 2020, Vol. 40
2020, Vol. 40文章信息
- 赵洋洋, 祁玥, 王晓宁, 赵伟
- ZHAO Yangyang, QI Yue, WANG Xiaoning, ZHAO Wei
- 青海沙蜥消化道组织结构及嗜银细胞研究
- Histology of and argyrophil cells in digestive tract of Phrynocephalus vlangalii
- 生态学报. 2020, 40(16): 5855-5861
- Acta Ecologica Sinica. 2020, 40(16): 5855-5861
- http://dx.doi.org/10.5846/stxb201901130106
-
文章历史
- 收稿日期: 2019-01-13
- 网络出版日期: 2020-06-08

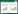




青海沙蜥(Phrynocephalus vlangalii)隶属于爬行纲(Reptilia), 蜥蜴目(Lacertiformes), 鬣蜥科(Agamidae), 沙蜥属(Phrynocephalus), 广泛分布在青藏高原2700—4500 m的荒漠和半荒漠干旱地区[1], 是研究爬行动物对高海拔适应的理想物种之一。近年来, 为探究爬行动物高海拔适应机制, 国内学者对青海沙蜥的生理特征、生活史特征、肠道微生物、转录组等方面进行了深入的研究[2-7]。
高海拔地区的低温低氧对高原土著动物, 造成了巨大的选择压力[8]。低温低氧限制动物的能量收入, 从而影响能量输出, 如随着海拔梯度的升高, 青海沙蜥雌性的繁殖输出下降[2-3]; 标准代谢速率下降[4]。然而有研究表明, 青海沙蜥高海拔种群虽然新生幼仔更小, 但生长速率更快, 从而在更短的时间内达到性成熟[5], 说明沙蜥可能通过其他的适应性进化增强自身的能量吸收或利用。
动物消化系统是摄取、消化和吸收能量的主要场所, 其功能受组织结构和肠道微生物的共同影响。为适应高海拔低温低氧的影响, 青海沙蜥消化系统随海拔升高而变化, 如胃的相对质量增大[9]; 肠道微生物组成发生改变[6]。然而, 高海拔能量限制是否会导致沙蜥消化道组织结构上的适应性变化仍未可知。为进一步理解青海沙蜥对高海拔的适应, 本文通过解剖和组织切片对其消化系统的组织结构和嗜银细胞进行了研究, 以期为揭示爬行类的高原适应机制提供科学依据。
1 材料与方法 1.1 材料实验材料取自保存在兰州大学生命科学学院的青海沙蜥成体标本20只(雌雄各10只)。标本保存在8%的福尔马林溶液中, 系2010年采集自青海省海西州(37°09′ N, 97°35′ E, 2980 m)。
1.2 方法解剖取其消化道, 并区分食管、贲门、胃体、幽门、十二指肠、空肠、回肠、直肠各部分。首先用游标卡尺测量各部位的长度, 精确到0.01 mm, 然后分别取各部分组织约5 mm进行切片。将固定好的组织进行酒精梯度脱水, 透明, 浸蜡, 包埋与修蜡, 并用KD-2020A切片机切成6 μm厚的条带。贴片后进行梯度脱蜡与复水。每个部位的装片分成两组, 其中一组进行H.E染色, 另一组采用Grimelius银染色法[10]进行染色。脱水透明后用树胶封片以备镜检。
用CX 40显微镜对装片进行观察, 每张装片随机选取3个视野(目镜10×, 物镜40×), 并用OD630K成像系统拍照, 使用MvImage vt软件对照片进行分析, 测量消化道各部位厚度, 精确到0.01 μm, 并记录每个部位的嗜银细胞数量。
1.3 数据分析由于消化道长度与吻肛长(Snout-vent length, SVL)显著正相关(r=0.289, P < 0.001), 故将消化道各段长度对SVL进行线性回归, 保留未标准化的残差作为SVL校正后的长度用于后续分析。采用two-way ANOVA分析消化道各段长度(SVL校正后)、管壁厚度和噬银细胞数量的差异, 其中长度、厚度或噬银细胞数量作为因变量, 性别和部位作为因子, 并用Tukey′s b进行多重比较分析。本文所有数据分析均使用SPSS 22完成, 数值均以mean±SE形式表示, 显著水平设置为a=0.05。
2 实验结果 2.1 消化管形态学青海沙蜥的消化道管壁可以分为4层结构, 由管腔内向外可依次分为黏膜层、黏膜下层、肌层和浆膜层。
食管 细长的管状结构, 位于消化道的前端, 前接咽部后接贲门部, 分界并不明显。长(10.60±0.26) mm, 管壁厚(71.90±4.71) μm。食管内腔分布有许多凸起的黏膜皱襞, 分支10—15个; 黏膜上皮为排列整齐的单层柱状上皮。黏膜肌层较薄。黏膜下层明显且较厚, 由疏松结缔组织构成, 内分布有血管和淋巴管。肌层分为两层:内侧的环肌和外侧的纵肌, 以内环肌为主, 内外肌均为平滑肌。浆膜较薄且明显。在食管中可见分布有大量的椭圆形的杯状细胞(图 1)。

