 2020, Vol. 40
2020, Vol. 40文章信息
- 杨锐, 吴鹏飞, 魏雪
- YANG Rui, WU Pengfei, WEI Xue
- 天然高寒草地转变为燕麦人工草地对土壤线虫群落的影响
- Effects of the transformation from natural alpine grassland to artificial oat grassland on the soil nematode communities
- 生态学报. 2020, 40(14): 4903-4920
- Acta Ecologica Sinica. 2020, 40(14): 4903-4920
- http://dx.doi.org/10.5846/stxb201910152152
-
文章历史
- 收稿日期: 2019-10-15
- 网络出版日期: 2020-04-29






青藏高原是地球上海拔最高、面积最大、形成最晚的高原, 素有“地球第三极”和“亚洲水塔”之称, 是我国长江、黄河两大水系的重要水源涵养区, 其生态地位、生态价值及战略意义突出。位于青藏东缘的川西北高寒草地, 面积达16.6万km2, 是我国五大牧区之一, 支撑着高原畜牧业的发展[1]。然而, 受“高”、“寒”、“旱”等恶劣自然条件的影响, 该区域草地生态系统十分脆弱, 加之近年来, 在过度放牧和全球气候变化的影响下, 高寒草地出现大面积退化甚至沙化, 草地生产力下降, 草畜平衡失调, 一定程度上限制了畜牧业的发展[2]。人工草地因为具有优良的牧草品质和较高的生产力, 能够有效缓解草畜矛盾, 在一定程度上减轻天然草场的放牧压力, 近些年在青藏高原上得到快速发展[3]。截止2011年, 四川省人工草地面积已达1.82×107 hm2, 较1990年增加了约5.7倍, 其中主要集中在青藏东缘的阿坝州和甘孜州[4]。燕麦作为青藏高原人工草地的主要栽培种, 种植面积约占人工草地面积的70%[5], 不仅具有耐盐碱、抗旱耐寒、产量高等生物学特性, 而且营养丰富、适口性好, 是高寒地区高产、稳产、优质的重要牧草品种[6], 在维持高寒草地草畜平衡、改善草地生态环境和促进畜牧业稳定可持续发展等方面发挥着重要作用。目前, 关于燕麦人工草地的研究主要集中在植物生物量[7]、草地碳储量[8]和地下土壤的理化性质[9]及土壤微生物群落[10]变化等方面, 本课题组前期虽然对不同人工草地土壤节肢动物群落多样性进行过研究[11], 但该研究是在小块的种质资源圃(2 m × 3 m)内开展, 并非大田生产状态的人工草地, 且未对土壤线虫群落进行研究。因此, 目前关于青藏高原地区建植燕麦人工草地对土壤动物群落多样性的影响还不清楚, 制约到对人工草地生态系统生物多样性的认识。
土壤线虫是土壤动物的主要组成部分[12], 也是草地生态系统碎屑食物网的优势类群, 食性多样, 能够分布在土壤食物网的多个营养级[13], 对草地生态系统的物质循环、能量流动及地上植物群落的组成、演替及生产力起着重要的调控作用[14-15]。同时, 土壤线虫也具有对环境短期变化敏感, 种类丰富、分布广泛、易于鉴定[14, 16]等特点, 被看作评价草地土壤生态系统健康状况的典型指示生物[16]。鉴于此, 本研究以川西北高寒地区燕麦地为研究对象, 以天然草地为对照, 通过对土壤线虫群落的调查, 研究燕麦地与天然草地土壤线虫群落的差异, 查明天然高寒草地转变为燕麦地后土壤线虫群落和土壤食物网的变化趋势及主要影响因子。研究结果不仅可为评估人工草地的健康状况及可持续利用提供科学依据, 还可提供高寒人工草地土壤动物多样性方面的基础数据。
1 材料与方法 1.1 研究区概况研究区位于四川省阿坝藏族羌族自治州的红原县(102°35′—102°58′ E, 32°5′—32°83′N)。该区地处青藏高原东部, 分属长江、黄河两大水系, 平均海拔3600 m以上, 属大陆性高原寒温带季风气候, 气候特点冬长无夏, 春秋短。日照时间长, 全年日照2417.9 h, 太阳辐射年总量61.5 kJ/cm2。年均气温0.9℃, ,最高气温在7月, 平均气温10.9℃, 最低气温在12月, 平均气温-10.3℃;年均降水量753.0 mm, 主要集中在5—10月, 水热同期, 无绝对无霜期。土壤类型以弱酸性的亚高山草甸土为主, 局部有沼泽土和风沙土。主要植被类型为亚高山草甸, 植被覆盖度为70%—90%, 优势种为四川嵩草(Kobresia setchwaneusis)、小嵩草(Kobresia pygmaea)、垂穗披碱草(Elymus nutans), 伴生种为鹅绒委陵菜(Potentilla anserina)、草玉梅(Anemone rivularis)、紫穗鹅观草(Roegneria purpurascens)、高原毛茛(Ranunculus tanguticus)、矮嵩草(Kobresia tibetica)等。红原县境内天然草场面积达7.76×105 hm2, 占国土总面积的92.97%, 在相关政策和项目推动下, 截止2016年人工种草面积已达2.91×104 hm2[17]。
1.2 样地设置2014年5月在西南民族大学青藏高原畜牧业高科技研发示范基地中选取燕麦地和天然草地作为观测样地。每种草地内, 各设置2个50 m×50 m的固定大样方。在每个大样方内沿对角线方向选取3个10 m×10 m的小样方, 每个小样方间距约10 m左右。其中, 燕麦地从2013年开始种植, 种植前为天然草地, 已连续种植2年。种植前进行机耕翻耙, 然后人工整地、耙平、开沟条播, 播时施225 kg/hm2复合肥(氮、磷、钾养分≥45%)做基肥, 旱作无灌溉, 生长期人工除杂[18]。
1.3 样品采集和处理2014年7月和9月进行取样。在每个小样方内, 用土钻按照五点法取混合土样, 取样深度为0—15 cm, 均匀混合后放入有编号的自封袋内。共采集用于提取土壤线虫的混合土壤样24份。另外, 于9月份在各个样方内, 用RR-730M土壤湿度仪测定0—15 cm左右土壤体积含水量;并在各样方中采集0—15 cm的混合土样3份, 共采集用于分析土壤化学性质混合土样12份。室内经自然风干、研磨后, 用于测定化学性质。
实验室内, 称取50 g混合土样, 用湿漏斗法(Baermann法)分离土壤线虫, 分离时间为48 h, 用5%的甲醛溶液固定保存线虫。根据《中国土壤动物检索图鉴》[19]和《荷兰线虫》[20]在光学显微镜(Leica DM4000B)下鉴定到属, 并统计个体数量。鉴定中, 如果采集到的土壤线虫低于100条, 则全部鉴定;如果多于100条, 则随机取其中的100条进行鉴定, 并统计总数。根据土壤的含水量, 将线虫的个体数量转化为条/100g干土。
土壤化学性质依据《土壤农业化学分析方法》[21]测定。其中, 土壤pH采用电位法;土壤有机质采用重铬酸钾氧化法;全氮采用半微量开氏法;速效氮采用碱解扩散法;全磷采用硫酸—高氯酸消解, 钼锑抗比色法;速效磷采用氟化铵-盐酸消解, 钼锑抗比色法;全钾采用氢氟酸—高氯酸消煮, 火焰光度计法;速效钾采用乙酸铵浸提, 火焰光度计法。
1.4 数据处理与分析按照线虫个体数量进行优势度的划分:个体数占总数10%以上者为优势类群, 1%—10%为常见类群, 1%以下为稀有类群[22]。
根据线虫的形态学特征和取食生境把所有的土壤线虫分为:食细菌类群(Bacterivores, Ba)、食真菌类群(Fungivores, Fu)、植物寄生类群(Plant-parasites, Pl)、捕食-杂食类群(Predators-omnivores, Pr)4个营养类群[23]。
土壤线虫群落多样性分析:采用密度、类群数、Shannon-Weiner多样性指数和Simpson优势度指数计算土壤线虫群落的多样性[24]。计算公式如下:

|
式中, Pi为第i类群个体数占总个体数的比例, S为土壤线虫群落总类群数。
线虫成熟度指数:包括自由生活线虫成熟度指数(Free-living nematode maturity index, MI)、植物寄生线虫成熟度指数(Plant parasite index, PPI)用来评价土壤线虫对外界扰动的响应[15]。公式如下:
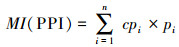
|
式中, cpi为非植物寄生线虫(植物寄生性)土壤线虫第i类群colonizer-persister值;n为非植物寄生线虫(植物寄生性)土壤线虫类群数;Pi为非植物寄生线虫(植物寄生性)土壤线虫第i类群个体数占总个体数的比例。MI值指示自由生活线虫r-选择和k-选择的比例, 值通常在1—4之间波动, 较低的成熟度指数表明土壤养分呈富集状态, 主要是cp-1的线虫占优势, 或土壤环境处于胁迫状态, 主要表现为c-p 3—5的线虫数量减少[25]。PPI指数则指示植物寄生线虫r-选择和k-选择的比例, 反映植物寄生线虫的抗干扰能力和繁殖能力[15]。
采用线虫功能团指数来反映土壤食物网的变化。包括基础指数(Basal index, BI)、通路指数(Channel index, CI)、结构指数(Structure index, SI)和富集指数(Enrichment index, EI)[26], 公式如下:
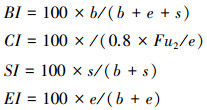
|
式中, b(basal)为食物网中的基础成分, 主要指Ba2和Fu2这两个类群, e(enrichment)代表食物网中的富集成分, 主要指Ba1和Fu2这两个类群, s(structure)代表食物网中的结构成分, 包括Ba3—Ba5、Fu3—Fu5、Pr3—Pr5类群;b、e和s的对应值分别为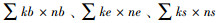
利用主成分分析(Principal component analysis, PCA)对各样地土壤线虫群落进行排序。利用独立样本t检验分析不同草地类型及季节间土壤线虫群落指标的差异性进行检验。对不符合正态分布的数据, 采用log(x+1)转换, 如仍不符合正态分布, 则采用Mann-Whitney U法进行检验。
合并7、9两月份土壤线虫群落的调查数据, 采用典范对应分析(Canonical correspondence analysis, CCA)分析燕麦地和天然草地土壤线虫群落结构与环境因子的关系。另外, 将7月和9月的密度及多样性等观测数据平均后, 采用多元回归分析土壤线虫群落各指标与环境因子的相关性。
