 2020, Vol. 40
2020, Vol. 40文章信息
- 刘敏, 张潇月, 李晓丽, 刘琦, 孟万忠, 徐满厚
- LIU Min, ZHANG Xiaoyue, LI Xiaoli, LIU Qi, MENG Wanzhong, XU Manhou
- 黄土高原林下草地对模拟增温的短期响应
- Short-term responses of under-canopy grasslands to the experimental warming on the Loess Plateau, China
- 生态学报. 2020, 40(17): 6009-6024
- Acta Ecologica Sinica. 2020, 40(17): 6009-6024
- http://dx.doi.org/10.5846/stxb201810222285
-
文章历史
- 收稿日期: 2018-10-22
- 网络出版日期: 2020-07-10






2. 太原师范学院汾河流域科学发展研究中心, 晋中 030619;
3. 山西旅游职业学院旅游文化艺术系, 太原 030031
2. Research Center for Scientific Development in Fenhe River Valley, Taiyuan Normal University, Jinzhong 030619, China;
3. Department of Tourism Culture and Art, Shanxi Vocational College Tourism, Taiyuan 030031, China
山地林下草地作为一种典型的生态系统复合体, 是现代生态学研究的重要内容之一, 然而对林下草地至今没有一个明确概念[1]。有人提出林下草地是乔木和灌木郁闭度分别为0.3和0.4以下的草地, 也有人认为林下草地是森林植被景观破碎化后形成的次生草本群落, 与森林群落共同构成林草复合生态景观[2]。林下草地作为森林生态系统的重要组成部分, 其物种多样性可提高土壤肥力、促进土壤养分循环和维护森林立地质量, 对提高森林水土保持能力、评估林间植物多样性有显著作用[3-6]。因此, 对林下草地的研究多是关注其在不同林地的物种多样性, 如岩溶区林地[7]、防护林林地[8]、橡胶林[9]、油松林[2]、刺槐林[3]等。
山地海拔、地形的变化直接导致热量、水分等环境因子发生改变, 进而影响山地植被的物种组成和群落结构, 这些环境因子沿海拔梯度的变化要比纬度梯度快1000倍[10-13]。山地空气温度随海拔升高而降低成为普遍事实, 而由温室气体增加引起的气候变暖也会影响山地温度的垂直变化[14]。2000—2015年全球变暖速率约为20世纪后半叶的一半[15], 未来40年全球变暖加速的可能性增大[16-17], 升温更为明显[18]。高海拔地区是响应全球变暖最敏感的区域之一, 其升温速率及幅度均远大于低海拔地区[19-20]。气候变暖势必对山地植被产生重要影响, 而山地林下草地对气候变暖的响应也更为迅速。
黄土高原作为中国乃至世界上黄土分布面积最大的地区, 降水稀少、蒸发强烈, 植被盖度低, 水土流失严重, 是典型的生态环境恶劣地区。前期已对黄土高原东部山地植被群落多样性的空间格局进行了相关研究[12, 21], 发现黄土高原东部山地具有明显的纬度梯度和海拔梯度, 这两大地理因素通过改变植物生长所需的水热环境因子, 对山地林下草本植物的群落结构产生显著影响。但是, 这些研究大多是采用调查样地法对山地植被的空间分布及其原因进行探讨, 较少通过试验样地法对该区植被响应气候变暖的特征作深入研究, 而对该区植被响应气候变暖的空间格局进行研究更是少之又少。前期已在黄土高原东部山地布设了模拟气候变暖的野外增温试验样地, 初步研究了模拟增温对吕梁山草本植物群落物种多样性的影响[22]。该增温实验于2016年实施, 只获得了增温1年的实验数据, 反映的是短期增温效应;而且, 研究区只选取了位于吕梁山较高纬度的管涔山和较低纬度的五鹿山, 并未将位于吕梁山中等纬度的关帝山纳入研究中。因此, 为了深入探究黄土高原林下草地对气候变暖的短期响应及其随纬度、海拔的变化格局, 本研究在黄土高原东部选择吕梁山北段的管涔山、中段的关帝山和南段的五鹿山, 分别设置不同幅度的模拟增温样地, 开展三个方面的研究:(1)林下草地水热因子对短期增温的响应;(2)林下草地植物生长特征对短期增温的响应;(3)林下草地植物生长特征与水热因子关系对短期增温的响应。在不同地理梯度研究山地林下草地对气候变暖的短期响应, 有助于理解全球变暖下山地森林植被的响应机制。
1 研究方法 1.1 研究区概况吕梁山系位于黄土高原东部, 南北延长约400 km, 纬度跨度近3°, 从北向南主要包括管涔山、关帝山和五鹿山[12]。管涔山位于忻州市宁武县东寨镇(38°57′—39°03′ N、112°36′—112°37′ E), 年均气温7℃, 年均降水量475 mm, 无霜期105 d;土壤类型主要是山地褐土、棕壤, 植被类型主要有针阔叶混交林、针叶林、亚高山草甸。关帝山位于吕梁市交城县庞泉沟镇(37°20′—38°20′ N、110°18′—111°18′ E), 年均气温4℃, 年均降水量831 mm, 无霜期115 d;土壤类型从低海拔到高海拔依次为淡褐土、淋溶褐土、棕壤和亚高山草甸土, 分布有阔叶林、针阔叶混交林、针叶林、亚高山草甸等植被类型。五鹿山位于临汾市蒲县和隰县交界处(36°23′—36°38′ N、111°2′—111°18′ E), 年均气温11℃, 年均降水量490 mm, 无霜期165 d;土壤类型有棕壤、褐土、草甸土、山地草甸土等, 植被类型较为单一, 地带性植被为落叶阔叶林。
1.2 试验样地设置于2016年9月进行试验样地设置。根据管涔山、关帝山和五鹿山的实际高度, 分别划分高、中、低3种海拔梯度, 每一海拔梯度均用开顶式生长室(Open-top chamber, OTC)设置高、低2种幅度的模拟增温样地。其中, 管涔山的高、中、低平均海拔依次为2571.3、2395.0、2222.4 m, 关帝山为2179.1、2000.7、1899.7 m, 五鹿山为1602.0、1458.6、1318.0 m。
在每一海拔梯度, 选取植被分布均匀、物种较多的林间草本群落进行增温样地布设, 样地面积为35—40 m2。采用随机区组设计, 设置3个区组, 每一区组包括对照(Control, CK)、低度增温(Low temperature increase, LW)、高度增温(High temperature increase, HW)3种实验处理, 共布设9个增温样方。同一区组的3个增温样方随机排列, 各增温样方间距为1—3 m。在整个吕梁山划分出的9个海拔梯度, 共设置81个增温样方。
