental warming on soil temperature and moisture of the shallow active layer of wet meadows on the Qinghai-Tibet Plateau. Cold Regions Science and Technology, 2013, 90-91: 1-8. DOI:10.1016/j.coldregions.2013.03.005 |
| [29] |
Lang L Z, Xiang W, Huang W, Cui D S, Schanz T. An experimental study on oven-drying methods for laboratory determination of water content of a calcium-rich bentonite. Applied Clay Science, 2017, 150: 153-162. DOI:10.1016/j.clay.2017.09.022 |
| [30] |
Xu M H, Peng F, You Q G, Guo J, Tian X F, Liu M, Xue X. Effects of warming and clipping on plant and soil properties of an alpine meadow in the Qinghai-Tibetan Plateau, China. Journal of Arid Land, 2015, 7(2): 189-204. DOI:10.1007/s40333-014-0010-z |
| [31] |
周华坤, 周兴民, 赵新全. 模拟增温效应对矮嵩草草甸影响的初步研究. 植物生态学报, 2000, 24(5): 547-553. |
| [32] |
石福孙, 吴宁, 罗鹏. 川西北亚高山草甸植物群落结构及生物量对温度升高的响应. 生态学报, 2008, 28(11): 5286-5293. |
| [33] |
李英年, 赵亮, 赵新全, 周华坤. 5年模拟增温后矮蒿草草甸群落结构及生产量的变化. 草地学报, 2004, 12(3): 236-239. |
| [34] |
姜炎彬, 范苗, 张扬建. 短期增温对藏北高寒草甸植物群落特征的影响. 生态学杂志, 2017, 36(3): 616-622. |
| [35] |
Xu M H, Peng F, You Q G, Guo J, Tian X F, Liu M, Xue X. Initial effects of experimental warming on temperature, moisture, and vegetation characteristics in an alpine meadow on the Qinghai-Tibetan Plateau. Polish Journal of Ecology, 2014, 62(3): 491-509. DOI:10.3161/104.062.0310 |
| [36] |
徐满厚, 薛娴. 青藏高原高寒草甸植被特征与温度、水分因子关系. 生态学报, 2013, 33(10): 3158-3168. |
| [37] |
徐满厚, 薛娴. 青藏高原高寒草甸夏季植被特征及对模拟增温的短期响应. 生态学报, 2013, 33(7): 2071-2083. |
| [38] |
徐满厚, 薛娴. 气候变暖对陆地植被-土壤生态系统的影响研究. 生命科学, 2012, 24(5): 492-500. |
| [39] |
马丽, 徐满厚, 翟大彤, 贾燕燕. 高寒草甸植被-土壤系统对气候变暖响应的研究进展. 生态学杂志, 2017, 36(6): 1708-1717. |
| [40] |
宗宁, 柴曦, 石培礼, 蒋婧, 牛犇, 张宪洲, 何永涛. 藏北高寒草甸群落结构与物种组成对增温与施氮的响应. 应用生态学报, 2016, 27(12): 3739-3748. |
| [41] |
Sheridan J A, Bickford D. Shrinking body size as an ecological response to climate change. Nature Climate Change, 2011, 1(8): 401-406. DOI:10.1038/nclimate1259 |
| [42] |
Zavaleta E S, Thomas B D, Chiariello N R, Asner G P, Shaw M R, Field C B. Plants reverse warming effect on ecosystem water balance. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(17): 9892-9893. DOI:10.1073/pnas.1732012100 |
| [43] |
Dieleman C M, Branfireun B A, McLaughlin J W, Lindo Z. Climate change drives a shift in peatland ecosystem plant community:implications for ecosystem function and stability. Global Change Biology, 2015, 21(1): 388-395. |
| [44] |
Harte J, Shaw R. Shifting dominance within a montane vegetation community:results of a climate-warming experiment. Science, 1995, 267(5199): 876-880. DOI:10.1126/science.267.5199.876 |
| [45] |
Weltzin J F, Pastor J, Harth C, Bridgham S D, Updegraff K, Chapin C T. Response of bog and fen plant communities to warming and water-table manipulations. Ecology, 2000, 81(12): 3464-3478. DOI:10.1890/0012-9658(2000)081[3464:ROBAFP]2.0.CO;2 |
| [46] |
Hoeppner S S, Dukes J S. Interactive responses of old-field plant growth and composition to warming and precipitation. Global Change Biology, 2012, 18(5): 1754-1768. DOI:10.1111/j.1365-2486.2011.02626.x |