 2020, Vol. 40
2020, Vol. 40文章信息
- 王丽虹, 杨磊, 刘玲, 何亮, 蒋万祥, 申恒伦, 朱天顺, 潘保柱
- WANG Lihong, YANG Lei, LIU Ling, HE Liang, JIANG Wanxiang, SHEN Henglun, ZHU Tianshun, PAN Baozhu
- 南四湖沉水植物物种多样性和功能多样性对水深梯度的响应
- Species diversity and functional diversity of submerged vegetation community in response to water depth gradient in Nansi Lake, China
- 生态学报. 2020, 40(17): 6233-6242
- Acta Ecologica Sinica. 2020, 40(17): 6233-6242
- http://dx.doi.org/10.5846/stxb201904100715
-
文章历史
- 收稿日期: 2019-04-10
- 网络出版日期: 2020-07-10






2. 南昌大学鄱阳湖环境与资源利用教育部重点实验室, 南昌 330031;
3. 西安理工大学水利水电学院, 西安 710048
2. Key Laboratory of Poyang Lake Environment and Resource Utilization, Ministry of Education, Nanchang University, Nanchang 330031, China;
3. Institute of Water Resources and Hydro-electric Engineering, Xi'an University of Technology, Xi'an 710048, China
近年来, 物种多样性和功能多样性作为重要研究手段被广泛应用于生态学研究的各个领域, 为揭示生态系统物种共存及群落构建机制提供了独特视角[1-3]。物种多样性和功能多样性对生态系统结构和功能起决定性作用, 是生物多样性在不同层次上的表现形式[4-6]。功能多样性结合了群落物种组成、多度以及种间功能性状参数, 能够综合表征群落总体信息[7-10]。物种多样性、功能多样性以及二者关系随环境梯度的响应研究, 有助于理解不同环境条件下物种的适应策略和群落构建的生态过程[11-14]。然而, 不同环境尺度下, 不同测度的物种多样性和功能多样性指数(优势度、丰富度、多样性、均匀度)对环境变化的响应仍无统一定论[1, 7, 15]。此外, 物种多样性和功能多样性关系可表现为正相关、负相关或不相关[16-18]。因此, 群落水平的物种多样性和功能多样性对环境梯度的响应关系仍值得进一步研究。
物种多样性和功能多样性对环境梯度的响应研究最早源于陆生植物群落, 近年来被广泛应用于淡水生态学各研究领域, 有力地促进了相关理论的发展, 成为水域生态学研究的热点之一[19-21]。沉水植物是湖泊重要的初级生产者之一, 对湖泊生态系统结构和功能维持发挥着关键性作用[21-22]。水深是影响沉水植物生长、繁殖与分布的重要环境因子。水深变化可引起水体温度、光照、风浪、溶解氧、底质等众多环境因子的关联变化, 驱动着植物群落的构建过程[22-23]。关于水深对沉水植物影响的研究已有报道, 水深变化可引起株高、分株数、比叶面积、器官生物量、化学计量学特征等功能性状的改变[24-26]。然而, 这些研究大多基于个体或物种水平, 而有关群落层次的功能性状、物种多样性、功能多样性及其关系对水深的响应研究鲜有报道。
南四湖是南水北调东线工程重要调蓄湖泊, 20世纪60年代修建的二级坝水利枢纽将南四湖拦腰分隔为上、下两级湖区。调水工程的运行改变了湖泊原有的水文节律, 调水期间上、下级湖区水位比年均水位分别提高了0.5 m和1.1 m。基于此背景下开展沉水植物群落对水深梯度响应研究兼具理论和实际意义。本文拟对南四湖不同水深梯度下沉水植物群落物种多样性、功能多样性以及二者关系对水深梯度的响应进行分析, 以期回答以下问题:(1)随水深梯度变化, 群落功能性状如何响应?(2)水深对物种多样性和功能多样性有何影响?(3)物种多样性和功能多样性关系是否会随水深的变化而改变?以上科学问题的回答, 不仅为沉水植物物种多样性和功能多样性的深入研究提供科学依据, 也为调蓄湖群沉水植物多样性保护与管理提供理论指导。