增温样地采用OTC进行模拟增温处理, 箱体为半封闭的正六边形, 上下两端开口, 各斜边与地面夹角均为60°, 由透光率可达90%的透明PC板材料建构(图 1)。其中, CK样方无OTC, 不进行增温, 保持植被自然状态, 取样面积为1 m×1 m;LW样方OTC高度为40 cm, 顶面边长25 cm, 底面边长50 cm;HW样方OTC高度为80 cm, 顶面边长25 cm, 底面边长75 cm。

|
| 图 1 模拟增温试验样地和样方框 Fig. 1 Simulated warming plots and sampling frame |
在不同山地选取合适的林下草地作为试验对象。其中, 五鹿山选取的林下草地位于落叶阔叶林带, 以辽东栎(Quercus liaotungensis)为建群种;关帝山选取的林下草地位于针阔叶混交林带, 阔叶林以白桦(Betula platyphylla)为建群种, 针叶林以华北落叶松(Larix principis-rupprechtii)为建群种;管涔山选取的林下草地位于纯针叶林带, 以云杉(Picea asperata)为建群种。因此, 由于纬度、海拔差异, 吕梁山不同山地形成不同的森林林带结构, 造成在不同山地所选取的林下草地也具有一定差异, 需要对林下草地及其所在林带同时进行调查。于2017年8月(已增温1a)植被生长季对林间草本群落的响应特征进行调查, 同时调查所在林带的乔木层生长特征。增温样方内草本植物的调查指标包括高度、盖度、密度、频度;增温样地外乔木植物的调查指标包括高度、盖度、密度, 所设调查面积为25 m×25 m。采用大小为0.5 m×0.5 m的样方框测量草本群落特征值, 测量时将其分成25个0.1 m×0.1 m的网格, 按物种进行统计(图 1)。其中, 高度为各物种自然状态下的最高值, 密度为各物种单位面积内的个体数, 盖度为各物种在网格中所占的面积, 频度为各物种在网格中出现的次数。在五鹿山试验样地共统计到林下草本植物30个物种, 在关帝山统计到37个物种, 在管涔山统计到28个物种, 其生长指标如附表所示。关帝山林下草本植物的物种数高于相邻两个山地, 这也是将关帝山纳入本研究区域的一个重要原因。
1.4 环境因子调查由于3个山地相隔距离太远(最远可达约300 km), 只能保证同一山地不同海拔梯度的环境因子同步测量, 不同山地环境因子的测量则相隔1—2 d。同时由于尚未建成定位观测试验站, 无专门人员对增温样方的实验仪器进行看护, 故环境因子的测量只能在白天进行。测量增温样方中空气-土壤水热环境因子, 所使用仪器均来自美国Onset计算机公司生产的HOBO系列产品。空气温度和空气湿度由架设在距地面20 cm高度处的温湿度传感器(S-THB-M008)测定;土壤温度用温度传感器(S-TMB-M006)分5个深度进行测定:10、20、30、40、50 cm。所有传感器均连接到数据记录仪(H21-USB), 以30 s为时间间隔存储数据, 取1 min内的数据平均值进行分析。土壤水分采用烘干法进行分层测定:用内径为3 cm的土钻钻取5层土样(0—10、10—20、20—30、30—40、40—50 cm), 将土样用铝盒密封, 放进105℃的烘箱中烘干至恒重, 称量后计算土壤水分。
1.5 数据统计分析(1) 土壤水分计算
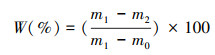
|
式中, W为所测样品的土壤水分/%, m0为空铝盒质量/g, m1为烘干前铝盒和土样的质量/g, m2为烘干后铝盒和土样的质量/g。
(2) 统计分析
在不同时空尺度上, 用OriginPro 9.1软件分析不同处理下(CK、LW、HW)吕梁山不同海拔空气温湿度随时间的变化趋势、土壤温湿度随深度的变化趋势及不同处理下吕梁山植被群落特征随海拔的变化趋势;同时采用IBM SPSS Statistics 22.0软件中的GLM(General linear model)模型, 对吕梁山温度、水分、植被在不同山地、土层、海拔、处理下的差异进行多因素方差分析。其中, 各指标在不同处理下的差异分析是指LW(LW—CK)和HW(HW—CK)分别与CK的比较, 得到低度增温和高度增温下的变化幅度。
对于吕梁山不同处理下温度、水分、植被的关系, 首先利用Canoco for Windows 4.5软件对植被群落特征数据进行DCA(Detrended correspondence analysis)分析, 确定用线性模型进行直接梯度分析(4个排序轴的最大梯度长度小于3), 再选择其中的RDA(Redundancy analysis)模型进行约束性排序分析。在进行DCA分析时, 整理每个处理下的植被群落特征数据(密度、频度、盖度、高度), 由于每个处理包括27个样方(3个山地×3个海拔/山地×3个重复/海拔), 故以样方为单位对27组植被群落特征数据进行处理;之后, 选择合适的RDA对这27组植被群落特征数据和相应样方内的环境因子数据(20 cm高度空气温度、20 cm高度空气湿度、0—50 cm深度土壤温度、0—50 cm深度土壤水分)进行排序分析, 其中由传感器以时间为单位测定的环境因子数据在进行RDA分析之前需要以样方为单位取平均值作为处理数据(环境因子也为27组数据), 从而得到不同处理下植被群落特征与水热环境因子的排序关系。
2 结果与分析 2.1 山地不同海拔空气温湿度对增温的响应LW和HW处理下空气温度在五鹿山分别增加0.56℃和1.48℃, 在关帝山分别增加0.44℃和0.88℃, 在管涔山分别增加0.40℃和0.65℃(图 2)。空气温度在增温处理下出现极显著差异(P < 0.001), 不同海拔的空气温度在增温处理下出现显著差异(P=0.012), 不同山地不同海拔的空气温度在增温处理下也出现极显著差异(P < 0.001)(表 1)。因此, 不同增温处理下吕梁山空气温度显著增加, 增幅在LW和HW处理下分别达到0.47℃和1.00℃;且增幅随纬度增加呈减小趋势(P=0.092), 随海拔增加显著增大(P=0.012)。

|
| 图 2 不同增温处理下山地不同海拔空气温度随时间的变化 Fig. 2 Variations of air temperature with time in different warming treatments at different altitudes CK:对照, Control;LW:低度增温, Low temperature increase;HW:高度增温, High temperature increase |