|
| 图 1 青海沙蜥消化道各部分显微结构 Fig. 1 Microstructure of various parts of digestive tract of P. vlangalii Gc:杯状细胞Goblet cell; Mm:黏膜肌层Muscularis mucosa; Cm:环肌Circular muscle; Lm:纵肌Longitudinal muscle; Cg:贲门腺Cardiac gland; Fg:胃底腺Fundic gland; Bv:血管Blood vessel; Pg:幽门腺Pyloric gland; Ig:肠腺Intestinal gland; Cl:中央乳糜管Central lacteal; Iv:小肠绒毛Intestinal villi; Tm:肌层Tunica muscularis |
胃 最为膨大的部分, 呈U型, 从上端到下端依次分为贲门、胃体、幽门, 各部分无明显的分界线, 全长(18.78±0.44) mm; 贲门部位于胃的最上端, 黏膜凸起形成皱襞, 排列规则且分枝较少, 管壁厚(309.06±26.44) μm; 胃体部最为粗大, 腔内有大量纵行连续、排列整齐的黏膜皱襞, 管壁厚(303.16±15.77) μm。幽门部的皱襞同样排列规则整齐, 并且有一个粗大的皱襞, 管壁厚(442.74±24.73) μm。胃中具有发达的贲门腺、胃底腺和幽门腺, 并有黏膜上皮下陷形成的胃小凹。黏膜肌层薄且明显。黏膜下层较厚, 分布有血管和淋巴组织。肌层由内环外纵的两层平滑肌构成, 肌层平均厚度为(107.05±3.64) μm(图 1)。
小肠 消化道内最为细长的部分, 由前至后依次分为十二指肠、空肠和回肠, 全长(42.94±1.14)mm。十二指肠与幽门分界明显, 形状弯曲, 长(11.70±0.18) mm, 管径较粗, 管壁厚(76.66±6.06) μm。空肠长(11.45±0.37) mm, 管径较十二指肠略细, 管壁厚(77.65±5.58)μm。回肠长(19.85±0.70) mm, 是小肠中最细长的部分, 管壁厚(71.86±6.61) μm。肠腔内的绒毛发达, 无分枝, 十二指肠绒毛较为细长, 多呈弯曲的指状, 约11—13个; 空肠内的绒毛较为粗大, 多呈直立状或弯曲状, 约17—20个; 回肠部绒毛呈直立或弯曲状, 约15—17个。小肠中还具有发达的肠腺和中央乳糜管, 黏膜肌层薄且明显; 黏膜下层分布有血管、淋巴管等; 肌层同样由内环肌和外纵肌两层平滑肌构成, 浆膜较薄(图 1)。
直肠 管径明显增粗, 与小肠的分界明显。长(21.71±0.37) mm, 管壁厚(212.74±12.64) μm。黏膜上皮中分布有大量的杯状细胞, 比小肠部增多; 黏膜皱襞不发达, 变的低矮粗大。黏膜下层较厚, 有血管分布直肠的肌层明显变厚, 以环形肌为主; 浆膜膜为上皮细胞(图 1)。
统计发现, 性别对消化道各段长度没有显著性影响(F1, 227=1.495, P=0.223), 各段的长度差异显著(F5, 227=141.328, P < 0.001) (图 2), 性别与部位之间交互作用并不显著(F5, 227=0.384, P=0.859)。同样, 性别对消化道各段管壁厚度没有显著性影响(F1, 144=1.139, P=0.288), 各段的厚度差异显著(F7, 144=91.285, P < 0.001) (图 2), 性别与部位之间交互作用并不显著(F7, 144=1.501, P=0.171)。

|
| 图 2 青海沙蜥消化道各段长度和管壁厚度的对比 Fig. 2 Comparison of the length and wall thickness of digestive tract of P. vlangalii |
嗜银细胞在青海沙蜥的消化道中广泛分布, 其中胃体部分布密度最大, 贲门部次之, 回肠部最少(表 1)。整体而言, 从食管开始下行至直肠, 青海沙蜥嗜银细胞的分布密度曲线形状类似于倒“V”形。
| 部位 Region |
分布密度(个/视野)D istribution density(number/field of vision) |
显著水平 Significance level |
| 食管Esophagus | 4.33±0.33 | b |
| 贲门Cardia | 10.10±0.46 | a |
| 胃体Gastric body | 10.45±0.45 | a |
| 幽门Pylorus | 4.53±0.16 | b |