1 研究区域概况南四湖(116°34′ E—117°21′ E, 34°27′ N—35°20′ N)位于山东省西南部, 华北地区最大的淡水湖泊, 是南阳湖、独山湖、昭阳湖和微山湖4个湖泊的总称。南北长约126 km, 东西宽约5—25 km, 湖面面积1266 km2, 总容积47.3亿m3。1960年修建的二级坝将南四湖拦腰分为上、下两级湖区。上级湖允许最高水位36.5 m, 兴利水位34.2 m, 死水位33.0 m;下级湖允许最高水位35.0 m, 兴利水位32.5 m, 死水位31.5 m。南水北调东线一期工程运行期间, 上、下级湖区水位比年均水位提高了0.5—1.0 m。南四湖现有沉水植物6科6属12种, 菹草(Potamogeton crispus L.)、篦齿眼子菜(Potamogeton pectinatus L.)及穗花狐尾藻(Myriophyllum spicatum L.)为主要优势种, 沉水植物优势群落主要有菹草群落、篦齿眼子菜群落、穗花狐尾藻群落、篦齿眼子菜+穗花狐尾藻群落。
2 研究方法 2.1 样地设置及群落调查于2018年8月按照均匀同时兼顾沉水植物空间分布格局的原则, 在南四湖共设10个代表性样地(图 1)。每一样地在0—1.0 m(L)、1.0—2.0 m(M)、2.0—3.0 m(H)3个水深区域内各设置10个样方, 每个样方间隔不少于30 m, 每一样方处用底部面积为0.2 m2的旋转式水下镰刀采集沉水植物1次, 每一样地共采集沉水植物样方30个, 全湖共采集沉水植物群落300个。将每一群落内所有植物用湖水洗净沥干水分后按种类称重, 同时记录每种的个体数目。针对每一样地同一水深范围内出现的所有物种, 每种取20株完整个体洗净后装入封口袋置于低温箱中带回实验室进行功能性状和植物体磷含量测定。

|
| 图 1 南四湖调查样地示意图 Fig. 1 Distribution of sampling plots in Nansi Lake |
选取9个表征沉水植物水深适应策略的功能性状进行测定:株高、茎分支数、茎节数、根长、茎直径、根围直径、比叶面积、生物量分配比和植物体磷含量。根性状是沉水植物响应底质异质性的直接性状, 表征底质营养的吸收能力;茎性状是沉水植物适应水位波动时的最佳响应性状, 决定植物冠层的高度和光获取能力;叶性状是表征植物营养收支或叶组织经济谱的主要指标;化学计量特征是衡量植物体内稳态大小的关键性指标;器官生物量分配比表征植株地上与地下部分资源分配状况, 体现植物适应水深胁迫的策略选择;这些性状与植物的光合作用、生长速率、营养吸收等密切相关[18, 25]。本研究中, 株高为根部至主茎顶部距离;茎分支数为主茎分支个数;茎节数为主茎上茎节间数目;茎直径为植物主茎上全部茎节间直径的平均值;根长为单株植物任意10条根长度的平均值;根围直径为根系垂直舒展状态下最宽处宽度;植物体磷含量为整株个体磷含量;生物量分配比为个体地上部分和地下部分生物量干重比值;比叶面积为单位叶片干重的叶面积。叶面积采用叶面积分析仪(CI-202)测定, 测定时分别在主茎上、中、下部各选取3片完整叶片进行测定。
2.3 数据计算与统计分析 2.3.1 群落加权功能性状计算沉水植物群落加权功能性状(Community Weighted Means, CWM)由物种功能性状值和物种相对丰富度加权平均得到[27], 其计算公式为:
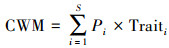
|
式中, S为群落物种数, Pi为物种i相对丰富度, Traiti为物种i某一功能性状特征值。
2.3.2 多样性指数选取与计算选用5种常用的物种多样性指数, 即Berger-Parker生态优势度指数(d)、Margalef丰富度指数(dMa)、Simpson多样性指数(λ)、Shannon-Wiener多样性指数(He′)、Pielou均匀度指数(Je)表征群落物种多样性。物种多样性指数采用基于Excel的宏程序计算[28]。
选用功能丰富度、功能均匀度和功能离散度3个维度的5种功能多样性指数来表征群落功能多样性。功能丰富度可量化群落功能生态位大小, 体现群落空间资源利用程度, 本文选用功能体积FRic指数来表征;功能均匀度指示群落功能性状的空间分布均匀程度, 本文选用多维功能均匀度指数FEve来表示;功能离散度指示群落功能性状值的离散状况, 表征群落种间生态位互补程度, 本文选用功能离散FDiv指数、功能分散FDis指数和二次熵RaoQ指数3个指数来表征[10, 13]。功能多样性指数计算采用R 3.5.1软件FD包。
2.3.3 统计分析单因素方差分析(One-way ANOVA)用于比较不同水深条件下群落加权功能性状值、物种多样性指数和功能多样性指数间的均值差异;分析之前, 对数据的正态性和方差齐性进行检验, 必要时进行对数、平方根转换;方差总体显著时, 采用Tukey HSD多重比较法进行均值间的多重比较。Pearson相关用于分析不同水深条件下物种多样性指数和功能多样性指数之间的关系。单因素方差分析和Pearson相关分析通过PASW Statistics 22.0实现, 图形制作由Origin 8.5完成。