| 项目 Item | 土壤水分 Soil moisture | 土壤温度 Soil temperature | 空气温度 Air temperature | 空气湿度 Air humidity | 植物密度 Vegetation density | 植物频度 Vegetation frequency | 植物盖度 Vegetation coverage | 植物高度 Vegetation height |
| 山地Mountain | 0.000 | 0.000 | 0.000 | 0.000 | 0.070 | 0.137 | 0.013 | 0.000 |
| 海拔Altitude | 0.000 | 0.000 | 0.000 | 0.042 | 0.006 | 0.002 | 0.001 | 0.129 |
| 处理Treatment | 0.182 | 0.055 | 0.000 | 0.610 | 0.077 | 0.473 | 0.042 | 0.066 |
| 山地×海拔 Mountain×Altitude | 0.000 | 0.000 | 0.000 | 0.031 | 0.020 | 0.065 | 0.032 | 0.000 |
| 山地×处理 Mountain×Treatment | 0.094 | 0.003 | 0.092 | 0.766 | 0.201 | 0.747 | 0.252 | 0.518 |
| 海拔×处理 Altitude×Treatment | 0.209 | 0.347 | 0.012 | 0.693 | 0.366 | 0.507 | 0.405 | 0.896 |
| 山地×海拔×处理 Mountain×Altitude×Treatment | 0.590 | 0.000 | 0.000 | 0.212 | 0.912 | 0.953 | 0.525 | 0.283 |
| 土层Layer | 0.000 | 0.000 | — | — | — | — | — | — |
| 山地×土层 Mountain×Layer | 0.000 | 0.000 | — | — | — | — | — | — |
| 海拔×土层 Altitude×Layer | 0.000 | 0.000 | — | — | — | — | — | — |
| 处理×土层 Treatment×Layer | 0.946 | 0.647 | — | — | — | — | — | — |
| 山地×海拔×土层 Mountain×Altitude×Layer | 0.000 | 0.000 | — | — | — | — | — | — |
| 山地×处理×土层 Mountain×Treatment×Layer | 0.953 | 0.545 | — | — | — | — | — | — |
| 海拔×处理×土层 Altitude×Treatment×Layer | 0.592 | 0.925 | — | — | — | — | — | — |
| 山地×海拔×处理×土层 Mountain×Altitude× Treatment×Layer | 0.998 | 0.810 | — | — | — | — | — | — |
| ×:交互作用;—:无此项 | ||||||||
LW和HW处理下空气湿度在五鹿山分别增加0.87%和减小0.40%, 在关帝山分别增加3.10%和5.29%, 在管涔山分别增加0.13%和0.94%(图 3)。空气湿度在山地(P < 0.001)和海拔(P=0.042)间达到显著差异, 但在增温处理下差异并不显著(P=0.61), 说明空气湿度对增温响应不敏感(表 1)。因此, 不同增温处理下吕梁山空气湿度呈增加趋势, 增幅在LW和HW处理下分别为1.37%和1.94%;且增幅随纬度(P=0.766)和海拔(P=0.693)增加均呈先增大后减小趋势。

|
| 图 3 不同增温处理下山地不同海拔空气湿度随时间的变化 Fig. 3 Variations of air humidity with time in different warming treatments at different altitudes |
LW和HW处理下土壤温度在五鹿山分别增加0.08℃和0.38℃, 增幅随深度增加略呈减小趋势;在关帝山分别减小0.22℃和0.19℃, 增幅随深度增加略呈增大趋势;在管涔山分别减小0.08℃和无变化, 增幅随深度增加略呈减小趋势(图 4)。土壤温度在山地、海拔、土层间的差异均达到极显著水平(P < 0.001), 且不同山地的土壤温度(P=0.003)和不同山地不同海拔的土壤温度(P < 0.001)在增温处理下差异也均极显著(表 1)。因此, 不同增温处理下吕梁山土壤温度波动不一致(P=0.055), 在LW处理下减小0.07℃, 在HW处理下增加0.06℃;且增幅随纬度增加显著减小(P=0.003), 随海拔增加呈减小趋势(P=0.347)。

|
| 图 4 不同增温处理下山地不同海拔土壤温湿度随深度的变化 Fig. 4 Variations of soil temperature and moisture with depths in different warming treatments at different altitudes |
LW和HW处理下土壤水分在五鹿山分别增加0.31%和减小0.94%, 增幅随土层加深略呈减小趋势;在关帝山分别减小0.72%和1.28%, 增幅随土层加深略呈增大趋势;在管涔山分别减小0.54%和增加0.31%, 增幅随土层加深呈减小趋势(图 4)。土壤水分在山地、海拔、土层间的差异均达到极显著水平(P < 0.001), 但在增温处理下差异均不显著(P>0.05), 说明土壤水分对增温响应不敏感(表 1)。因此, 不同增温处理下吕梁山土壤水分呈减小趋势(P=0.182), 减幅在LW和HW处理下分别为0.32%和0.64%;且增幅随纬度增加分别呈减小和增大趋势(P=0.094), 随海拔增加均呈增大趋势(P=0.209)。
2.3 山地不同海拔植被群落特征对增温的响应除五鹿山低海拔处密度最大的植物为铁杆蒿(Artemisia gmelinii)外, 其它样地密度最大的植物均为披针苔草(Carex lanceolata), 表明吕梁山林下草本群落的优势种为披针苔草(表 2)。该苔草在五鹿山平均密度为291.11株/m2, 在关帝山平均为454.37株/m2, 在管涔山平均为711.26株/m2, 即随纬度增加, 优势种密度增加。
| 山地 Mountain | 位置 Location | 优势种 Dominant species | 密度 Density/ (株/m2) |
| 五鹿山 | 低海拔 | 铁杆蒿 | 128.89 |