数据处理和分析采用Excel 2010、IBM SPSS 22.0、Canoco 4.5软件。
2 结果与分析 2.1 土壤线虫群落组成两次采样共分离土壤线虫10179条, 隶属于2纲8目50科143属(表 1), 平均密度477条/100g干土。常见类群包括拟丽突属(Acrobeloides)等24属, 共占总采集数量的74.91%;稀有类群共119属, 占总数量的25.09%。群落组成中无优势类群。从营养类群的组成来看, 食细菌线虫有49属, 个体数量占总数的47.60%;植物寄生线虫50属, 占总数的33.02%;食真菌线虫14属, 占总数的11.93%;捕食-杂食线虫30属, 占总数的7.45%(表 1)。
| 类群 Group |
营养类群 Trophic group |
燕麦地Artificial oat grassland | 天然草地Nature alpine grassland | 总数 Total |
% | |||||||||||||
| 7月July | 9月September | 总数 Total |
% | 7月July | 9月September | 总数 Total |
% | |||||||||||
| 密度 Abundances/(条/100g干土) |
% | 密度 Abundances/(条/100g干土) |
% | 密度 Abundances/(条/100g干土) |
% | 密度 Abundances/(条/100g干土) |
% | |||||||||||
| 食细菌线虫Bacterivores (Ba) | ||||||||||||||||||
| 拟丽突属Acrobeloides | Ba2 | 515.00 | 13.23 | 153.92 | 5.01 | 668.92 | 9.61 | 123.64 | 4.82 | 101.33 | 5.27 | 224.98 | 5.02 | 893.89 | 7.81 | |||
| 棱咽属Prismatolaimus | Ba3 | 265.59 | 6.82 | 184.00 | 5.99 | 449.59 | 6.46 | 140.78 | 5.49 | 152.00 | 7.91 | 292.78 | 6.53 | 742.38 | 6.48 | |||
| 原杆属Protorhabditis | Ba1 | 358.82 | 9.22 | 156.00 | 5.08 | 514.82 | 7.39 | 62.28 | 2.43 | 67.58 | 3.52 | 129.86 | 2.90 | 644.68 | 5.63 | |||
| 绕线属Plectus | Ba2 | 170.24 | 4.37 | 102.20 | 3.33 | 272.44 | 3.91 | 29.65 | 1.16 | 93.33 | 4.86 | 122.98 | 2.74 | 395.42 | 3.45 | |||
| 角绕线属Ceratoplectus | Ba2 | 214.39 | 5.51 | 86.42 | 2.81 | 300.81 | 4.32 | 25.33 | 0.99 | 19.07 | 0.99 | 44.40 | 0.99 | 345.21 | 3.02 | |||
| 唇绕线属Chiloplectus | Ba2 | 100.95 | 2.59 | 94.05 | 3.06 | 195.00 | 2.80 | 52.11 | 2.03 | 46.72 | 2.43 | 98.83 | 2.20 | 293.83 | 2.57 | |||
| 巴氏属Bastiania | Ba3 | 132.84 | 3.41 | 78.69 | 2.56 | 211.53 | 3.04 | 65.13 | 2.54 | 8.63 | 0.45 | 73.75 | 1.64 | 285.28 | 2.49 | |||
| 杆咽属Rhabdolaimus | Ba3 | 78.34 | 2.01 | 104.06 | 3.39 | 182.40 | 2.62 | 53.60 | 2.09 | 1.76 | 0.09 | 55.35 | 1.23 | 237.76 | 2.08 | |||
| 威尔疣线属Wilsotylus | Ba2 | 75.94 | 1.95 | 20.22 | 0.66 | 96.17 | 1.38 | 95.49 | 3.72 | 32.67 | 1.70 | 128.15 | 2.86 | 224.32 | 1.96 | |||
| 简咽属Deontolaimus | Ba2 | 181.38 | 4.66 | 17.35 | 0.56 | 198.73 | 2.85 | — | — | 9.52 | 0.50 | 9.52 | 0.21 | 208.24 | 1.82 | |||
| 头叶属Cephalobus | Ba2 | 48.34 | 1.24 | 67.10 | 2.18 | 115.45 | 1.66 | — | — | 4.70 | 0.24 | 4.70 | 0.10 | 120.15 | 1.05 | |||
| 单宫属Monhystera | Ba2 | — | — | 18.89 | 0.61 | 18.89 | 0.27 | 17.65 | 0.69 | 51.25 | 2.67 | 68.90 | 1.54 | 87.79 | 0.77 | |||
| 柱咽属Cylindrolaimus | Ba3 | 55.86 | 1.44 | 5.86 | 0.19 | 61.72 | 0.89 | 16.18 | 0.63 | 4.40 | 0.23 | 20.58 | 0.46 | 82.30 | 0.72 | |||
| 拟绕线属Anaplectus | Ba2 | 37.79 | 0.97 | 37.06 | 1.21 | 74.85 | 1.07 | — | — | 2.99 | 0.16 | 2.99 | 0.07 | 77.83 | 0.68 | |||
| 角头属Tylocephalus | Ba2 | — | — | 7.97 | 0.26 | 7.97 | 0.11 | 25.31 | 0.99 | 32.79 | 1.71 | 58.10 | 1.30 | 66.07 | 0.58 | |||
| 连胃属Chronogaster | Ba2 | — | — | 64.60 | 2.10 | 64.60 | 0.93 | — | — | — | — | — | — | 64.60 | 0.56 | |||
| 真单宫属Eumonhystera | Ba2 | — | — | 40.59 | 1.32 | 40.59 | 0.58 | — | — | 16.32 | 0.85 | 16.32 | 0.36 | 56.91 | 0.50 | |||
| 双胃属Diplogaster | Ba1 | 32.24 | 0.83 | 24.49 | 0.80 | 56.72 | 0.81 | — | — | — | — | — | — | 56.72 | 0.50 | |||
| 短腔属Brevibucca | Ba1 | — | — | 18.46 | 0.60 | 18.46 | 0.27 | 5.96 | 0.23 | 32.07 | 1.67 | 38.03 | 0.85 | 56.49 | 0.49 | |||
| 齿咽属Odotopharynx | Ba1 | — | — | 26.67 | 0.87 | 26.67 | 0.38 | 23.45 | 0.91 | 2.04 | 0.11 | 25.49 | 0.57 | 52.16 | 0.46 | |||
| 板环属Placodira | Ba2 | 40.20 | 1.03 | — | — | 40.20 | 0.58 | — | — | — | — | — | — | 40.20 | 0.35 | |||
| 无咽属Alaimus | Ba4 | 23.52 | 0.60 | 14.51 | 0.47 | 38.03 | 0.55 | — | — | — | — | — | — | 38.03 | 0.33 | |||
| 伪双胃属 Pseudodiplogasteroides |
Ba1 | — | — | 25.54 | 0.83 | 25.54 | 0.37 | 5.57 | 0.22 | 5.65 | 0.29 | 11.22 | 0.25 | 36.76 | 0.32 | |||
| 真头叶属Eucephalobus | Ba2 | — | — | 9.31 | 0.30 | 9.31 | 0.13 | 13.57 | 0.53 | 4.18 | 0.22 | 17.75 | 0.40 | 27.06 | 0.24 | |||
| 隐咽属Aphanolaimus | Ba2 | — | — | 15.99 | 0.52 | 15.99 | 0.23 | 10.62 | 0.41 | — | — | 10.62 | 0.24 | 26.60 | 0.23 | |||
| 丽突属Acrobeles | Ba2 | 12.60 | 0.32 | 8.25 | 0.27 | 20.85 | 0.30 | — | — | 1.45 | 0.08 | 1.45 | 0.03 | 22.30 | 0.19 | |||
| 明杆属Rhabditophanes | Ba1 | — | — | 21.76 | 0.71 | 21.76 | 0.31 | — | — | — | — | — | — | 21.76 | 0.19 | |||
| 后双胃属Metadiplogaster | Ba1 | 21.49 | 0.55 | — | — | 21.49 | 0.31 | — | — | — | — | — | — | 21.49 | 0.19 | |||
| 小杆属Rhabditis | Ba1 | — | — | 2.97 | 0.10 | 2.97 | 0.04 | — | — | 17.32 | 0.90 | 17.32 | 0.39 | 20.30 | 0.18 | |||