3 结果与分析 3.1 群落物种组成及其分布格局本次调查共采集到沉水植物7种(不包括轮藻和菹草石芽), 其中篦齿眼子菜(P. pectinatus L.)、穗花狐尾藻(M. spicatum L.)、金鱼藻(Ceratophyllum demersum L.)在全湖分布范围最广;其次为光叶眼子菜(P. lucens L.)和竹叶眼子菜(P. wrightii Morong);轮叶黑藻(Hydrilla verticillata)和苦草(Vallisneria natans L.)仅在微山湖中部湖区和南部湖区(样地8和样地10)有分布(表 1)。样地10内物种最多, 7种沉水植物在该样地均有分布;其次为样地8和样地9(表 1)。样地内沉水植物群落多以斑块状分布, 篦齿眼子菜群落、穗花狐尾藻群落、篦齿眼子菜+穗花狐尾藻群落为主要优势群落(表 1)。下级湖区(样地6—样地10)沉水植物较上级湖区(样地1—样地5)丰富(表 1)。
| 样地 Sampling plots |
篦齿眼子菜 P. pectinatus |
穗花狐尾藻 M. spicatum |
光叶眼子菜 P. lucens |
竹叶眼子菜 P. wrightii |
金鱼藻 C.. demersum |
轮叶黑藻 H. verticillata |
苦草 V. natans |
| 1 | + | + | |||||
| 2 | + | + | + | ||||
| 3 | + | + | + | ||||
| 4 | + | + | + | + | |||
| 5 | + | + | + | ||||
| 6 | + | + | |||||
| 7 | + | + | + | ||||
| 8 | + | + | + | + | + | + | |
| 9 | + | + | + | + | + | ||
| 10 | + | + | + | + | + | + | + |
| +表示在该样地有分布 | |||||||
群落各加权性状对水深变化的响应迥异。3个水深梯度内群落茎分支数、茎节数和茎直径范围分别为1—20、1—111、0.42—2.61 mm, 3个水深梯度间各性状均无显著差异(P>0.05)(图 2)。M水深内群落株高和根长均值分别为130.57 cm和6.31 cm, 显著低于L(159.26 cm和9.80 cm)和H(154.41 cm和8.74cm)(P < 0.05), L和H间无显著差异(图 2)。M水深内群落生物量分配比(150.92)显著高于L(70.09)和H(65.87)(P < 0.05), L和H间无显著差异(P>0.05)(图 2)。根围直径和植物体磷含量表现为H(31.39 mm和5.32 mg/g)>L(25.71 mm和4.91 mg/g)>M(19.51 mm和4.59 mg/g)(P < 0.05)(图 2)。比叶面积在水深H(7.69 cm2/g)处最大, L(4.12 cm2/g)和M(4.66 cm2/g)间无显著差异(P>0.05)(图 2)。

|
| 图 2 不同水深沉水植物群落加权性状(平均值±标准误差) Fig. 2 The community weighted means traits of submerged vegetation community at different water depth (mean ± SE) L:0—1.0 m;M:1.0—2.0m;H:2.0—3.0 m; a b代表两两比较均值差异 |
不同水深沉水植物群落物种多样性对比结果表明, M水深内群落Margalef丰富度指数(0.69)和Shannon-Wiener多样性指数(0.64)显著高于L(0.51和0.35)和H(0.33和0.11)(P < 0.05), L和H间无显著差异(P>0.05)(图 3)。Berger-Parker生态优势度指数和Pielou均匀度指数表现为M(1.46和0.37)>L(1.25和0.26)>H(1.08和0.08)(P < 0.05)(图 3);而Simpson多样性指数却表现为M(0.64) < L(0.79) < H(0.93)(P < 0.05)(图 3)。

|
| 图 3 不同水深沉水植物群落物种多样性(平均值±标准误差) Fig. 3 Species diversity of submerged vegetation community at different water depth (mean ± SE) |
不同水深群落功能多样性对比结果表明, M水深内群落功能丰富度FRic指数(0.38)、功能均匀度FEve指数(0.39)、功能离散度FDis指数(1.46)和二次熵Rao指数(0.31)均显著高于L(0.24、0.11、0.70和0.13)和H(0.22、0.05、0.10和0.01)(P < 0.05), L和H间无显著差异(P>0.05)(图 4);而功能离散度FDiv指数在各水深梯度间无显著差异(P>0.05)(图 4)。

|