| 中海拔 | 披针苔草 | 318.22 | |
| 高海拔 | 披针苔草 | 515.11 | |
| 关帝山 | 低海拔 | 披针苔草 | 602.22 |
| 中海拔 | 披针苔草 | 412.00 | |
| 高海拔 | 披针苔草 | 348.89 | |
| 管涔山 | 低海拔 | 披针苔草 | 651.56 |
| 中海拔 | 披针苔草 | 1152.00 | |
| 高海拔 | 披针苔草 | 330.22 |
LW和HW处理下植物密度在五鹿山分别增加123.94株/m2和72.94株/m2, 在关帝山分别减少6.05株/m2和增加18.25株/m2, 在管涔山分别增加5.92株/m2和144.40株/m2(图 5)。植物密度在海拔(P=0.006)间和不同山地的海拔(P=0.02)间差异达到显著水平, 但在增温处理下均未出现显著性差异(P>0.05), 说明植物密度对增温响应不敏感(表 1)。因此, 不同增温处理下吕梁山植物密度呈增加趋势(P=0.077), 增幅在LW和HW处理下分别为41.27株/m2和78.53株/m2;且增幅随纬度(P=0.201)增加在LW处理下呈减小趋势, 在HW处理下呈增大趋势, 随海拔(P=0.366)增加均呈增大趋势。

|
| 图 5 不同增温处理下山地植被群落特征随海拔的变化 Fig. 5 Variations of plant community properties with altitudes in different warming treatments |
LW和HW处理下植物频度在五鹿山分别增加15.91%和6.85%, 在关帝山分别减小1.09%和增加0.38%, 在管涔山分别增加1.60%和3.19%(图 5)。植物频度只在海拔间差异达到显著水平(P=0.002), 在增温处理下均未出现显著性差异(P>0.05), 说明植物频度对增温响应不敏感(表 1)。因此, 不同增温处理下吕梁山植物频度呈增加趋势(P=0.473), 增幅在LW和HW处理下分别为5.47%和3.47%;且增幅随纬度增加呈减小趋势(P=0.747), 随海拔增加呈增大趋势(P=0.507)。
LW和HW处理下植物盖度在五鹿山分别增加13.19%和3.19%, 在关帝山分别增加0.94%和0.95%, 在管涔山分别增加1.83%和减小1.51%(图 5)。植物盖度在山地(P=0.013)、海拔(P=0.001)、处理(P=0.042)间差异均达到显著水平, 但不同山地(P=0.252)、不同海拔(P=0.405)和不同山地不同海拔(P=525)的植物盖度在增温处理下未达到显著差异(表 1)。因此, 不同增温处理下吕梁山植物盖度显著增加, 增幅在LW和HW处理下分别达到5.32%和0.88%;且增幅随纬度增加呈减小趋势, 随海拔增加呈增大趋势。
LW和HW处理下植物高度在五鹿山分别增加0.06 m和未发生变化, 在关帝山分别增加0.05 m和0.06 m, 在管涔山均增加0.02 m(图 5)。植物高度在山地(P < 0.001)间和不同山地的海拔(P < 0.001)间差异达到极显著水平, 但在增温处理下均未达到显著性差异(P>0.05), 说明植物高度对增温响应不敏感(表 1)。因此, 不同增温处理下吕梁山植物高度呈增加趋势(P=0.066), 增幅在LW和HW处理下分别为0.04 m和0.03 m;且增幅随纬度(P=0.518)增加在LW处理下呈减小趋势, 在HW处理下呈增大趋势, 随海拔(P=0.896)增加均呈增大趋势。
2.4 山地不同增温处理下温度、水分、植被的关系植物密度在CK处理下与12个温度、水分因子的相关性分析中显著水平占50%, 其中极显著水平只有1个, 与土壤温度呈显著负相关;在LW处理下的显著水平占92%, 其中极显著水平有5个, 与空气-土壤温度呈显著负相关, 与土壤水分呈显著正相关;在HW处理下的显著水平占100%, 其中极显著水平有11个, 与温度因子呈极显著负相关, 与水分因子呈极显著正相关(表 3和图 6)。相关系数绝对值在增温处理下呈递增趋势的因子有11个, 占总因子数的92%。
| AT | AH | ST1 | ST2 | ST3 | ST4 | ST5 | SM1 | SM2 | SM3 | SM4 | SM5 | ||
| 密度Density | CK | -0.229 | 0.223 | -0.433 | -0.330 | -0.435 | -0.439 | -0.439 | 0.525 | 0.420 | 0.336 | 0.165 | 0.062 |
| LW | -0.499 | 0.370 | -0.515 | -0.506 | -0.496 | -0.485 | -0.480 | 0.484 | 0.459 | 0.505 | 0.452 | 0.483 | |
| HW | -0.804 | 0.397 | -0.797 | -0.789 | -0.785 | -0.786 | -0.788 | 0.734 | 0.856 | 0.808 | 0.722 | 0.681 | |
| 频度Frequency | CK | -0.151 | 0.133 | -0.270 | -0.239 | -0.229 | -0.213 | -0.193 | 0.421 | 0.336 | 0.236 | 0.062 | 0.000 |
| LW | -0.330 | 0.208 | -0.308 | -0.267 | -0.247 | -0.228 | -0.215 | 0.515 | 0.445 | 0.345 | 0.216 | 0.041 | |
| HW | -0.558 | 0.295 | -0.561 | -0.551 | -0.541 | -0.535 | -0.524 | 0.589 | 0.595 | 0.505 | 0.303 | 0.368 | |
| 盖度Coverage | CK | 0.142 | -0.149 | 0.004 | 0.047 | 0.070 | 0.090 | 0.109 | 0.126 | 0.111 | 0.054 | -0.025 | -0.075 |
| LW | 0.103 | -0.151 | 0.169 | 0.208 | 0.225 | 0.239 | 0.248 | 0.174 | 0.053 | -0.030 | -0.175 | -0.260 | |
| HW | 0.156 | 0.087 | 0.193 | 0.215 | 0.223 | 0.226 | 0.241 | 0.118 | -0.021 | -0.143 | -0.363 | -0.265 | |