| 十二指肠Duodenum | 3.33±0.26 | b |
| 空肠Jejunum | 3.87±0.41 | b |
| 回肠Ileum | 3.08±0.37 | b |
| 直肠Rectum | 3.50±0.31 | b |
| 样本数量n=20;同列数据, 具有相同小写字母表示不存在显著差异(P > 0.05), 具有完全不同的小写字母存在显著差异(P < 0.05) | ||
统计表明, 性别对消化道各段的嗜银细胞密度没有显著性影响(F1, 144=0.017, P=0.896), 各部位嗜银细胞密度差异显著(F7, 144=71.863, P < 0.001) (表 1), 性别与部位之间交互作用并不显著(F7, 144=0.474, P=0.852)。
青海沙蜥消化道嗜银细胞的形状多样, 食管胃体幽门十二指肠直肠部位的嗜银细胞主要呈锥体形或梭形, 贲门空肠回肠的嗜银细胞则主要呈椭圆形。嗜银细胞主要分布于上皮细胞基部或上皮细胞之间(图 3)。

|
| 图 3 青海沙蜥消化道各段嗜银细胞的形状与分布 Fig. 3 The shape and distribution of argyrophil cell in each part of digestive tract of P. vlangalii 图中箭头所指即为嗜银细胞 |
消化系统是动物摄食、消化和吸收的主要场所, 决定了动物对食物消化和吸收的速率[11-12]。小肠是营养物质吸收的主要部位, 与低海拔蜥蜴[13-14]相比, 青海沙蜥小肠(十二指肠、空肠和回肠)的占比更高, 有利于增强对营养物质的吸收。其中, 钙等物质主要在十二指肠内被吸收; 糖类、蛋白质和脂肪的消化产物及维生素、水和无机盐主要在十二指肠和空肠被吸收; 回肠可主动吸收胆盐和维生素B12。研究表明, 高海拔蜥蜴脂肪代谢率增加, 而胆盐和维生素B12对脂肪的消化吸收具有重要意义。此外, 钙离子有助于心肌收缩, 促进血液循环并增加氧气供应[15-18]。这说明青海沙蜥肠道结构的变化有助于对高海拔低温低氧的适应。
消化道形态结构的变化对动物自身能量平衡和食物变异的适应具有重要作用[19]。与丽斑麻蜥(Eremias argus)[20]或蜡皮蜥(Leiolepis reevesii)[21]的复层鳞状上皮不同, 青海沙蜥的食管上皮为单层柱状上皮。区别于丽斑麻蜥的骨骼肌肌层[20], 青海沙蜥食管肌层为平滑肌, 且食管内具有发达的黏膜皱襞和可分泌粘液的杯状细胞, 有利于润滑食物和保护黏膜[22]。青海沙蜥胃内具有连续的发达皱襞和肌层, 尤其是内环肌特别厚, 可以扩大胃的容积, 并增强胃的蠕动能力。与蜡皮蜥[21]、凹耳臭蛙(Odorrana tormota)[23]、山溪鲵(Ba trachuperus pinchonii)[24]的小肠不同, 青海沙蜥具有空肠且中央乳糜管特别发达, 可能与增强消化吸收能力有关。青海沙蜥直肠内有少量粗大皱襞, 杯状细胞增多, 可以有效润滑食物残渣和粪便, 并且较厚的肌层增强了直肠的蠕动, 有利于残渣和粪便的排出。
3.2 嗜银细胞嗜银细胞在脊椎动物(圆口纲除外)消化道内广泛分布[25], 且在消化道内的分布特点存在种间差异[26]。与绝大多数两栖爬行动物研究相同[27-29], 青海沙蜥的嗜银细胞广泛分布在消化道各部分。在秦岭滑蜥(Scincella tsinlingensis)[14]、丽斑麻蜥[30]中, 嗜银细胞在胃体部分布最高。黑龙江草蜥(Takydromus amurensis)[31]、白条草蜥(Takydromus wolteri)[32]中, 嗜银细胞在幽门部分布最高。多疣壁虎(Gekko japonicus)[33]的嗜银细胞在十二指肠分布密度最高。变色树蜥(Calotes versicolor)[34]在空肠分布最高。青海沙蜥的的嗜银细胞在胃体部分布密度最高, 其次是贲门部, 这种分布情况与丽斑麻蜥[30]类似。
嗜银细胞是一种内分泌细胞, 主要分泌5-羟色胺(5-hydroxy tryptamine, 5-HT), 在调节消化功能中起着重要作用[35]。丽斑麻蜥的消化道内具有圆形和椭圆形的嗜银细胞, 分泌的激素不进入管道, 直接进入血液, 具有内分泌的功能[30]。有学者认为内分泌细胞可能以外分泌方式将5-HT排到肠腔, 但形态学依据不足[36]。本研究发现青海沙蜥消化道内有锥体形的嗜银细胞, 其尖端朝向消化道肠腔或者朝向腺体, 这种形状和分布特点可能揭示了嗜银细胞的外分泌功能。还有些学者认为, 嗜银细胞具有内、外分泌功能[37-38]。青海沙蜥的消化道内观察到形状类似于长梭形的嗜银细胞, 其两端分别指向消化腔和固有层, 表明这种内分泌细胞可能具有内分泌和外分泌两种功能。