| 图 4 不同水深沉水植物群落功能多样性(平均值±标准误差) Fig. 4 Functional diversity of submerged vegetation community at different water depth (mean ± SE) FRic:功能丰富度指数, functional volume index;FEve:功能均匀度指数, functional evenness index;FDiv:功能离散度指数, functional division index;FDis:功能分散度指数, functional dispersion index;RaoQ:二次熵指数, functional division two-times entropy index |
由表 2可知, Berger-Parker生态优势度指数、Margalef丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数、Pielou均匀度指数5个物种多样性指数与功能分散度FDis指数、二次熵RaoQ指数2个功能多样性指数在各水深梯度内均显著相关;Simpson物种多样性指数与2个功能多样性指数呈负相关, 其余4个物种多样性指数与2个功能多样性指数为正相关;水深对5个物种多样性指数与2个功能多样性指数间的相关关系无显著影响。
| WD | d | dMa | λ | He′ | Je | |
| FRic | L | 0.530* | 0.474 | -0.634** | 0.624** | 0.609** |
| M | 0.432 | 0.572* | -0.426 | 0.491* | 0.426 | |
| H | 0.255 | 0.448 | -0.300 | 0.324 | 0.317 | |
| FEve | L | 0.828** | 0.964** | -0.804** | 0.853** | 0.848** |
| M | 0.083 | 0.234 | -0.109 | 0.146 | 0.103 | |
| H | 0.465** | 0.459** | -0.406 | 0.411 | 0.416 | |
| FDiv | L | -0.787** | -0.692** | 0.675** | -0.664** | -0.687** |
| M | -0.242 | -0.502* | 0.378 | -0.417 | -0.467 | |
| H | -0.195 | -0.023 | 0.153 | -0.139 | -0.140 | |
| FDis | L | 0.904** | 0.875** | -0.988** | 0.985** | 0.979** |
| M | 0.925** | 0.663** | -0.969** | 0.957** | 0.947** | |
| H | 0.903** | 0.625** | -0.959** | 0.961** | 0.961** | |
| RaoQ | L | 0.922** | 0.932** | -0.963** | 0.973** | 0.966** |
| M | 0.884** | 0.662** | -0.909** | 0.919** | 0.897** | |
| H | 0.79** | 0.588* | -0.876** | 0.833** | 0.833** | |
| *表示P < 0.05, **表示P < 0.01;WD:水深, Water depth;d:贝格-派克生态优势度指数, Berger-Parker dominance index;dMa:玛格列夫丰富度指数, Margalef richness index;λ:辛普森多样性指数, Simpson diversity index;He′:香农-威纳多样性指数, Shannon-Wiener diversity index;Je:皮洛均匀度指数, Pielou evenness index;FRic:功能丰富度指数, functional volume index;FEve:功能均匀度指数, functional evenness index;FDiv:功能离散度指数, functional division index;FDis:功能分散度指数, functional dispersion index;RaoQ:二次熵指数, functional division two-times entropy index;L:0—1.0 m;M:1.0—2.0m;H:2.0—3.0 m | ||||||
水深可显著改变5个物种多样性指数与功能丰富度FRic指数、功能均匀度FEve指数、功能离散度FDiv指数3个功能多样性指数间的相关关系。5个物种多样性指数与功能丰富度FRic指数(除Margalef丰富度指数与FRic指数外)、功能离散度FDiv指数2个功能多样性指数间关系强弱表现出L>H>M;5个物种多样性指数与功能均匀度FEve指数间关系表现为L>H>M(表 2)。因此, 水深可改变物种多样性与功能多样性间的关系, 但不同测度的物种多样性指数和功能多样性指数间关系对水深变化的响应各异。