| 高度Height | CK | 0.534 | -0.252 | 0.541 | 0.521 | 0.568 | 0.572 | 0.583 | -0.278 | -0.473 | -0.509 | -0.558 | -0.607 |
| LW | 0.601 | -0.365 | 0.668 | 0.674 | 0.672 | 0.668 | 0.671 | -0.303 | -0.559 | -0.678 | -0.617 | -0.636 | |
| HW | 0.370 | -0.027 | 0.386 | 0.389 | 0.388 | 0.387 | 0.398 | -0.076 | -0.332 | -0.456 | -0.675 | -0.587 | |
| AT:空气温度, Air temperature;AH:空气湿度, Air humidity;ST1:10 cm深度土壤温度, Soil temperature at 10 cm depth;ST2:20 cm深度土壤温度, Soil temperature at 20 cm depth;ST3:30 cm深度土壤温度, Soil temperature at 30 cm depth;ST4:40 cm深度土壤温度, Soil temperature at 40 cm depth;ST5:50 cm深度土壤温度, Soil temperature at 50 cm depth;SM1:0—10 cm深度土壤水分, Soil moisture at 0—10 cm depth;SM2:10—20 cm深度土壤水分, Soil moisture at 10—20 cm depth;SM3:20—30 cm深度土壤水分, Soil moisture at 20—30 cm depth;SM4:30—40 cm深度土壤水分, Soil moisture at 30—40 cm depth;SM5:40—50 cm深度土壤水分, Soil moisture at 40—50 cm depth;CK:对照Control; LW:低度增温Low temperature increase; HW:高度增温High temperature increase | |||||||||||||

|
| 图 6 不同增温处理下山地温度、水分、植被的关系 Fig. 6 Correlations among temperature, moisture and vegetation in different warming treatments AT:空气温度, Air temperature;AH:空气湿度, Air humidity;ST1:10 cm深度土壤温度, Soil temperature at 10 cm depth;ST2:20 cm深度土壤温度, Soil temperature at 20 cm depth;ST3:30 cm深度土壤温度, Soil temperature at 30 cm depth;ST4:40 cm深度土壤温度, Soil temperature at 40 cm depth;ST5:50 cm深度土壤温度, Soil temperature at 50 cm depth;SM1:0—10 cm深度土壤水分, Soil moisture at 0—10 cm depth;SM2:10—20 cm深度土壤水分, Soil moisture at 10—20 cm depth;SM3:20—30 cm深度土壤水分, Soil moisture at 20—30 cm depth;SM4:30—40 cm深度土壤水分, Soil moisture at 30—40 cm depth;SM5:40—50 cm深度土壤水分, Soil moisture at 40—50 cm depth |
植物频度在CK处理下的显著水平只有1个, 占总因子数的8%, 与0—10 cm土壤水分呈显著正相关;在LW处理下的显著水平占17%, 其中极显著水平有1个, 与0—20 cm土壤水分呈显著正相关;在HW处理下的显著水平占75%, 且均为极显著水平, 与空气-土壤温度呈极显著负相关, 与0—30 cm土壤水分呈极显著正相关(表 3和图 6)。相关系数绝对值在增温处理下呈递增趋势的因子有12个, 占总因子数的100%。
植物盖度在不同增温处理下的相关性分析均未达到显著水平(表 3和图 6), 相关系数绝对值在增温处理下呈递增趋势的因子有4个, 占总因子数的33%。
植物高度在CK处理下的显著水平占83%, 其中极显著水平有9个, 与空气-土壤温度呈极显著正相关, 与10—50 cm土壤水分呈显著负相关;在LW处理下的显著水平占83%, 且均为极显著水平, 与空气-土壤温度呈极显著正相关, 与10—50 cm土壤水分呈极显著负相关;在HW处理下的显著水平占67%, 其中极显著水平有2个, 与土壤温度呈显著正相关, 与20—50 cm土壤水分呈显著负相关(表 3和图 6)。相关系数绝对值在增温处理下呈递增趋势的因子只有1个, 占总因子数的8%。
总体而言, 不同增温处理下植被与温度、水分因子的关系趋于增强, RDA各排序轴的特征值总和在LW和HW处理下分别增加6.74%和33.71%, 各排序轴关系总和在LW和HW处理下分别增加1.30%和3.55%(表 4)。在LW和HW处理下, 植被与温度因子关系的相关系数绝对值分别增加31.49%和56.82%, 与水分因子关系的相关系数绝对值分别增加38.67%和62.89%;后者的增幅略大于前者, 表明植被与水分的关系对增温响应更为敏感(图 6)。
| 排序轴 Ordination axe | 1 | 2 | 3 | 4 | 总和 Total | |
| 特征值Eigenvalue | CK | 0.581 | 0.038 | 0.004 | 0.000 | 0.623 |
| LW | 0.617 | 0.045 | 0.003 | 0.000 | 0.665 | |
| HW | 0.827 | 0.005 | 0.001 | 0.000 | 0.833 | |
| 关系值Correlation | CK | 0.786 | 0.856 | 0.666 | 0.000 | 2.308 |
| LW | 0.812 | 0.900 | 0.626 | 0.000 | 2.338 | |
| HW | 0.914 | 0.833 | 0.643 | 0.000 | 2.390 |