| 浅腔属Panagrellus | Ba1 | — | — | 17.92 | 0.58 | 17.92 | 0.26 | — | — | 1.93 | 0.10 | 1.93 | 0.04 | 19.86 | 0.17 | |||
| 盆咽属Panagrolaimus | Ba1 | 13.33 | 0.34 | — | — | 13.33 | 0.19 | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 19.30 | 0.17 | |||
| 微咽属Microlaimus | Ba2 | — | — | 17.56 | 0.57 | 17.56 | 0.25 | — | — | — | — | — | — | 17.56 | 0.15 | |||
| 活跃属Greenenema | Ba3 | 10.92 | 0.28 | — | — | 10.92 | 0.16 | — | — | 3.32 | 0.17 | 3.32 | 0.07 | 14.24 | 0.12 | |||
| 异咽属Poikilolaimus | Ba1 | — | — | — | — | — | — | 13.57 | 0.53 | — | — | 13.57 | 0.30 | 13.57 | 0.12 | |||
| 小单宫属Monhystrella | Ba2 | — | — | 13.38 | 0.44 | 13.38 | 0.19 | — | — | — | — | — | — | 13.38 | 0.12 | |||
| 异双胃属Diplogasteriana | Ba1 | 13.33 | 0.34 | — | — | 13.33 | 0.19 | — | — | — | — | — | — | 13.33 | 0.12 | |||
| 管咽属Aulolaimus | Ba3 | — | — | — | — | — | — | 12.24 | 0.48 | — | — | 12.24 | 0.27 | 12.24 | 0.11 | |||
| 无齿属Anonchus | Ba2 | — | — | 11.59 | 0.38 | 11.59 | 0.17 | — | — | — | — | — | — | 11.59 | 0.10 | |||
| 杯咽属Cytholaimus | Ba2 | — | — | 6.08 | 0.20 | 6.08 | 0.09 | — | — | 5.33 | 0.28 | 5.33 | 0.12 | 11.41 | 0.10 | |||
| 拟鳗线属Anguilluloides | Ba1 | — | — | 2.97 | 0.10 | 2.97 | 0.04 | — | — | 8.37 | 0.44 | 8.37 | 0.19 | 11.34 | 0.10 | |||
| 瓣唇属Panagrobelus | Ba1 | 10.92 | 0.28 | — | — | 10.92 | 0.16 | — | — | — | — | — | — | 10.92 | 0.10 | |||
| 中杆属Mesorhabditis | Ba1 | — | — | 2.89 | 0.09 | 2.89 | 0.04 | — | — | 4.67 | 0.24 | 4.67 | 0.10 | 7.56 | 0.07 | |||
| 扭沟属Plectonchus | Ba1 | — | — | — | — | — | — | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 5.96 | 0.05 | |||
| 圆顶属Domorganus | Ba2 | — | — | — | — | — | — | 5.57 | 0.22 | — | — | 5.57 | 0.12 | 5.57 | 0.05 | |||
| 板唇属Chiloplacus | Ba2 | — | — | 5.29 | 0.17 | 5.29 | 0.08 | — | — | — | — | — | — | 5.29 | 0.05 | |||
| 细咽属Tylopharynx | Ba2 | — | — | 2.97 | 0.10 | 2.97 | 0.04 | — | — | — | — | — | — | 2.97 | 0.03 | |||
| 原细咽属Protylopharynx | Ba2 | — | — | 2.97 | 0.10 | 2.97 | 0.04 | — | — | — | — | — | — | 2.97 | 0.03 | |||
| 三等齿属Pelodera | Ba1 | — | — | 2.68 | 0.09 | 2.68 | 0.04 | — | — | — | — | — | — | 2.68 | 0.02 | |||
| 伪杆咽属Rhabdontolaimus | Ba1 | — | — | — | — | — | — | — | — | 1.45 | 0.08 | 1.45 | 0.03 | 1.45 | 0.01 | |||
| 食真菌线虫Fungivores (Fu) | ||||||||||||||||||
| 滑刃属Aphelenchoides | Fu2 | 410.14 | 10.54 | 114.08 | 3.71 | 524.23 | 7.53 | 49.18 | 1.92 | 60.50 | 3.15 | 109.68 | 2.45 | 633.90 | 5.54 | |||
| 真滑刃属Aphelenchus | Fu2 | 116.71 | 3.00 | 89.85 | 2.92 | 206.56 | 2.97 | 30.28 | 1.18 | 13.09 | 0.68 | 43.38 | 0.97 | 249.94 | 2.18 | |||
| 垫咽属Tylencholaimus | Fu2 | — | — | 107.26 | 3.49 | 107.26 | 1.54 | 55.40 | 2.16 | 25.39 | 1.32 | 80.79 | 1.80 | 188.04 | 1.64 | |||
| 散香属Beleodorus | Fu2 | 69.28 | 1.78 | 42.00 | 1.37 | 111.28 | 1.60 | — | — | — | — | — | — | 111.28 | 0.97 | |||
| 茎属Ditylenchus | Fu2 | 22.60 | 0.58 | 29.59 | 0.96 | 52.19 | 0.75 | 33.16 | 1.29 | 23.27 | 1.21 | 56.42 | 1.26 | 108.61 | 0.95 | |||
| 粒属Anguina | Fu2 | — | — | 5.25 | 0.17 | 5.25 | 0.08 | 13.57 | 0.53 | — | — | 13.57 | 0.30 | 18.82 | 0.16 | |||
| 异球属Stictylus | Fu2 | 10.75 | 0.28 | — | — | 10.75 | 0.15 | — | — | 3.87 | 0.20 | 3.87 | 0.09 | 14.61 | 0.13 | |||
| 柄端球属Paurodontus | Fu2 | 10.92 | 0.28 | — | — | 10.92 | 0.16 | — | — | — | — | — | — | 10.92 | 0.10 | |||
| 原细齿属Proleptonchus | Fu2 | — | — | 8.73 | 0.28 | 8.73 | 0.13 | — | — | — | — | — | — | 8.73 | 0.08 | |||
| 伪垫刃属Nothotylenchus | Fu2 | — | — | 6.00 | 0.20 | 6.00 | 0.09 | — | — | — | — | — | — | 6.00 | 0.05 | |||
| 细杆滑刃属 Rhadinaphelenchus |
Fu4 | — | — | — | — | — | — | 5.57 | 0.22 | — | — | 5.57 | 0.12 | 5.57 | 0.05 | |||
| 细齿属Leptonchus | Fu4 | — | — | 3.25 | 0.11 | 3.25 | 0.05 | — | — | — | — | — | — | 3.25 | 0.03 | |||
| 亚粒属Subanguina | Fu4 | — | — | 2.89 | 0.09 | 2.89 | 0.04 | — | — | — | — | — | — | 2.89 | 0.03 | |||
| 短矛属Tylencholaimellus | Fu4 | — | — | 2.78 | 0.09 | 2.78 | 0.04 | — | — | — | — | — | — | 2.78 | 0.02 | |||
| 植物寄生线虫Plantparasites (Pl) | ||||||||||||||||||
| 丝尾垫刃属Filenchus | Pl2 | 334.04 | 8.58 | 232.67 | 7.57 | 566.70 | 8.14 | 182.35 | 7.11 | 134.25 | 6.99 | 316.59 | 7.06 | 883.30 | 7.71 | |||
| 垫刃属Tylenchus | Pl2 | 69.65 | 1.79 | 259.90 | 8.46 | 329.54 | 4.73 | 187.79 | 7.32 | 159.33 | 8.29 | 347.12 | 7.74 | 676.67 | 5.91 | |||
| 剑尾垫刃属Malenchus | Pl2 | 80.41 | 2.07 | 32.55 | 1.06 | 112.97 | 1.62 | 104.77 | 4.09 | 112.25 | 5.84 | 217.02 | 4.84 | 329.99 | 2.88 | |||
| 长针属Longidorus | Pl2 | 51.69 | 1.33 | 52.17 | 1.70 | 103.85 | 1.49 | 56.55 | 2.21 | 97.33 | 5.07 | 153.89 | 3.43 | 257.74 | 2.25 | |||