4 讨论 4.1 群落加权功能性状对水深的响应水深是影响沉水植物群落构建的重要因素, 其变化可引起水体光照、温度、溶解氧等众多环境因子的关联变化。群落功能性状对水深的响应是多个环境因子共同作用的结果[22-23]。根据环境筛理论, 沉水植物经过水深的过滤作用, 具有相似性状的物种分布于特定水深范围内, 从而使群落性状在同一水深内趋同以及在不同水深间趋异[18, 26]。同一水深内的物种因具有相似的生境需求, 物种间的资源竞争作用增强, 又使得群落性状分异。因此, 群落性状由水深筛选作用和种间相互作用共同决定[21, 24]。本研究发现, 水深对沉水植物群落性状的影响并不是通过对每个功能性状的作用实现的, 而是作用于一系列的组合性状, 植物在此过程中通过性状权衡关系, 形成最优的水深适应策略。例如茎分支数、茎节数、茎直径等3个性状对水深梯度并无显著响应, 而株高、根长、根围直径、生物量分配比、比叶面积和植物体磷含量等6个性状的差异化响应使得每一水深范围内的植物群落形成了特定的性状组合。这可能有两个方面的原因:一是水深筛选作用和种间相互作用大致相抵, 使得综合驱动力对茎分支数、茎节数和茎直径等性状的影响相对较弱;二是这些性状对综合作用力的承受范围较宽, 并不是群落构建的敏感性状, 这些特定性状既不表现出趋同也不表现出趋异。
4.2 物种多样性对水深的响应研究发现, 水深可显著影响沉水植物群落物种多样性。中等水深(M)物种多样性最高, 该结论与中等程度的外界干扰利于群落维持较高的物种多样性的“中度干扰假说”一致[29]。在随水深关联变化的众多环境因子中, 光照可能是影响物种多样性形成的重要因子之一[23, 25]。光照在水体中随水深增加成指数式衰减, 深水区(H)光照不足使沉水植物群落生长受限;浅水区(L)较强的光照会产生光抑制, 同样不利于沉水植物群落的发展。从浅水区到深水区, 植物往往会通过增加株高、加大地上部分生物量分配等策略来适应水深引发的弱光胁迫。然而本研究中, 我们并没有发现这一规律, 反倒是在中等水深(M)植物群落表现出了欠发达的根部性状及较高的地上部分生物量投入, 结合群落生物量分配、根部性状的差异关系, 我们推测局部底质异质性可能是造成物种多样性差异的另一重要原因[22, 26]。因中等水深处底泥中丰富的植物残体, 使得植物群落对根部少许的投入就能满足根部的觅食行为, 这也为生物量分配和根部性状的差异找到了很好的解释。
4.3 功能多样性对水深的响应研究沉水植物群落功能多样性随水深梯度的变化规律, 其实质是探讨不同水深植物群落对生态位空间的占据情况以及功能性状在生态位空间的分布格局[11, 14, 25]。功能丰富度反映群落物种占据的生态位空间大小, 意味着植物群落占有的可利用资源的多少[10, 13]。本研究中, M水深处功能丰富度最高, 表明在中等水深条件下沉水植物群落利用了生境中的大量资源, 未被利用的剩余资源较少。功能均匀度表征群落性状在所占据的生态位空间的分布规律, 功能均匀度高意味着植物可充分地将其所占据的资源有效利用[10, 13]。本研究中, M水深处植物群落占据了大量的资源, 且利用率高;相反, L和H水深处植物群落即使占据了较小的资源空间, 但仍有部分资源未被完全利用。功能离散度反映群落总体性状的离散程度, 离散度高表明群落生态位分化程度高, 物种资源竞争弱[10, 13]。本研究中, M水深处FDis指数和二次熵RaoQ指数均高于L和H水深处, 表明中等水深处群落内种间资源竞争较弱。本研究中不同维度的功能多样性指数对水深的综合响应规律表明, 中等水深条件下, 植物群落占据了大量的资源, 群落总体性状高度离散并均匀分布于其所占据的资源空间中, 群落同时表现出较高的丰富度、均匀度以及离散度。
4.4 物种多样性和功能多样性关系对水深的响应物种组成和种间性状差异是造成群落功能多样性差异的重要因素[8-10]。因大多数功能多样性指数的计算需基于物种水平的参数, 因此功能多样性指数与物种多样性指数间存在着密切联系[1, 4]。前期研究表明, 功能多样性与物种多样性间的相关关系主要取决于群落所处的生境条件[2, 7, 15]。在浅水区(L), 功能多样性指数与物种多样性指数间(除FRic与dMa)均显著相关, 表明浅水区群落功能多样性主要依赖于群落物种组成而非种间性状差异, 功能冗余度低。随水深增加(M和H), 功能多样性指数与物种多样性指数间(除FRic与dMa)的相关关系减弱或不显著。其可能原因为:随水深增加, 沉水植物群落胁迫压力增加, 群落功能性状向着适应深水策略的方向趋同发展, 群落性状组成将被限制在深水范围, 种间性状差异变小, 物种多样性增加只会引起生态位进一步分化, 种间资源竞争加大, 功能多样性不再增加。
5 结论本文对南四湖不同水深下沉水植物群落加权性状、物种多样性、功能多样性进行了对比分析, 并对物种多样性与功能多样性关系随水深的响应进行了研究, 结果表明:
(1) 水深可显著影响株高、根长、根围直径、生物量分配比、比叶面积和植物体磷含量6个群落性状;茎分支数、茎节数和茎直径3个群落性状对水深变化无显著响应。水深对沉水植物群落性状的影响并不是通过对每个功能性状的作用而实现的, 而是作用于一系列的组合性状, 植物在此过程中通过性状权衡关系, 形成最优的水深适应策略。
(2) 水深可显著影响沉水植物群落物种多样性和功能多样性, 中等水深(M)处沉水植物群落具有较高的物种多样性指数和功能多样性指数, 浅水(L)处和深水(H)处物种多样性和功能多样性差别不大。
(3) 不同测度的物种多样性指数和功能多样性指数间相关关系对水深变化的响应各异, 水深可显著改变5个物种多样性指数与功能丰富度FRic指数、功能均匀度FEve指数、功能离散度FDiv指数3个功能多样性指数间的相关关系, 但对5个物种多样性指数与功能离散度FDis指数、二次熵Rao指数2个功能多样性指数间的相关关系无显著影响。
综上, 沉水植物群落加权性状、物种多样性、功能多样性及其相互关系对水深变化响应迥异。因此, 在阐释水深对沉水植物群落构建影响机制时要综合考虑多个度量维度。
| [1] |
董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472-1483. |
| [2] |
朱云云, 王孝安, 王贤, 邓美皎. 坡向因子对黄土高原草地群落功能多样性的影响. 生态学报, 2016, 36(21): 6823-6833. |
| [3] |
Ackerly D D, Cornwell W K. A trait-based approach to community assembly:partitioning of species trait values into within-and among-community components. Ecology Letters, 2007, 10(2): 135-145. |
| [4] |
陈静, 蒋万祥, 贺诗水, 王洪凯, 卓丽玲, 陈青, 王红妹. 新薛河底栖动物物种多样性与功能多样性研究. 生态学报, 2018, 38(9): 3328-3336. |
| [5] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学:生命科学, 2015, 45(4): 325-339. |
| [6] |
Mokany K, Ash J, Roxburgh S. Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. Journal of Ecology, 2008, 96(5): 884-893. DOI:10.1111/j.1365-2745.2008.01395.x |
| [7] |
张雪妮, 李岩, 何学敏, 杨晓东, 吕光辉. 荒漠植物功能性状及其多样性对土壤水盐变化的响应. 生态学报, 2019, 39(5): 1541-1550. |
| [8] |
薛倩妮, 闫明, 毕润成. 山西五鹿山森林群落木本植物功能多样性. 生态学报, 2015, 35(21): 7023-7032. |
| [9] |
Cadotte M W, Carscadden K, Mirotchnick N. Beyond species:functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 2011, 48(5): 1079-1087. DOI:10.1111/j.1365-2664.2011.02048.x |
| [10] |
Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence:the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. DOI:10.1111/j.0030-1299.2005.13886.x |
| [11] |
Mouchet M A, Villéger S, Mason N W H, Mouillot D. Functional diversity measures:an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 2010, 24(4): 867-876. |
| [12] |
江小雷, 张卫国. 功能多样性及其研究方法. 生态学报, 2010, 30(10): 2766-2773. |
| [13] |
Petchey O L, Gaston K J. Functional Diversity (FD), species richness and community composition. Ecology Letters, 2002, 5(3): 402-411. DOI:10.1046/j.1461-0248.2002.00339.x |
| [14] |
柴永福, 岳明. 植物群落构建机制研究进展. 生态学报, 2016, 36(15): 4557-4572. |