本研究采用OTC作为增温装置, 设置低、高两个增温梯度, 分别使空气温度最高增加1.36℃和2.81℃。在那曲站进行的梯度增温实验认为, OTC高度越高、开口越小, 其增温效果就越明显[23], 这与本研究的结论相一致。OTC由于阻挡了空气的水平运动, 空气由上端口进入后不易流通, 起到聚热效果, 因而箱内空气温度升高[23]。本研究得到增温对空气温湿度均产生正效应且空气温度增加显著。空气温度的显著升高增强土壤水分蒸发和植物蒸腾, 但由于近地面植被层和OTC的阻挡作用, 水分不易扩散, 从而使得OTC内空气湿度增加。李英年等[24]研究得出OTC内地表蒸散较对照高, 导致OTC内地表至植物层出现暂时相对干燥的低湿度环境, 有利于禾草类植物生长发育。这与本研究的结论相类似, 本研究测定的空气温湿度是距地面20 cm高度的植被冠层, 受植被盖度影响较大, 因而在近地面易形成空气湿度相对较高的环境。本研究也得到, 增温加强了植被与水分因子的关系, 并显著影响植被盖度。然而, 空气温湿度对增温的响应还与增温幅度有关。本研究得到, 高度增温处理下空气湿度在五鹿山减小0.40%, 在关帝山和管涔山分别增加5.29%和0.94%, 这与增温幅度过高引起的干旱有关。五鹿山地处吕梁山系的低纬度地区, 其空气温度在增温处理下升幅最大, 其中在高度增温处理下可增加高达1.48℃, 远远高于关帝山(0.88℃)和管涔山(0.65℃)的温度升幅。在五鹿山的1.48℃温度升幅已经接近政府间气候变化专门委员会(IPCC)提出的1.5℃升温阈值, 对生态系统产生不可预料的后果, 本研究所出现的空气湿度在五鹿山呈减小趋势便是其中之一, 与增温幅度过高引起的干旱有关, 这也是《IPCC全球升温1.5℃特别报告》中所验证了的。
本研究得到增温对土壤温湿度产生一定的负效应。其中增温对土壤湿度产生负效应, 这与大多数学者的研究一致, 即增温增强了土壤水分蒸发, 导致土壤出现暖干化趋势[23, 25-26]。而本研究得出在增温处理下土壤温度出现下降趋势, 这与数据采集时的降雨天气有关。降雨的干扰使得土壤水分随土层的变化出现异常, 图 3中也显示出土壤水分随土层加深而增加。因此, 土壤水分由于降雨干扰而突然增加, 影响了增温对土壤温度的效应。有研究表明土壤温度与水分呈线性负相关关系[27], 图 3也显示出土壤温度与土壤水分随土层呈相反的变化趋势。因而, 雨水下渗导致的土壤水分增加, 会降低土壤温度, 如果这种减弱程度超过了增温对土壤温度的增加程度, 便会出现增温下土壤温度降低的情况。还有研究得出温度较高、热量条件较好的地区, 降水为生长季植被覆盖度变化的主导因子[28]。前期对吕梁山亚高山草甸的研究也得出, 物种多样性在时空上呈现出受降水影响较大, 且由北向南逐渐升高的变化格局[21]。五鹿山为吕梁山的最南端, 处于整个山系的低纬度地区, 温度较高, 在降水起主导作用的条件下, 该区的增温效应受降水影响较大。因此, 降水增加可能会减弱增温对土壤的效应, 从而出现增温对土壤温度产生负效应的情况。
3.2 植被群落特征对增温的响应本研究中短期增温显著改变了群落盖度, 群落密度、频度和高度呈增加趋势但不明显。在青藏高原进行的短期增温实验也表明, 增温对高寒植被有正效应, 但各植被指标的方差分析都未达到显著水平, 表明短期增温对植被影响不显著[23]。2年的模拟增温实验得到, 增温使高寒草甸植物群落的高度整体增加, 多数物种的密度有所增加, 处于群落上层的建群种和主要伴生种的盖度有所增加[29]。对矮嵩草(Kobresia humilis)草甸的研究也发现, 多数物种的高度和密度均有所增加[30]。利用红外线辐射器作为增温装置的实验发现, 物种频度先减小后增大, 增温降低了美丽风毛菊(Saussurea pulchra)的频度, 却增加了黑褐苔草(Carex atrofusca)和珠芽蓼(Polygonum viviparum)的频度[31]。在内蒙古荒漠草原进行的长期增温实验发现, 群落高度整体增加[32]。在松嫩草原的增温实验也表明, 增温对羊草(Leymus chinensis)种群高度的影响不显著, 而对密度具有一定的促进作用[33]。这些研究位于我国不同生态区, 选择的植被类型、采用的增温装置、实验的持续时间均有所不同, 但结论与本研究基本一致, 说明增温在一定程度上促进植物生长, 影响群落发育, 即增温对植被产生的效应具有时间尺度依赖性。
本研究在黄土高原东部进行梯度增温实验, 得到低度增温下植物群落特征指数的增幅基本都是略高于高度增温。在青藏高原的实验也表明, 增温显著促进了沼泽草甸的生长, 但大幅度增温抑制了这种促进作用[29]。这说明不同增温水平对整个植被群落特征有一定影响:增温幅度较低时, 群落的高度、盖度、生物量及多样性有一定程度增加;但当温度升高超过一定值, 群落的盖度、高度、地上生物量又逐渐降低[23]。之所以会这样, 很大程度上是由于大幅度增温加剧了土壤水分蒸发, 不利于植物生长, 从而导致高度增温条件下植被群落特征呈下降趋势。因此, 增温要控制在一定幅度内, 才能对植被产生正效应。
另外, 本研究还得到, 低度增温处理下植物密度和频度在关帝山分别减小6.05株/m2和1.09%, 但在五鹿山和管涔山均呈增加趋势。从草本群落结构来看, 五鹿山中海拔开始群落中的优势种均为披针苔草, 且随纬度增加优势种密度增加, 但物种较为贫乏。关帝山地处吕梁山系的中等纬度地区, 其林下草本群落以莎草科的披针苔草为主, 且在增温处理下群落物种数未出现显著变化。因此, 增温处理下关帝山植物密度和频度呈减小趋势, 很可能与其它物种的优势度发生变化有关。在今后的研究中, 我们将群落内的所有物种划分成禾草、莎草、杂草三种植物功能型, 对其在增温处理下的动态变化进行深入研究。
3.3 植被与温度、水分因子关系对增温的响应通过探究增温处理下植被与温度、水分因子的关系, 得到水分因子对增温响应不敏感, 可能是由于增温时间短(只有1年), 也有可能是受降雨天气影响;但增温加强了植被与水分因子的关系, 促进植物对水分的依赖性, 进而显著影响植被盖度。气温升高将加剧植被对水的需求, 当降水较少的区域温度升高时, 植被出现退化现象[34]。这首先表现在高寒地区, 土壤水分是影响高寒草甸植被盖度的重要因素, 而增温导致土壤水分显著降低, 在降雨较少的生长季初期尤为显著, 这对植物生长发育将产生重要影响[35]。因此, 在气候暖化背景下, 降水下降, 土壤有效水分减少, 土壤湿度降低, 随着气温持续上升, 植被蒸散作用加强, 导致高寒植被生长速度降低, 植被呈现退化趋势[36]。其次在黄土高原地区, 由于温度上升加速了地表蒸散发过程, 潜在地加剧了水分缺乏, 从而造成黄土高原地区土壤干层的发育, 对植被生长具有明显的抑制作用[37]。由此可见, 不管是高寒地区[38-41]还是黄土高原[12, 21-22]地区, 增温加强植物对水的依赖性, 暖干化不利于植被生长;在水分较充沛的地区, 温度成为植被生长的主导因子, 因而增温可促进植物生长。