| 螺旋属Helicotylenchus | Pl2 | 21.85 | 0.56 | 39.06 | 1.27 | 60.91 | 0.87 | 72.63 | 2.83 | 78.26 | 4.07 | 150.89 | 3.36 | 211.80 | 1.85 | |||
| 大矛属Enchodelum | Pl2 | 76.89 | 1.98 | 27.17 | 0.88 | 104.06 | 1.49 | 58.51 | 2.28 | 8.81 | 0.46 | 67.32 | 1.50 | 171.38 | 1.50 | |||
| 盘旋属Rotylenchus | Pl2 | — | — | 43.53 | 1.42 | 43.53 | 0.63 | 19.76 | 0.77 | 90.77 | 4.72 | 110.54 | 2.46 | 154.07 | 1.35 | |||
| 细纹垫刃属Lelenchus | Pl2 | 10.75 | 0.28 | 37.49 | 1.22 | 48.24 | 0.69 | 31.46 | 1.23 | 52.69 | 2.74 | 84.15 | 1.88 | 132.39 | 1.16 | |||
| 短体属Pratylenchus | Pl2 | 10.92 | 0.28 | 13.35 | 0.43 | 24.27 | 0.35 | 69.11 | 2.70 | 24.98 | 1.30 | 94.08 | 2.10 | 118.35 | 1.03 | |||
| 矮化属Tylenchorynchus | Pl2 | 12.60 | 0.32 | 13.90 | 0.45 | 26.50 | 0.38 | 63.25 | 2.47 | 8.22 | 0.43 | 71.46 | 1.59 | 97.96 | 0.86 | |||
| 新锥属Neodolichodorus | Pl2 | — | — | — | — | — | — | 73.81 | 2.88 | — | — | 73.81 | 1.65 | 73.81 | 0.64 | |||
| 短锥属Brachydorus | Pl2 | — | — | 12.02 | 0.39 | 12.02 | 0.17 | 57.28 | 2.23 | 2.94 | 0.15 | 60.23 | 1.34 | 72.25 | 0.63 | |||
| 拟长针属Paralongidorus | Pl3 | — | — | 3.10 | 0.10 | 3.10 | 0.04 | 67.77 | 2.64 | — | — | 67.77 | 1.51 | 70.87 | 0.62 | |||
| 穿孔属Radopholus | Pl3 | 21.85 | 0.56 | 12.70 | 0.41 | 34.55 | 0.50 | 19.76 | 0.77 | 15.35 | 0.80 | 35.11 | 0.78 | 69.66 | 0.61 | |||
| 潜根属Hirschmanniella | Pl3 | — | — | 19.32 | 0.63 | 19.32 | 0.28 | — | — | 28.78 | 1.50 | 28.78 | 0.64 | 48.10 | 0.42 | |||
| 拟剑针属Paraxiphidorus | Pl3 | — | — | 5.18 | 0.17 | 5.18 | 0.07 | 26.80 | 1.05 | — | — | 26.80 | 0.60 | 31.98 | 0.28 | |||
| 杆垫刃属Rhabdotylenchus | Pl3 | — | — | 4.09 | 0.13 | 4.09 | 0.06 | 13.57 | 0.53 | 14.00 | 0.73 | 27.57 | 0.61 | 31.66 | 0.28 | |||
| 头垫刃属Cephalenchus | Pl3 | — | — | 10.36 | 0.34 | 10.36 | 0.15 | 16.18 | 0.63 | 4.36 | 0.23 | 20.54 | 0.46 | 30.91 | 0.27 | |||
| 短体长针属Longidorella | Pl3 | — | — | — | — | — | — | — | — | 30.00 | 1.56 | 30.00 | 0.67 | 30.00 | 0.26 | |||
| 新平滑垫刃属Neopsilenchus | Pl3 | — | — | 25.96 | 0.84 | 25.96 | 0.37 | — | — | 2.05 | 0.11 | 2.05 | 0.05 | 28.01 | 0.24 | |||
| 默林属Merlinius | Pl3 | — | — | 1.50 | 0.05 | 1.50 | 0.02 | 17.89 | 0.70 | 4.69 | 0.24 | 22.58 | 0.50 | 24.08 | 0.21 | |||
| 类短体属Pratylenchoides | Pl3 | — | — | — | — | — | — | 21.23 | 0.83 | 2.67 | 0.14 | 23.90 | 0.53 | 23.90 | 0.21 | |||
| 叉针属Boleodorus | Pl3 | — | — | 2.73 | 0.09 | 2.73 | 0.04 | 15.61 | 0.61 | — | — | 15.61 | 0.35 | 18.34 | 0.16 | |||
| 大默林属Amplimerlinius | Pl3 | — | — | — | — | — | — | 17.89 | 0.70 | — | — | 17.89 | 0.40 | 17.89 | 0.16 | |||
| 巴兹尔垫刃属Basiria | Pl3 | 10.92 | 0.28 | 5.63 | 0.18 | 16.55 | 0.24 | — | — | — | — | — | — | 16.55 | 0.14 | |||
| 沟线虫属Ogma | Pl3 | — | — | — | — | — | — | — | — | 15.37 | 0.80 | 15.37 | 0.34 | 15.37 | 0.13 | |||
| 毛刺属Trichodorus | Pl3 | — | — | — | — | — | — | 11.14 | 0.43 | 2.86 | 0.15 | 14.00 | 0.31 | 14.00 | 0.12 | |||
| 剑属Xiphinema | Pl3 | — | — | 6.68 | 0.22 | 6.68 | 0.10 | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 12.64 | 0.11 | |||
| 肾状属Rotylenchulus | Pl3 | — | — | — | — | — | — | 11.93 | 0.47 | — | — | 11.93 | 0.27 | 11.93 | 0.10 | |||
| 异毛刺属Allotrichodorus | Pl3 | — | — | — | — | — | — | 11.14 | 0.43 | — | — | 11.14 | 0.25 | 11.14 | 0.10 | |||
| 拟盘旋属Pararotylenchus | Pl3 | — | — | — | — | — | — | 5.96 | 0.23 | 3.28 | 0.17 | 9.24 | 0.21 | 9.24 | 0.08 | |||
| 南球属Antarctylus | Pl3 | — | — | 2.73 | 0.09 | 2.73 | 0.04 | — | — | 5.67 | 0.29 | 5.67 | 0.13 | 8.40 | 0.07 | |||
| 米卡垫刃属Miculenchus | Pl3 | — | — | 7.96 | 0.26 | 7.96 | 0.11 | — | — | — | — | — | — | 7.96 | 0.07 | |||
| 裸矛属Psilenchus | Pl3 | — | — | — | — | — | — | — | — | 7.57 | 0.39 | 7.57 | 0.17 | 7.57 | 0.07 | |||
| 刺属Belonolaimus | Pl3 | — | — | 3.25 | 0.11 | 3.25 | 0.05 | — | — | 2.86 | 0.15 | 2.86 | 0.06 | 6.11 | 0.05 | |||
| 塞内加尔线虫属 Senegalonema |
Pl3 | — | — | — | — | — | — | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 5.96 | 0.05 | |||
| 无侧尾腺属 Aphasmatylenchus |
Pl3 | — | — | — | — | — | — | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 5.96 | 0.05 | |||
| 颚针属Belondira | Pl3 | — | — | — | — | — | — | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 5.96 | 0.05 | |||
| 接合垫刃属Zygotylenchus | Pl4 | — | — | — | — | — | — | — | — | 4.18 | 0.22 | 4.18 | 0.09 | 4.18 | 0.04 | |||
| 单毛刺属Monotrichodorus | Pl4 | — | — | — | — | — | — | 4.08 | 0.16 | — | — | 4.08 | 0.09 | 4.08 | 0.04 | |||
| 拟毛刺属Paratrichodorus | Pl4 | — | — | — | — | — | — | 4.08 | 0.16 | — | — | 4.08 | 0.09 | 4.08 | 0.04 | |||