3.3 结论综上, 青海沙蜥小肠占消化道总长的比例增加延长了食物在消化道内的停留时间, 增强了对钙、脂类和维生素等营养物质的吸收能力; 肠道内中央乳糜管发达, 加强了吸收营养物质的能力; 嗜银细胞在消化道内广泛分布, 以胃部最多, 其分泌物可有效的增强消化能力。本文表明, 青海沙蜥消化道特有的组织结构特点为青海沙蜥适应高海拔环境提供了有力的保障。
| [1] |
赵肯堂. 中国沙蜥属的分类和分布研究. 内蒙古大学学报:自然科学版, 1979(2): 111-121. |
| [2] |
Jin Y T, Liu N F. Altitudinal variation in reproductive strategy of the toad-headed lizard, Phrynocephalus vlangalii in North Tibet Plateau (Qinghai). Amphibia-Reptilia, 2007, 28(4): 509-515. DOI:10.1163/156853807782152507 |
| [3] |
Li J, Zhou R, Liu N. Life-history variation among three populations of the toad-headed lizard Phrynocephalus vlangalii along an elevation gradient on the northeastern Tibetan Plateau. The Herpetological Journal, 2014, 24(1): 17-23. |
| [4] |
Li W X, Liang S W, Wang H H, Xin Y, Lu S S, Tang X L, Chen Q. The Effects of chronic hypoxia on thermoregulation and metabolism in Phrynocephalus vlangalii. Asian Herpetological Research, 2016, 7(2): 103-111. |
| [5] |
Lu H L, Xu C X, Jin Y T, Hero J M, Du W G. Proximate causes of altitudinal differences in body size in an agamid lizard. Ecology and Evolution, 2018, 8(1): 645-654. |
| [6] |
Zhang W Y, Li N, Tang X L, Liu N F, Zhao W. Changes in intestinal microbiota across an altitudinal gradient in the lizard Phrynocephalus vlangalii. Ecology and Evolution, 2018, 8(9): 4695-4703. DOI:10.1002/ece3.4029 |
| [7] |
Yang W Z, Qi Y, Fu J Z. Exploring the genetic basis of Adaptation to high elevations in reptiles:a comparative transcriptome analysis of two toad-headed agamas (genus Phrynocephalus). PLoS One, 2014, 9(11): e112218. DOI:10.1371/journal.pone.0112218 |
| [8] |
Storz J F, Scott G R, Cheviron Z A. Phenotypic plasticity and genetic adaptation to high-altitude hypoxia in vertebrates. The Journal of Experimental Biology, 2010, 213(24): 4125-4136. DOI:10.1242/jeb.048181 |
| [9] |
Han J M, Guo R H, Li J Q, Guan C, Chen Y, Zhao W. Organ mass variation in a toad headed lizard Phrynocephalus vlangalii in response to hypoxia and low temperature in the Qinghai-Tibet Plateau, China. PLoS One, 2016, 11(9): e0162572. DOI:10.1371/journal.pone.0162572 |