4 结论空气-土壤水热因子对增温响应差异较大。增温对空气温湿度产生正效应, 且空气温度的响应程度具有明显的海拔梯度格局, 但空气湿度对增温响应不敏感, 其响应程度呈现中间高值的空间变化格局。增温对土壤温湿度产生负效应, 且土壤温度的响应程度具有明显的纬度梯度格局, 但土壤水分对增温响应不敏感, 其响应程度呈现与土壤温度相反的海拔梯度格局。因此, 温度因子对增温响应更大, 且空气温度具有海拔依赖性, 土壤温度具有纬度依赖性。
增温对植被群落特征产生正效应。增温显著影响植物盖度, 但植物密度、频度、高度对增温响应不敏感, 其响应程度均呈现减小的纬度梯度格局和增大的海拔梯度格局。增温下植被与温度、水分因子的关系趋于增强, 而且植被与水分因子的关系对增温响应更为迅速。可见, 在水分缺乏的吕梁山区, 增温会加强植被与水分的关系, 促进植物对水的依赖性。但增温要控制在一定幅度内(本研究中为 < 1.36℃), 才能对植被产生正效应, 从而促进群落发育。
然而, 在水分为限制因子的黄土高原地区, 增温效应受降雨影响较大, 由降雨发生导致的水分增加可能会减弱增温对土壤的效应。因此, 增温的短期效应易受到降雨等环境条件的影响, 使实验结果出现误差。同时, 我们在青藏高原的研究也表明, 短期增温很难作出长期预测, 不能全面解释全球变暖的内在机制。建议加强增温试验的时间尺度, 以获取长时间序列的实验数据, 进而揭示山地增温效应的空间变化格局。
| [1] |
Colwell R K, Lees D C. The mid-domain effect:geometric constraints on the geography of species richness. Trends in Ecology & Evolution, 2000, 15(2): 70-76. |
| [2] |
雷彩芳, 上官铁梁, 赵冰清, 张婕, 许驭丹, 郭东罡. 灵空山采伐干扰下油松林林间草地物种多样性分析. 草业科学, 2014, 31(11): 2060-2068. |
| [3] |
杨丽霞, 任广鑫, 韩新辉, 康迪, 赵发珠, 邓健, 杨改河. 黄土高原退耕区不同林龄刺槐林下草本植物的多样性. 西北农业学报, 2014, 23(7): 172-178. |
| [4] |
Carnus J M, Parrotta J, Brockerhoff E, Arbez M, Jactel H, Kremer A, Lamb D, O'Hara K, Walters B. Planted forests and biodiversity. Journal of Forestry, 2006, 104(2): 65-77. |
| [5] |
Chastain Jr R A, Currie W S, Townsend P A. Carbon sequestration and nutrient cycling implications of the evergreen understory layer in Appalachian forests. Forest Ecology and Management, 2006, 231(1/3): 63-77. |
| [6] |
Gilliam F S. The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience, 2007, 57(10): 845-858. DOI:10.1641/B571007 |
| [7] |
农友, 卢立华, 贾宏炎, 雷丽群, 明安刚, 李华, 王亚南, 韦菊玲. 桂西南岩溶区不同恢复模式群落生物量及林下植物多样性. 林业科学研究, 2017, 30(2): 200-205. |
| [8] |
谭一波, 何琴飞, 郑威, 彭玉华, 侯远瑞, 何峰, 申文辉. 珠江流域中上游防护林冠层结构对林下植被的影响. 生态学杂志, 2016, 35(12): 3148-3156. |
| [9] |
黄先寒, 兰国玉, 杨川, 肖笔文, 吴志祥, 陶忠良. 云南橡胶林林下植物群落物种多样性. 生态学杂志, 2017, 36(8): 2138-2148. |
| [10] |
Wang G H, Zhou G S, Yang L M, Li Z Q. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecology, 2003, 165(2): 169-181. |
| [11] |
Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227. DOI:10.1038/35012228 |
| [12] |
Xu M H, Ma L, Jia Y Y, Liu M. Integrating the effects of latitude and altitude on the spatial differentiation of plant community diversity in a mountainous ecosystem in China. PLoS One, 2017, 12(3): e0174231. DOI:10.1371/journal.pone.0174231 |
| [13] |
Kessler M. Elevational gradients in species richness and endemism of selected plant groups in the central Bolivian Andes. Plant Ecology, 2000, 149(2): 181-193. |
| [14] |
焦翠翠, 于贵瑞, 何念鹏, 马安娜, 葛剑平, 胡中民. 欧亚大陆草原地上生物量的空间格局及其与环境因子的关系. 地理学报, 2016, 71(5): 781-796. |
| [15] |
Karl T R, Arguez A, Huang B Y, Lawrimore J H, McMahon J R, Menne M J, Peterson T C, Vose R S, Zhang H M. Possible artifacts of data biases in the recent global surface warming hiatus. Science, 2015, 348(6242): 1469-1472. DOI:10.1126/science.aaa5632 |
| [16] |
Roberts C D, Palmer M D, McNeall D, Collins M. Quantifying the likelihood of a continued hiatus in global warming. Nature Climate Change, 2015, 5(4): 337-342. DOI:10.1038/nclimate2531 |