| 盾状线虫属Scutellonema | Pl4 | — | — | — | — | — | — | — | — | 3.52 | 0.18 | 3.52 | 0.08 | 3.52 | 0.03 | |||
| 环属Criconema | Pl4 | — | — | 3.04 | 0.10 | 3.04 | 0.04 | — | — | — | — | — | — | 3.04 | 0.03 | |||
| 鞘属Hemicycliophora | Pl4 | — | — | 3.04 | 0.10 | 3.04 | 0.04 | — | — | — | — | — | — | 3.04 | 0.03 | |||
| 野外垫刃属Aglenchus | Pl5 | — | — | 2.78 | 0.09 | 2.78 | 0.04 | — | — | — | — | — | — | 2.78 | 0.02 | |||
| 纽带属Hoplolaimus | Pl5 | — | — | 2.63 | 0.09 | 2.63 | 0.04 | — | — | — | — | — | — | 2.63 | 0.02 | |||
| 秆矛属Carphodorus | Pl5 | — | — | 2.63 | 0.09 | 2.63 | 0.04 | — | — | — | — | — | — | 2.63 | 0.02 | |||
| 类长针属Longidoroides | Pl5 | — | — | 2.63 | 0.09 | 2.63 | 0.04 | — | — | — | — | — | — | 2.63 | 0.02 | |||
| 卡卢斯属Caloosia | Pl5 | — | — | 2.56 | 0.08 | 2.56 | 0.04 | — | — | — | — | — | — | 2.56 | 0.02 | |||
| 居中属Geocenamus | Pl5 | — | — | — | — | — | — | — | — | 1.97 | 0.10 | 1.97 | 0.04 | 1.97 | 0.02 | |||
| 捕食杂食线虫Predators omnivores (Pr) | ||||||||||||||||||
| 色矛属Chromadorita | Pr3 | 21.49 | 0.55 | 77.64 | 2.53 | 99.13 | 1.42 | 66.81 | 2.61 | 12.17 | 0.63 | 78.98 | 1.76 | 178.11 | 1.56 | |||
| 类管咽属Aulolaimoides | Pr3 | 10.92 | 0.28 | 2.56 | 0.08 | 13.49 | 0.19 | 81.41 | 3.18 | — | — | 81.41 | 1.82 | 94.90 | 0.83 | |||
| 托布利属Tobrilus | Pr3 | 24.46 | 0.63 | 33.96 | 1.11 | 58.42 | 0.84 | 33.73 | 1.32 | — | — | 33.73 | 0.75 | 92.15 | 0.80 | |||
| 单棘属Mononchulus | Pr3 | 13.33 | 0.34 | 20.69 | 0.67 | 34.03 | 0.49 | — | — | 23.33 | 1.21 | 23.33 | 0.52 | 57.36 | 0.50 | |||
| 基齿属Iotonchus | Pr3 | 10.92 | 0.28 | 2.63 | 0.09 | 13.55 | 0.19 | 31.92 | 1.25 | 8.23 | 0.43 | 40.15 | 0.90 | 53.70 | 0.47 | |||
| 矛线属Dorylaimus | Pr3 | 10.92 | 0.28 | 15.97 | 0.52 | 26.89 | 0.39 | — | — | 7.84 | 0.41 | 7.84 | 0.17 | 34.73 | 0.30 | |||
| 单色矛属Monochromadora | Pr3 | — | — | 12.63 | 0.41 | 12.63 | 0.18 | — | — | 19.03 | 0.99 | 19.03 | 0.42 | 31.65 | 0.28 | |||
| 下齿属Hypodontolaimus | Pr4 | — | — | 8.89 | 0.29 | 8.89 | 0.13 | 20.40 | 0.80 | 1.93 | 0.10 | 22.33 | 0.50 | 31.23 | 0.27 | |||
| 真矛线属Eudorylaimus | Pr4 | — | — | 2.63 | 0.09 | 2.63 | 0.04 | 10.62 | 0.41 | 17.58 | 0.91 | 28.19 | 0.63 | 30.82 | 0.27 | |||
| 伊龙属Ironus | Pr4 | — | — | 12.25 | 0.40 | 12.25 | 0.18 | 11.53 | 0.45 | — | — | 11.53 | 0.26 | 23.79 | 0.21 | |||
| 等齿属Miconchus | Pr4 | — | — | — | — | — | — | 23.45 | 0.91 | — | — | 23.45 | 0.52 | 23.45 | 0.20 | |||
| 前矛线属Prodorylaimus | Pr4 | — | — | 12.42 | 0.40 | 12.42 | 0.18 | — | — | 10.16 | 0.53 | 10.16 | 0.23 | 22.57 | 0.20 | |||
| 盘腔属Discomyctus | Pr4 | 10.92 | 0.28 | 5.26 | 0.17 | 16.18 | 0.23 | — | — | — | — | — | — | 16.18 | 0.14 | |||
| 小剑属Xiphinemella | Pr4 | — | — | 3.10 | 0.10 | 3.10 | 0.04 | 5.57 | 0.22 | 6.89 | 0.36 | 12.46 | 0.28 | 15.55 | 0.14 | |||
| 盘咽属Discolaimus | Pr4 | — | — | 8.00 | 0.26 | 8.00 | 0.11 | 5.57 | 0.22 | 1.36 | 0.07 | 6.93 | 0.15 | 14.93 | 0.13 | |||
| 孔咽属Aporcelaimus | Pr4 | — | — | 12.27 | 0.40 | 12.27 | 0.18 | — | — | — | — | — | — | 12.27 | 0.11 | |||
| 拱唇属Labronema | Pr4 | — | — | 2.97 | 0.10 | 2.97 | 0.04 | — | — | 8.79 | 0.46 | 8.79 | 0.20 | 11.76 | 0.10 | |||
| 拟桑尼属Thorneella | Pr4 | — | — | — | — | — | — | — | — | 11.44 | 0.60 | 11.44 | 0.26 | 11.44 | 0.10 | |||
| 拟矛线属Dorylaimoides | Pr4 | 10.92 | 0.28 | — | — | 10.92 | 0.16 | — | — | — | — | — | — | 10.92 | 0.10 | |||
| 小穿咽属Nygolaimellus | Pr4 | 10.92 | 0.28 | — | — | 10.92 | 0.16 | — | — | — | — | — | — | 10.92 | 0.10 | |||
| 锉齿属Mylonchulus | Pr4 | 10.75 | 0.28 | — | — | 10.75 | 0.15 | — | — | — | — | — | — | 10.75 | 0.09 | |||
| 中矛线属Mesodorylaimus | Pr4 | — | — | 3.25 | 0.11 | 3.25 | 0.05 | — | — | 7.47 | 0.39 | 7.47 | 0.17 | 10.73 | 0.09 | |||
| 桑尼属Thornia | Pr5 | — | — | 10.51 | 0.34 | 10.51 | 0.15 | — | — | — | — | — | — | 10.51 | 0.09 | |||
| 类矛线属Amphidorylaimus | Pr5 | — | — | 2.78 | 0.09 | 2.78 | 0.04 | 5.96 | 0.23 | — | — | 5.96 | 0.13 | 8.75 | 0.08 | |||
| 金线属Chrysonema | Pr5 | — | — | 5.20 | 0.17 | 5.20 | 0.07 | — | — | 3.52 | 0.18 | 3.52 | 0.08 | 8.72 | 0.08 | |||
| 异色矛属Achromadora | Pr5 | — | — | 8.35 | 0.27 | 8.35 | 0.12 | — | — | — | — | — | — | 8.35 | 0.07 | |||
| 三孔属Tripyla | Pr5 | — | — | 2.97 | 0.10 | 2.97 | 0.04 | — | — | 3.55 | 0.18 | 3.55 | 0.08 | 6.53 | 0.06 | |||
| 角咽属Actinolaimus | Pr5 | — | — | — | — | — | — | 4.08 | 0.16 | — | — | 4.08 | 0.09 | 4.08 | 0.04 | |||
| 原色矛属Prochromadora | Pr5 | — | — | 3.53 | 0.12 | 3.53 | 0.05 | — | — | — | — | — | — | 3.53 | 0.03 | |||
| 单齿属Mononchus | Pr5 | — | — | 2.59 | 0.08 | 2.59 | 0.04 | — | — | — | — | — | — | 2.59 | 0.02 | |||