| [10] |
Grimelius L. The argyrophil reaction in islet cells of adult human pancreas studies with a new silver nitrate procedure. Acta Societatis Medicorum Upsaliensis, 1968, 73(5/6): 271-294. |
| [11] |
Corp N, Gorman M L, Speakman J R. Apparent absorption efficiency and gut morphometry of wood mice, Apodemus sylvaticus, from two distinct populations with different diets. Physiological Zoology, 1997, 70(6): 610-614. DOI:10.1086/515876 |
| [12] |
Gross J E, Wang Z W, Wunder B A. Effects of food quality and energy needs:changes in gut morphology and capacity of Microtus ochrogaster. Journal of Mammalogy, 1985, 66(4): 661-667. DOI:10.2307/1380792 |
| [13] |
王昱. 丽纹龙蜥消化管的组织结构观察. 北京联合大学学报:自然科学版, 2011, 25(3): 41-46. DOI:10.3969/j.issn.1005-0310.2011.03.011 |
| [14] |
王丽敏, 杨纯, 郭俐. 秦岭滑蜥消化系统组织结构及消化管嗜银细胞观察. 动物学杂志, 2016, 51(4): 614-622. |
| [15] |
Tang X L, Xin Y, Wang H H, Li W X, Zhang Y, Liang S W, He J Z, Wang N B, Ma M, Chen Q. Metabolic characteristics and response to high altitude in Phrynocephalus erythrurus (Lacertilia:Agamidae), a lizard dwell at altitudes higher than any other living lizards in the world. PLoS One, 2013, 8(8): e71976. DOI:10.1371/journal.pone.0071976 |
| [16] |
王启华, 邱学才. 临床人体解剖生理学. 广州: 中山大学出版社, 2007: 381-457.
|
| [17] |
Thakkar K, Billa G. Treatment of vitamin B12 deficiency-Methylcobalamine? Cyancobalamine? Hydroxocobalamin?-clearing the confusion. European Journal of Clinical Nutrition, 2015, 69(1): 1-2. DOI:10.1038/ejcn.2014.165 |
| [18] |
Tibbits G F, Hamman B N. Regulation of myocardial contractility. Medicine and Science in Sports and Exercise, 1991, 23(10): 1140-1144. |
| [19] |
徐金会, 安书成, 邰发道. 棕色田鼠消化道形态变化与能量需求的关系. 动物学报, 2003, 49(1): 32-39. |
| [20] |
李洁, 尤旭. 丽斑麻蜥(Eremias argus)消化系统组织结构研究. 哈尔滨师范大学:自然科学学报, 1999, 15(6): 84-87. |
| [21] |
洪美玲, 傅丽容, 王力军, 黎江娃. 蜡皮蜥消化系统组织学初步研究. 四川动物, 2005, 24(3): 333-336. DOI:10.3969/j.issn.1000-7083.2005.03.018 |
| [22] |
钟慈声. 细胞和组织的超微结构. 北京: 人民卫生出版社, 1984: 12-29.