| [17] |
Smith S J, Edmonds J, Hartin C A, Mundra A, Calvin K. Near-term acceleration in the rate of temperature change. Nature Climate Change, 2015, 5(4): 333-336. |
| [18] |
Marotzke J, Forster P M. Forcing, feedback and internal variability in global temperature trends. Nature, 2015, 517(7536): 565-570. DOI:10.1038/nature14117 |
| [19] |
Peñuelas J, Sardans J, Estiarte M, Ogaya R, Carnicer J, Coll M, Barbeta A, Rivas-Ubach A, Llusià J, Garbulsky M, Filella I, Jump A S. Evidence of current impact of climate change on life:a walk from genes to the biosphere. Global Change Biology, 2013, 19(8): 2303-2338. DOI:10.1111/gcb.12143 |
| [20] |
Xia J Y, Chen J Q, Piao S L, Ciais P, Luo Y Q, Wan S Q. Terrestrial carbon cycle affected by non-uniform climate warming. Nature Geoscience, 2014, 7(3): 173-180. DOI:10.1038/ngeo2093 |
| [21] |
张世雄, 杨晓艳, 温静, 徐满厚. 山西吕梁山亚高山草甸物种多样性的时空变化格局. 生态学报, 2018, 38(18): 6685-6693. |
| [22] |
杨晓艳, 张世雄, 温静, 徐满厚. 吕梁山森林群落草本层植物物种多样性的空间格局及其对模拟增温的响应. 生态学报, 2018, 38(18): 6642-6654. |
| [23] |
姜炎彬, 范苗, 张扬建. 短期增温对藏北高寒草甸植物群落特征的影响. 生态学杂志, 2017, 36(3): 616-622. |
| [24] |
李英年, 赵亮, 赵新全, 周华坤. 5年模拟增温后矮嵩草草甸群落结构及生产量的变化. 草地学报, 2004, 12(3): 236-239. |
| [25] |
王谋, 李勇, 白宪洲, 黄润秋. 全球变暖对青藏高原腹地草地资源的影响. 自然资源学报, 2004, 19(3): 331-336. |
| [26] |
王根绪, 李娜, 胡宏昌. 气候变化对长江黄河源区生态系统的影响及其水文效应. 气候变化研究进展, 2009, 5(4): 202-208. |
| [27] |
车宗玺, 李进军, 汪有奎, 丁国民, 李世霞, 邸华, 车宗彩, 潘欣, 毛志明, 郭朝霞. 祁连山西段草地土壤温度、水分变化特征. 生态学报, 2018, 38(1): 105-111. |
| [28] |
陆晴, 吴绍洪, 赵东升. 1982~2013年青藏高原高寒草地覆盖变化及与气候之间的关系. 地理科学, 2017, 37(2): 292-300. |
| [29] |
李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895-905. |
| [30] |
周华坤, 周兴民, 赵新全. 模拟增温效应对矮嵩草草甸影响的初步研究. 植物生态学报, 2000, 24(5): 547-553. |
| [31] |
陈翔, 彭飞, 尤全刚, 王涛, 薛娴. 高寒草甸植被特征对模拟增温的响应——以青藏高原多年冻土区为例. 草业科学, 2016, 33(5): 825-834. |
| [32] |
张东, 钞然, 万志强, 杨劼, 包铁军, 高清竹. 模拟增温增雨对典型草原优势种羊草功能性状的影响. 草业科学, 2018, 35(8): 1919-1928. |
| [33] |
齐红, 罗微, 孙亚娟, 郭继勋. 松嫩草原羊草种群数量特征对增温和施氮的响应. 东北师大学报:自然科学版, 2013, 45(2): 112-117. |
| [34] |
徐兴奎, 陈红, Levy J K. 气候变暖背景下青藏高原植被覆盖特征的时空变化及其成因分析. 科学通报, 2008, 53(4): 456-462. |
| [35] |
徐振锋, 胡庭兴, 李小艳, 张远彬, 鲜骏仁, 王开运. 川西亚高山采伐迹地草坡群落对模拟增温的短期响应. 生态学报, 2009, 29(6): 2899-2905. |
| [36] |
Xu M H, Ma L, Xue X, Zhai D T. Warming effects on plant biomass allocation and correlations with the soil environment in an alpine meadow, China. Journal of Arid Land, 2016, 8(5): 773-786. DOI:10.1007/s40333-016-0013-z |
| [37] |
信忠保, 许炯心, 郑伟. 气候变化和人类活动对黄土高原植被覆盖变化的影响. 中国科学D辑:地球科学, 2007, 37(11): 1504-1514. |
| [38] |
徐满厚, 薛娴. 青藏高原高寒草甸夏季植被特征及对模拟增温的短期响应. 生态学报, 2013, 33(7): 2071-2083. |
| [39] |
徐满厚, 刘敏, 翟大彤, 薛娴, 彭飞, 尤全刚. 青藏高原高寒草甸生物量动态变化及与环境因子的关系——基于模拟增温实验. 生态学报, 2016, 36(18): 5759-5767. |
| [40] |
徐满厚, 刘敏, 翟大彤, 薛娴, 彭飞, 尤全刚. 模拟增温对青藏高原高寒草甸根系生物量的影响. 生态学报, 2016, 36(21): 6812-6822. |
| [41] |
徐满厚, 薛娴. 青藏高原高寒草甸植被特征与温度、水分因子关系. 生态学报, 2013, 33(10): 3158-3168. |