| 食细菌线虫密度 Ba abundances |
2414.04 | 62.03 | 1493.23 | 48.60 | 3, 907.27 | 56.11 | 809.63 | 31.58 | 732.84 | 38.14 | 1542.47 | 34.39 | 4950.95 | 46.19 | ||||
| 食真菌线虫密度 Fu abundances |
640.40 | 16.46 | 411.70 | 13.40 | 1, 052.10 | 15.11 | 187.16 | 7.30 | 126.11 | 6.56 | 313.27 | 6.98 | 1220.48 | 11.39 | ||||
| 植物寄生线虫密度 Pl abundances |
701.56 | 18.03 | 894.32 | 29.11 | 1, 595.87 | 22.92 | 1266.17 | 49.38 | 919.00 | 47.83 | 2185.17 | 48.72 | 3703.84 | 34.56 | ||||
| 捕食杂食线虫密度 Pr abundances |
135.56 | 3.48 | 273.07 | 8.89 | 408.63 | 5.87 | 301.05 | 11.74 | 143.28 | 7.46 | 444.33 | 9.91 | 842.83 | 7.86 | ||||
| 总密度 Total abundances |
3892.56 | 100 | 3072.32 | 100 | 6369.88 | 100 | 2564.01 | 100 | 1921.23 | 100 | 4485.24 | 100 | 11449.12 | 100.00 | ||||
| 总类群数 Total taxonomic richness |
100 | 105 | 118 | 77 | 103 | 103 | 143 | |||||||||||
燕麦地共采集6015条, 隶属于2纲8目116属, 平均密度508.3条/100g干土, 常见类群包括拟丽突属等22属, 占总数量的79.64%;盘旋属(Rotylenchus)等94属为稀有类群, 占20.36%;天然草地共采集线虫4164条, 隶属于2纲8目103属, 平均密度373.8条/100g干土;垫刃属(Tylenchus)等28属为常见类群, 占总数量的77.50%;角绕线属(Ceratoplectus)等75属为稀有类群, 占22.50%;其中两种草地类型均无优势类群(表 1)。从营养类群的组成来看, 在燕麦地和天然草地中优势营养类群均为食细菌线虫和植物寄生线虫, 分别占总数的56.11%、34.39%和22.92%、48.72%;食真菌线虫仅在燕麦地中为优势营养类群, 占总数的15.11%(表 1)。
2.2 土壤线虫群落结构差异土壤线虫群落的主成分分析(PCA)结果如图 1。7月份和9月份的前两个排序轴累计贡献率分别达52.4%和52.6%, 且两个月份燕麦地与天然草地均被PC1和PC2明显分开, 说明两种草地类型的土壤线虫群落结构差异明显。7月份, 拟丽突属和简咽属(Deontolaimus)对PC1的投影较大, 表明PC1主要反映这两个类群的变化;同理, 排序轴PC2主要反映原杆属(Protorhabditis)和板环属(Placodira)等的数量变化。9月份, PC1轴主要反映丝尾垫刃属(Filenchus)和垫刃属等类群的数量变化, PC2轴主要反映盘旋属和色矛属(Chromadorita)等类群的数量变化。

|
| 图 1 燕麦地和天然草地土壤线虫群落排序 Fig. 1 Principal component analysis on the soil nematode communities in artificial oat grassland and nature alpine grassland |
燕麦地土壤线虫群落的密度、类群数、香农多样性指数和Simpson优势度指数均高于天然草地(图 2), 但统计分析结果显示仅群落密度差异性显著(P<0.05)。

|
| 图 2 燕麦地和天然草地土壤线虫群落密度及多样性(平均值±标准误) Fig. 2 Abundances and diversity of the soil nematode communities in artificial oat grassland and nature alpine grassland (Mean±SE) |
在月份变化方面, 7月份两种草地类型的土壤线虫群落密度和Simpson优势度指数均高于9月份, 而类群数、香农多样性指数则相反(图 2)。统计分析结果显示, 季节变化对燕麦地土壤线虫群落的类群数、香农多样性指数、Simpson优势度指数均有显著影响(P<0.01; P<0.001; P<0.01), 但仅对天然草地的类群数和香农多样性指数有显著影响(P<0.01; P<0.05)。
2.4 土壤线虫群落营养结构差异从7、9月份的整体情况来看, 燕麦地的食细菌和食真菌线虫密度显著高于天然草地(P<0.05; P<0.001)(图 3)。而植物寄生和捕食-杂食性线虫密度与天然草地间无显著差异(P>0.05)(图 3)。此外, 燕麦地7月份的食细菌和食真菌密度显著高于9月(P<0.001; P<0.05)(图 3), 而燕麦地的植物寄生和捕食杂食线虫和天然草地中各营养类群的线虫密度在两月份间差异均不显著(P>0.05)(图 3)。

|
| 图 3 燕麦地与天然草地土壤线虫各营养类群的密度(平均值±标准误) Fig. 3 Abundances of the four trophic groups of nematodes in artificial oat grassland and nature alpine grassland (Mean±SE) |
在计算的6个生态指数中, 燕麦地EI、BI、CI和MI值大于天然草地;SI和PPI值则相反。统计分析结果表明仅MI和PPI值在两种样地间差异显著(P<0.01; P<0.05), 其他生态指数均无显著差异(P>0.05)(图 4)。此外, 统计分析结果显示, 两种草地类型的各生态指数季节间均无显著差异(P>0.05)(图 4)。

|
| 图 4 不同草地类型土壤线虫群落生态指数(平均值±标准误) Fig. 4 Ecological indices of soil nematode communities in artificial oat grassland and nature alpine grassland (Mean±SE) |
燕麦地的土壤pH和含水量显著高于天然草地(P<0.01);土壤有机质、全氮和速效氮含量显著低于天然草地(P<0.01; P<0.01; P<0.05)。土壤全磷、速效磷、全钾及速效钾的含量在燕麦地和天然草地间差异不显著(表 2)。
| 土壤性质Soil properties | 天然草地NG | 燕麦地OG | t | P |
| pH | 4.62±0.05 | 5.01±0.06 | 5.05 | <0.01 |
| 土壤含水量WC/(%) | 29.4±0.42 | 35.05±0.87 | 5.83 | <0.01 |
| 有机质OM/(g/kg) | 48.33±1.15 | 43.00±0.21 | 4.57 | <0.01 |
| 全氮TN/(g/kg) | 3.11±0.07 | 2.82±0.01 | 4.34 | <0.01 |
| 全磷TP/(g/kg) | 0.74±0.01 | 0.72±0.01 | 1.10 | >0.05 |
| 全钾TK/(g/kg) | 14.50±0.25 | 14.89±0.23 | 1.15 | >0.05 |
| 速效磷AP/(mg/kg) | 2.90±0.32 | 5.86±1.92 | 1.52 | >0.05 |
| 速效钾AK/(mg/kg) | 103.40±4.60 | 89.60±10.45 | 1.21 | >0.05 |
| 速效氮AN/(mg/kg) | 114.90±7.49 | 75.26±14.34 | 2.45 | <0.05 |
| NG:天然草地Nature alpine grassland;OG:燕麦地Artificial oat grassland;AP:速效磷Available P;AK:速效钾Available K;AN:速效氮Available N;TP:全磷Total P;TK:全钾Total K;TK:全氮Total N;WC:土壤含水量Water content;OM:有机质Organic material | ||||
对各样地土壤线虫群落与环境因子进行CCA排序, 第一排序轴和第二排序轴对土壤线虫群落和土壤环境因子关系的解释率分别为22.8%和17.0%(图 5), 且蒙特卡洛检验结果显示第一排序轴解释量具有显著性(P<0.05)。由图 5可知, 第一轴与全氮(r=0.91, P<0.01)、有机质(r=0.90, P<0.01)、速效氮(r=0.70, P<0.05)呈显著正相关关系, 与pH(r=-0.58, P<0.01)和含水量(r=-0.62, P<0.05)显著负相关;第二轴与速效钾(r=0.66, P>0.05)呈正相关、速效磷(r=-0.50, P>0.05)呈负相关。此外, 燕麦地土壤线虫群落主要与土壤pH、含水量、速效磷和全钾含量呈正相关, 与有机质、全氮、速效氮、全磷、速效钾负相关;天然草地则相反。

|
| 图 5 土壤线虫群落结构与环境因子的典范对应分析 Fig. 5 Canonical correspondence analysis on soil nematode communities and environmental factors AP:速效磷Available P;AK:速效钾Available K;AN:速效氮Available N;TP:全磷Total P;TK:全钾Total K;TK:全氮Total N;WC:土壤含水量Water content;OM:有机质Organic material |