| [15] |
龚时慧, 温仲明, 施宇. 延河流域植物群落功能性状对环境梯度的响应. 生态学报, 2011, 31(20): 6088-6097. |
| [16] |
Sasaki T, Okubo S, Okayasu T, Jamsran U, Ohkuro T, Takeuchi K. Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands. Ecology, 2009, 90(9): 2598-2608. DOI:10.1890/08-1850.1 |
| [17] |
Bu W S, Zang R G, Ding Y. Functional diversity increases with species diversity along successional gradient in a secondary tropical lowland rainforest. Tropical Ecology, 2014, 55(3): 393-401. |
| [18] |
Fu H, Zhong J Y, Yuan G X, Ni L Y, Xie P, Cao T. Functional traits composition predict macrophytes community productivity along a water depth gradient in a freshwater lake. Ecology and Evolution, 2014, 4(9): 1516-1523. DOI:10.1002/ece3.1022 |
| [19] |
帅方敏, 李新辉, 陈方灿, 李跃飞, 杨计平, 李捷, 武智. 淡水鱼类功能多样性及其研究方法. 生态学报, 2017, 37(15): 5228-5237. |
| [20] |
蒋万祥, 陈静, 王红妹, 贺诗水, 卓丽玲, 陈青, 王洪凯, 蔡庆华. 新薛河典型生境底栖动物功能性状及其多样性. 生态学报, 2018, 38(6): 2007-2016. |
| [21] |
刘洋, 付文龙, 操瑜, 李伟. 沉水植物功能性状研究的思考. 植物科学学报, 2017, 35(3): 444-451. |
| [22] |
王华, 逄勇, 刘申宝, 马璇. 沉水植物生长影响因子研究进展. 生态学报, 2008, 28(8): 3958-3968. |
| [23] |
吴明丽, 李叙勇. 光衰减及其相关环境因子对沉水植物生长影响研究进展. 生态学报, 2012, 32(22): 7202-7212. |
| [24] |
符辉, 钟家有, 袁桂香, 曹特, 倪乐意. 沉水植物功能性状变异的来源与结构——以微齿眼子菜(Potamogeton maackianus)为例. 湖泊科学, 2015, 27(3): 429-435. |
| [25] |
Fu H, Zhong J Y, Fang S W, Hu J M, Guo C J, Lou Q, Yuan G X, Dai T T, Li Z Q, Zhang M, Li W, Xu J, Cao T. Scale-dependent changes in the functional diversity of macrophytes in subtropical freshwater lakes in south China. Scientific Reports, 2017, 7(1): 8294. DOI:10.1038/s41598-017-08844-8 |
| [26] |
翟水晶, 胡维平, 邓建才, 陈桥, 韩红娟, 刘伟龙. 不同水深和底质对太湖马来眼子菜(Potamogeton malaianus)生长的影响. 生态学报, 2008, 28(7): 3035-3041. |
| [27] |
Lavorel S, Grigulis K, McIntyre S, Williams N S G, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A. Assessing functional diversity in the field-methodology matters!. Functional Ecology, 2008, 22(1): 134-147. |
| [28] |
孔凡洲, 于仁成, 徐子钧, 周名江. 应用Excel软件计算生物多样性指数. 海洋科学, 2012, 36(4): 57-62. |
| [29] |
Tilman D, Wedin D, Knops J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature, 1996, 379(6567): 718-720. DOI:10.1038/379718a0 |