|
| [23] |
吴雨函, 熊荣川, 雷飞宙, 江建平. 凹耳臭蛙消化系统解剖学及组织学观察. 四川动物, 2012, 31(4): 583-588. DOI:10.3969/j.issn.1000-7083.2012.04.014 |
| [24] |
陈红, 陈欣, 赵尔宓. 山溪鲵消化系统和呼吸系统的解剖. 四川动物, 2009, 28(4): 565-568. |
| [25] |
唐晓雯, 郑一守. 不同脊椎动物消化道内5-羟色胺免疫染色细胞的分布. 动物学研究, 1991, 12(3): 293-298. |
| [26] |
谢冰楠, 李丕鹏, 陆宇燕. 爬行动物消化道嗜银细胞的研究进展. 四川动物, 2012, 31(3): 504-507. |
| [27] |
王晓平, 谢文海, 李家洲. 两种蜥蜴类动物胃肠道嗜银细胞的形态学观察及比较. 玉林师范学院学报, 2004, 25(5): 70-72. DOI:10.3969/j.issn.1004-4671.2004.05.020 |
| [28] |
张彦华, 高欣, 李淑兰. 棕黑锦蛇(E. schrenchii)消化道嗜银细胞的分布及形态学观. 哈尔滨师范大学:自然科学学报, 2005, 21(1): 77-81. |
| [29] |
王淼, 甄靓靓, 李淑兰, 赵文阁. 东亚腹链蛇消化道嗜银细胞的分布及形态学观察. 四川动物, 2007, 26(2): 278-280. DOI:10.3969/j.issn.1000-7083.2007.02.009 |
| [30] |
金琳姊, 张欣, 李淑兰, 赵文阁. 丽斑麻蜥消化道嗜银细胞的分布及形态学观察. 中国农学通报, 2012, 28(32): 119-122. DOI:10.3969/j.issn.1000-6850.2012.32.023 |
| [31] |
于萍, 李淑兰. 黑龙江草蜥(Takydromus amurensis)消化道嗜银细胞的研究. 黑龙江畜牧兽医, 2016(7): 226-228. |
| [32] |
王丽影, 任媛, 刘超, 李淑兰. 白条草蜥消化道嗜银细胞的分布和形态学观察. 四川动物, 2009, 28(2): 247-249. |
| [33] |
李伟, 李霖, 吴孝兵, 杜开和, 王义权. 多疣壁虎肠道内分泌细胞的分布及形态学观察. 动物学杂志, 2001, 36(1): 11-14. DOI:10.3969/j.issn.0250-3263.2001.01.003 |
| [34] |
李淑兰, 赵文艳, 冷超, 赵文阁. 变色树蜥消化道5-羟色胺细胞的免疫组织化学定位. 动物学研究, 2004, 25(5): 436-441. DOI:10.3321/j.issn:0254-5853.2004.05.011 |
| [35] |
Forssman W G, Orci L, Pictet R, Renold A E, Rouiller C. The endocrine cells in the epithelium of the gastrointestinal mucosa of the rat:an electron microscope study. The Journal of Cell Biology, 1969, 40(3): 692-715. DOI:10.1083/jcb.40.3.692 |
| [36] |
Nilsson O, Ahlman H, Geffard M, Dahlstrom A, Ericson L E. Bipolarity of duodenal enterochromaffin cells in the rat. Cell and Tissue Research, 1987, 284(1): 49-54. |
| [37] |
Bülbring E, Crema A. The action of 5-Hydroxytryptamine, 5-hydroxytryptophan and reserpine on intestinal peristalsis in anaesthetized guinea-pigs. The Journal of Physiology, 1959, 146(1): 29-53. DOI:10.1113/jphysiol.1959.sp006176 |
| [38] |
Uvnäs-Wallensten K, Rehfeld J F. Molecular forms of gastrin in antral mucosa, plasma and gastric juice during vagal stimulation of anesthetized cats. Acta Physiologica Scandinavica, 1976, 98(2): 217-226. DOI:10.1111/j.1748-1716.1976.tb00239.x |