多元回归分析(表 3)表明, 在测量的8个土壤环境因子中, 有6个因子与土壤线虫群落具有显著相关性。其中香农多样性指数与速效磷呈显著正相关(P<0.05), 群落密度和类群数分别与速效氮和有机质呈显著负相关(P<0.05);食细菌线虫密度与pH有显著正相关(P<0.01), 食真菌和植物寄生线虫密度则分别与土壤含水量呈显著正、负相关(P<0.05)。在土壤线虫的生态指数中, 自由生活线虫成熟度指数(MI)、植物线虫成熟度指数(PPI)、结构指数(SI)均与pH有显著正相关(P<0.05; P<0.01; P<0.05), 通路指数(CI)则与pH呈显著负相关(P<0.05);此外, 基础指数(BI)与速效钾(AK)呈显著正相关(P<0.01), 与速效磷(AP)和pH呈显著负相关(P<0.01)。
| 项目 Item |
pH | 土壤含水量 Soil water content |
有机质 Organic material |
速效磷 Available P |
速效钾 Available K |
速效氮 Available N |
| 香农多样性指数Shannon-Weiner index (H′) | — | — | — | 0.62* | — | — |
| 类群数Taxonomic richness | — | — | -0.63* | — | — | — |
| 群落密度Community abundances | — | — | — | — | — | -0.67* |
| 食细菌线虫密度Bacterivore abundances | 0.77** | — | — | — | — | — |
| 食真菌线虫密度Fungivore abundances | — | 0.78* | — | — | — | — |
| 植物寄生线虫密度Plant-parasite abundances | — | -0.67* | — | — | — | — |
| 结构指数Structure index (SI) | 0.60* | — | — | — | — | — |
| 基础指数Basal index (BI) | -0.53** | — | — | -0.56** | 0.73** | — |
| 通路指数Channel index (CI) | -0.66* | — | — | — | — | — |
| 自由生活成熟指数Maturity index (MI) | 0.70* | — | — | — | — | — |
| 植物寄生成熟指数 Plant-parasites index (PPI) |
0.72** | — | — | — | — | — |
| * P<0.05;** P<0.01 | ||||||
天然草地土壤线虫群落组成结构在种植燕麦后发生明显变化, 密度显著增加, 但多样性指数无显著变化。在该区域开展的其他研究也表明燕麦地的土壤动物密度及多样性高于天然草地和其他人工草地[11]。由表 1和图 3可知, 线虫群落组成结构和密度的变化主要是因燕麦地的食细菌和食真菌线虫密度显著增加造成, 其中原杆属、拟丽突属和滑刃属(Aphelenchoides)的密度变化最显著(表 1)。首先, 食物资源的供给状况是影响食细菌和食真菌线虫的主要因素之一[29-30]。已有研究表明, 植物生产力和凋落物质量的提高能促进土壤微生物的繁殖[31-32]。燕麦作为高寒地区应用最广泛的牧草, 其产量为天然高寒草地的10—15倍[5], 而燕麦植株体内的可溶性碳水化合物、粗蛋白含量也高于天然草地[6, 33]。因此, 其凋落物为土壤微生物提供了数量更多、质量更好的食物资源, 进而促进了食细菌和食真菌线虫。其次, 这三类线虫均属于c-p值较小的r-策略者(分别为Ba1、Ba2、Fu2), 其个体较小、生命周期短, 外源养分添加时能迅速繁殖[34], 因此在两种草地类型之间差异最大。此外, 耕作活动也是影响线虫群落结构的重要原因之一[35-36], 其他研究发现在耕种土壤中食细菌线虫取代植物寄生线虫成为土壤中的优势类群[34], 与本文的研究结果具有一致性。这可能是在耕种过程中, 对土壤的翻耕、施肥等措施能够改善土壤孔隙和通气状况, 促进微生物繁殖[37], 从而增加了食细菌和食真菌线虫的食物来源。食细菌和食真菌线虫数量的增加表明, 种植燕麦能够加强土壤线虫食物网的微生物能流渠道功能, 促进生态系统的物质循环和能量流动、提高草地生产力。
天然草地转变为燕麦地后, 虽然对植物寄生线虫密度无显著影响, 但能够降低植物寄生线虫的多样性, 改变其组成结构(表 1)。种植燕麦后, 植物寄生线虫中的丝尾垫刃属在群落中所占比例明显增加, 并出现巴兹尔垫刃属(Basiria)、米卡垫刃属(Miculenchus)等9个稀有类群;而新锥属(Neodolichodorus)、短体长针属(Longidorella)等16个属为天然草地所特有, 植物寄生线虫的多样性降低。植物寄生线虫组成的变化可能是受植物群落组成差异的影响[38]。在天然草地上种植燕麦后, 原有的植物群落地上和地下部分都发生较大改变, 尤其是植物种类显著减少, 引起植物寄生线虫生境的变化, 直接或间接的影响其类群组成和多样性[39]。
土壤理化性质对线虫群落也有重要影响[38]。回归分析结果显示, 食真菌和植物寄生线虫密度分别与土壤含水量呈显著正、负相关(表 2)。已有研究表明, 土壤水分增加可以显著提高真菌生物量[40], 从而促进食真菌线虫数量增加;而在一定干旱胁迫范围内, 燕麦植株体内有机物质向地下转移, 根系生物量增加[41-42], 可为植物寄生线虫提供更多的取食位点[28, 43], 间接促进植物寄生线虫。因此, 土壤水分变化对食真菌和植物寄生线虫的影响不同。本研究中, 燕麦地的土壤含水量显著高于天然草地(表 2), 因此燕麦地的食真菌线虫密度高于天然草地, 而植物寄生线虫低于天然草地。此外, 食细菌线虫与土壤pH呈显著正相关(表 3), 且燕麦地土壤pH也显著高于天然草地(表 2)。这表明建植燕麦人工草地也可通过影响土壤pH间接的提高食细菌线虫的数量。Korthals等研究也表明, 弱酸性土壤中食细菌线虫数量与土壤pH呈显著正相关, 且变化比其他线虫敏感[44]。而本研究中, 燕麦地在耕作过程中的翻耕、施肥等农措施, 可能促进了酸性土壤中的氧化还原反应和土壤呼吸, 降低土壤有机质含量, 使土壤pH升高[45], 进而提高了食细菌线虫密度。
本文及其他研究[11]表明天然草地转变为燕麦草地后可使土壤动物群落的密度及多样性增加, 因此建议在高寒草地上建植其他类型的人工草地时适当撒播燕麦种子, 建植混播草地, 以提高人工草地土壤动物群落多样性及其生态功能。
3.2 种植燕麦对土壤线虫群落生态指数影响种植燕麦后, MI指数显著增加但仍小于2, EI、BI指数也高于天然草地, 表明燕麦地的土壤养分富集状况和食物网抵抗力均高于天然草地。这可能是因为种植燕麦后, 线虫群落中Ba1、Ba2和Fu2功能团的线虫数量显著增加(附表 1), 从而影响了MI、EI和BI指数。燕麦地的PPI指数显著低于天然草地(图 4), 可能是由于植物群落多样性急剧下降, 对线虫取食承载能力降低[25]。
燕麦地SI指数低于天然草地但差异不显著, 这是因为SI指数主要受c-p值较大的捕食-杂食性线虫数量影响, 而种植燕麦后捕食-杂食性线虫密度变化不显著(图 3)。由于捕食-杂食性线虫是高营养级捕食者, 是决定土壤食物网复杂性、稳定性的关键生物类群[35], 因此由SI指数的变化可知种植燕麦后对土壤食物网结构稳定性影响较小。燕麦地CI指数与天然草地间无显著差异, 且均小于50, 说明两种草地的碎屑食物网均以细菌分解途径为主。而细菌的主要食物资源为低碳氮比的有机质, 易降解, 物质循环速率快, 有利于植物从土壤中吸收养分[46]。但从长期的观点看, 养分周转速率较快的燕麦地将会导致土壤中的有机质、氮、磷等养分消耗较快, 如果土壤有机质和养分等归还不足, 将会造成土壤肥力下降甚至衰竭[47]。这可能是高寒地区一些人工草地在建植4年左右后无法实现持续高产[48]的重要原因。因此, 建议在燕麦草地中适当增加凋落物及养分的还田量, 以保证燕麦草地土壤中的营养物质循环平衡。
3.3 种植燕麦对土壤线虫群落月份动态的影响线虫群落密度和多样性的季节变化动态受温度、湿度、食物供应、种内竞争及捕食作用等因素的综合影响[49]。本研究中, 燕麦地和天然草地土壤线虫的类群数和香农多样性指数均为9月显著大于7月, 而群落密度和优势度指数为7月大于9月。其原因可能是温度和湿度的月份变化对不同类群的线虫影响不同[50]。该区域7月的降水量和温度均高于9月[51], 使单宫属(Monhystera)等稀有类群消失, 拟丽突属、杆咽属(Rhabdolaimus)等少数线虫类群的密度和所占比例增加, 从而降低了7月份线虫群落的类群数和多样性指数, 这也与Papatheodorou等研究结果相似[52]。此外, 在7月份的燕麦地中拟丽突属和滑刃属显著增加并成为线虫群落的优势类群。因此, 7月燕麦地的优势度指数显著高于9月, 而天然草地差异不显著。土壤线虫群落密度及多样性指数的季节变化表明, 燕麦地土壤线虫群落组成结构在月份间的差异大于天然草地。
4 结论天然高寒草地转变为燕麦人工草地后, 短期内土壤线虫群落组成结构发生明显变化, 群落密度以及食细菌和食真菌线虫密度也显著增加, 使以植物寄生线虫为主的群落营养结构转变为以食细菌线虫为主, 增强了土壤线虫食物网的养分富集状况及抵抗力, 同时增加了土壤线虫群落的季节差异。由于天然草地转变为燕麦地后, 土壤动物多样性增加, 物质循环速率提高, 因此建议在高寒草地上建植人工草地时以燕麦混播草地为宜, 同时增加人工草地凋落物及养分的归还量, 以保证人工草地生态系统的物质循环平衡。
| [1] |
李其, 刘琳, 蔡义民, 裴姝婷, 罗英, 刘丽霞, 范慧, 孙飞达, 周春梅, 申旭东, 陈有军. 川西北高寒沙化草地治理恢复过程中CO2通量变化. 应用与环境生物学报, 2018, 24(3): 441-449. |
| [2] |
魏振海, 董治宝, 胡光印, 逯军峰. 近40a来若尔盖盆地沙丘时空变化. 中国沙漠, 2010, 30(1): 26-32. |
| [3] |
董世魁, 蒲小鹏, 胡自治. 青藏高原高寒人工草地生产-生态范式. 北京: 科学出版社, 2013.
|
| [4] |
侯众, 张新跃, 周俗, 唐川江, 张绪校, 严东海, 鲁岩. 2011年四川省草原监测报告//四川省畜牧兽医学会2012年学术年会论文集.成都: 四川省畜牧兽医学会, 2012: 553-557.
|
| [5] |
赵桂琴, 师尚礼. 青藏高原饲用燕麦研究与生产现状、存在问题与对策. 草业科学, 2004, 21(11): 17-21. |
| [6] |
彭先琴, 周青平, 刘文辉, 魏小星, 田莉华, 陈有军, 王沛. 川西北高寒地区6个燕麦品种生长特性的比较分析. 草业科学, 2018, 35(5): 1208-1217. |
| [7] |
官惠玲, 樊江文, 李愈哲. 不同人工草地对青藏高原温性草原群落生物量组成及物种多样性的影响. 草业学报, 2019, 28(9): 192-201. |
| [8] |
刘文辉, 魏小星, 刘芳, 秦燕, 张永超. 高寒区施肥和混播对燕麦人工草地植物器官碳氮储量分配的影响. 干旱地区农业研究, 2019, 37(4): 100-106. |
| [9] |
蒲琴, 胡玉福, 蒋双龙, 何剑锋, 舒向阳, 杨泽鹏. 不同生态治理措施下高寒沙化草地土壤氮素变化特征. 草业学报, 2016, 25(7): 24-33. |
| [10] |
伍文宪, 张蕾, 黄小琴, 杨潇湘, 薛龙海, 刘勇. 川西北高寒牧区不同人工草地对土壤微生物多样性影响. 草业学报, 2019, 28(3): 29-41. |
| [11] |
李雨, 吴鹏飞, 龙伟, 马金豪. 高寒地区种植不同种类牧草对土壤节肢动物群落的影响. 生态学报, 2019, 39(20): 7697-7708. |
| [12] |
Bardgett R D, Van Der Putten W H. Belowground biodiversity and ecosystem functioning. Nature, 2014, 515(7528): 505-511. |
| [13] |
Hunt H W, Coleman D C, Ingham E R, Ingham R E, Elliott E T, Moore J C, Rose S L, Reid C P P, Morley C R. The detrital food web in a shortgrass prairie. Biology and Fertility of Soils, 1987, 3(1/2): 57-68. |
| [14] |
Bardgett R D, Cook R, Yeates G W, Denton C S. The influence of nematodes on below-ground processes in grassland ecosystems. Plant and Soil, 1999, 212(1): 23-33. |
| [15] |
Bongers T. The maturity index:an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 1990, 83(1): 14-19. |
| [16] |
Ekschmitt K, Bakonyi G, Bongers M, Bongers T, Boström S, Dogan H, Harrison A, Nagy P, O'Donnell A G, Papatheodorou E M, Sohlenius B, Stamou G P, Wolters V. Nematode community structure as indicator of soil functioning in European grassland soils. European Journal of Soil Biology, 2001, 37(4): 263-268. |
| [17] |
肖冰雪, 陈琴, 郑群英, 道里刚, 李燕. 红原县饲草产业发展现状及对策探讨. 草学, 2018(6): 83-86. |
| [18] |
陈有军, 周青平, 孙建, 田莉华. 不同燕麦品种田间倒伏性状研究. 作物杂志, 2016(5): 44-49. |
| [19] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [20] |
Bongers T. Nematodes from Netherlands. Utrecht: Natuurhistorische Vereniging, 1994.
|
| [21] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
|
| [22] |
殷秀琴, 王海霞, 周道玮. 松嫩草原区不同农业生态系统土壤动物群落特征. 生态学报, 2003, 23(6): 1071-1078. |
| [23] |
Yeates G W, Bongers T, De Goede R G, Freckman D W, Georgieva S S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. Journal of Nematology, 1993, 25(3): 315-331. |
| [24] |
Yeates G W, Bongers T. Nematode diversity in agroecosystems. Agriculture, Ecosystems & Environment, 1999, 74(1/3): 113-135. |
| [25] |
Bongers T, van der Meulen H, Korthals G. Inverse relationship between the nematode maturity index and plant parasite index under enriched nutrient conditions. Applied Soil Ecology, 1997, 6(2): 195-199. |
| [26] |
Ferris H, Bongers T, de Goede R G M. A framework for soil food web diagnostics:extension of the nematode faunal analysis concept. Applied Soil Ecology, 2001, 18(1): 13-29. |
| [27] |
Berkelmans R, Ferris H, Tenuta M, van Bruggen A H C. Effects of long-term crop management on nematode trophic levels other than plant feeders disappear after 1 year of disruptive soil management. Applied Soil Ecology, 2003, 23(3): 223-235. |
| [28] |
Ferris H, Matute M M. Structural and functional succession in the nematode fauna of a soil food web. Applied Soil Ecology, 2003, 23(2): 93-110. |
| [29] |
Griffith B S, Ekelund F, Rønn R, Christensen S. Protozoa and nematodes on decomposing barley roots. Soil Biology and Biochemistry, 1993, 25(9): 1293-1295. |
| [30] |
李琪, 梁文举, 姜勇. 农田土壤线虫多样性研究现状及展望. 生物多样性, 2007, 15(2): 134-141. |
| [31] |
Balvanera P, Pfisterer A B, Buchmann N, He J S, Nakashizuka T, Raffaelli D, Schmid B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters, 2006, 9(10): 1146-1156. |
| [32] |
De Deyn G B, Quirk H, Bardgett R D. Plant species richness, identity and productivity differentially influence key groups of microbes in grassland soils of contrasting fertility. Biology Letters, 2010, 7(1): 75-78. |
| [33] |
吴发莉, 王之盛, 杨勤, 石红梅, 申俊华, 胡瑞, 邹华围. 甘南碌曲和合作地区冬夏季高寒天然牧草生产特性、营养成分和饲用价值分析. 草业学报, 2014, 23(4): 31-38. |
| [34] |
Bongers T, Bongers M. Functional diversity of nematodes. Applied Soil Ecology, 1998, 10(3): 239-251. |
| [35] |
Freckman D W, Ettema C H. Assessing nematode communities in agroecosystems of varying human intervention. Agriculture, Ecosystems & Environment, 1993, 45(3/4): 239-261. |
| [36] |
Lenz R, Eisenbeis G. Short-term effects of different tillage in a sustainable farming system on nematode community structure. Biology and Fertility of Soils, 2000, 31(3/4): 237-244. |
| [37] |
杨希智, 王长庭, 字洪标, 刘敏. 三江源区不同建植年限人工草地土壤微生物群落结构特征. 应用与环境生物学报, 2015, 21(2): 341-349. |
| [38] |
Wardle D A, Bardgett R D, Klironomos J N, Setälä H, Van Der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629-1633. |
| [39] |
Wardle D A, Yeates G W, Williamson W, Bonner K I. The response of a three trophic level soil food web to the identity and diversity of plant species and functional groups. Oikos, 2003, 102(1): 45-56. |
| [40] |
Williams M A, Rice C W. Seven years of enhanced water availability influences the physiological, structural, and functional attributes of a soil microbial community. Applied Soil Ecology, 2007, 35(3): 535-545. |
| [41] |
任永峰, 刘景辉, 李倩, 武俊英. 不同水分胁迫对裸燕麦根系生长的影响. 作物杂志, 2009(2): 17-21. |
| [42] |
张娜, 赵宝平, 任鹏, 刘景辉, 王莹. 水分胁迫对不同抗旱性燕麦品种根部生理特性的影响. 中国农学通报, 2012, 28(21): 63-69. |
| [43] |
Wang K H, McSorley R, Marshall A, Gallaher R N. Influence of organic Crotalaria juncea hay and ammonium nitrate fertilizers on soil nematode communities. Applied Soil Ecology, 2006, 31(3): 186-198. |
| [44] |
Korthals G W, Bongers T, Kammenga J E, Alexiev A D, Lexmond T M. Long-term effects of copper and pH on the nematode community in an agroecosystem. Environmental Toxicology and Chemistry, 1996, 15(6): 979-985. |
| [45] |
黄昌勇, 徐建明. 土壤学(第三版). 北京: 中国农业出版社, 2010.
|
| [46] |
Ingwersen J, Poll C, Streck T, Kandeler E. Micro-scale modelling of carbon turnover driven by microbial succession at a biogeochemical interface. Soil Biology and Biochemistry, 2008, 40(4): 864-878. |
| [47] |
Moretto A S, Distel R A. Competitive interactions between palatable and unpalatable grasses native to a temperate semi-arid grassland of Argentina. Plant Ecology, 1997, 130(2): 155-161. |
| [48] |
王长庭, 龙瑞军, 王启兰, 刘伟, 景增春, 张莉. 三江源区不同建植年代人工草地群落演替与土壤养分变化. 应用与环境生物学报, 2009, 15(6): 737-744. |
| [49] |
Freckman D W, Baldwin J G. Nematoda//Dindal D L, ed. Soil Biology Guide. New York: John Wiley & Sons, 1990: 155-200.
|
| [50] |
Bakonyi G, Nagy P, Kovács-Láng E, Kovács E, Barabás S, Répási V, Seres A. Soil nematode community structure as affected by temperature and moisture in a temperate semiarid shrubland. Applied Soil Ecology, 2007, 37(1/2): 31-40. |
| [51] |
李洪泉, 孙启忠, 李华德, 刘刚, 邱高林, 吴婍, 冯敬琪, 罗猛, 郭丽娟, 熊智斌, 杨龙, 杜寒晖. 易灾牧区红原县水热分布特点对抗灾饲草料生产收割和贮备的影响与对策. 草业与畜牧, 2015(4): 57-61. |
| [52] |
Papatheodorou E M, Argyropoulou M D, Stamou G P. The effects of large- and small-scale differences in soil temperature and moisture on bacterial functional diversity and the community of bacterivorous nematodes. Applied Soil Ecology, 2004, 25(1): 37-49. |