 2020, Vol. 40
2020, Vol. 40文章信息
- 杨威, 孙雨琛, 张婷婷, 刘琪, 黄悦, 葛茜, 邓道贵
- YANG Wei, SUN Yuchen, ZHANG Tingting, LIU Qi, HUANG Yue, GE Qian, DENG Daogui
- 富营养化对小型湖泊浮游甲壳动物群落结构及多样性的影响
- Impact of eutrophication on the community structure and species diversity of crustacean zooplankton in small lakes
- 生态学报. 2020, 40(14): 4874-4882
- Acta Ecologica Sinica. 2020, 40(14): 4874-4882
- http://dx.doi.org/10.5846/stxb201907251574
-
文章历史
- 收稿日期: 2019-07-25
- 修订日期: 2020-04-28






2. 淮北师范大学信息学院, 淮北 235000
2. Information College, Huaibei Normal University, Huaibei 235000, China
浮游甲壳动物(包括枝角类和桡足类), 处于水生食物链的中间环节, 主要以浮游植物为食, 同时也是鱼类的重要饵料, 在水体生态系统中具有重要的作用[1-3]。浮游甲壳动物个体小, 生命周期短, 对水环境变化敏感, 可在短时间内对环境变化做出反应。其群落多样性与水质状况密切相关, 可以作为水体营养状况的指示类群[4-5]。
湖泊富营养化已成为全球性的水环境污染问题[6]。我国湖泊数量多, 分布广, 以小型、浅水湖泊为主。目前, 我国约70%的湖泊受到了不同程度的污染, 多数湖泊出现了不同程度的富营养化[7]。研究表明, 湖泊富营养化对浮游动物的物种多样性和种群数量具有重要的影响[8]。随着水体富营养化的加剧, 浮游动物的密度和生物量也逐渐增加[9-10]。因此, 研究富营养化湖泊浮游动物群落结构和多样性的变化具有重要的意义。
近年来, 随着人类活动干扰和污染加剧, 临涣湖水体由轻度富营养化状态向富营养化状态转变[11-13]。因此, 本文通过对淮北临涣湖浮游甲壳动物群落结构和多样性的调查, 有助于探讨富营养化对湖泊浮游甲壳动物群落结构和多样性的影响, 以期为湖泊的生物多样性保护和富营养化治理提供参考。
1 材料与方法 1.1 采样地点和采样时间临涣湖(N33°37′, E116°36′)位于淮北市濉溪县西部, 属暖温带半湿润季风气候, 年均气温为15.2℃, 年均降雨量为842.7 mm[14]。临涣湖由采煤塌陷形成, 塌陷时间约17a, 水域面积约1.5 km2, 平均水深5.7 m。湖的东部为矸石山和粉煤灰厂, 西南部为临涣工业园和矸石堆积[14-15]。目前, 临涣湖用于渔业养殖, 主要养殖种类为鲢鱼和鳙鱼, 年均鱼产量约为2.7×104 kg/km2。
2017年3月至2018年2月对临涣湖浮游甲壳动物的群落结构进行调查, 根据湖泊的形态特征, 设置5个采样点(图 1), 每月采样一次。

|
| 图 1 临涣湖采样点位置 Fig. 1 The location of sampling sites in Lake Linhuan |
将13#浮游生物网(112 μm)在水面下约0.5 m处呈“∞”形缓慢拖行3—5 min, 将定性样品放入50 mL小塑料瓶中, 用4%的甲醛溶液固定。定量样品用5 L有机玻璃采水器在表层、中层和底层共采集30 L混合水样, 用25 #浮游生物网(64 μm)过滤后, 再用4%的甲醛溶液固定。
浮游甲壳动物定量样品用5 mL浮游生物计数框在显微镜下计数。种类鉴定参照《中国动物志·淡水枝角类》[16]和《中国动物志·淡水桡足类》[17], 生物量的计算参照章宗涉等的研究方法[18]。
1.3 理化指标的测定水温(water temperature, WT)和pH用pH计(Hach, 美国)现场测定, 透明度(secchi depth, SD)使用透明盘现场测定。总氮(total nitrogen, TN)用碱性过硫酸钾消解紫外分光光度法测定, 总磷(total phosphorus, TP)用过硫酸钾消解紫外分光光度法测定[19], 叶绿素a(chlorophyll a, Chl a)浓度的测定参照章宗涉等的研究方法[18]。
1.4 浮游甲壳动物的多样性和优势度采用Shannon多样性指数(H')、Pielou均匀度指数(J)、Simpson多样性指数(D)和优势度(Y)对浮游甲壳动物群落特征进行分析[20-22]:

|
(1) |
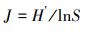
|
(2) |
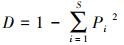
|
(3) |

|
(4) |
式中, S为种类总数;Pi为第i种浮游甲壳动物密度占总密度的比例;ni为第i种浮游甲壳动物的密度;fi为第i种浮游甲壳动物出现的频率;N为所有浮游甲壳动物的总密度。Y≥0.02的种类确定为优势种。
1.5 营养状态指数采用修正的卡尔森营养状态指数(TSIM)评价临涣湖营养化类型[23-24], 计算公式为:
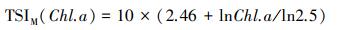
|
(5) |
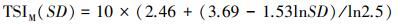
|
(6) |
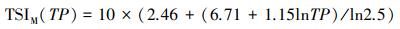
|
(7) |
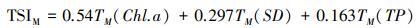
|
(8) |
式中, TSIM(Chl.a)、TSIM(SD)和TSIM(TP)是以叶绿素a浓度为基准的营养状态指数, Chl a、SD、TP的单位分别为μg/L、m、mg/L。
1.6 数据分析用SPSS 17.0软件分析浮游甲壳动物密度与理化因子的相关性。利用CANOCA 4.5软件分析浮游甲壳动物优势种与环境因子间的关系。先进行去趋势对应分析(DCA)以检验临涣湖的第1轴梯度长, 结果表明, 第1轴梯度长为2.334, 可见选择线性模型比选择单峰模型更为合理, 因此选择冗余分析(RDA)进行约束性排序。所有数据均进行ln(x+1)转换。
2 结果 2.1 理化因子临涣湖水温差异较大, 春季开始上升, 7月水温最高, 秋季水温开始下降, 1月最低, 年平均水温为17.8 ℃。总氮和总磷浓度的年平均值分别为1.19 mg/L和0.12 mg/L, 最大值均出现在10月, 最小值均出现在3月。透明度的年平均值为0.99 m, 最大值出现在1月, 最小值出现在5月。临涣湖pH偏碱性, 年平均值为8.45, 最大值出现在9月。叶绿素a浓度的变动范围较大, 年平均值为25.7 μg/L, 最大值出现在7月份(表 1)。
| 理化参数 Physico-chemical variables |
范围 Range |
平均值 Mean value |
相关性 Correlation |
显著性 Significance |
| 水温Water temperature/℃ | 2.5—31.0 | 17.8±9.7 | 0.679 | < 0.001 |
| 总氮浓度Total nitrogen concentration/(mg/L) | 0.59—1.98 | 1.19±0.41 | 0.055 | 0.677 |
| 总磷浓度Total phosphorus concentration/(mg/L) | 0.03—0.24 | 0.12±0.06 | 0.392 | 0.002 |
| 透明度Secchi depth/m | 0.73—1.03 | 0.99±0.25 | -0.458 | < 0.001 |
| 叶绿素a浓度Chlorophyll a concentration/(μg/L) | 8.8—43.4 | 25.7±11.6 | 0.614 | < 0.001 |
| pH | 8.23—8.69 | 8.45±0.12 | 0.382 | 0.003 |
临涣湖共记录浮游甲壳动物13种, 其中枝角类8属8种, 桡足类5属5种。春季种类数最多, 为10种, 秋冬季均为9种, 夏季种类数最少, 为5种, 且主要以小型浮游甲壳动物为主(表 2)。临涣湖在冬春季以象鼻溞(Bosmina sp.)、汤匙华哲水蚤(Sinocalanus dorrii)和近邻剑水蚤(Cyclops vicinus)为优势种。随着水温的上升, 夏秋季以短尾秀体溞(Diaphanosoma brachyurum)、广布中剑水蚤(Mesocyclops leuckarti)和象鼻溞(Bosmina sp.)为优势种。就全年的优势度而言, 象鼻溞(Bosmina sp.)为第一优势种, 其次是短尾秀体溞(D. brachyurum)和广布中剑水蚤(M. leuckarti)。
| 种类Species | 2017 | 2018 | |||||||||||
| 3月 March |
4月 April |
5月 May |
6月 June |
7月 July |
8月 August |
9月 September |
10月 October |
11月 November |
12月 December |
1月 January |
2月 February |
||
| 象鼻溞Bosmina sp. | 0.2 | < 0.1 | 10.1 | 9.1 | 6.3 | 4.7 | 40.6 | 30.7 | 11.8 | 4.4 | < 0.1 | 0.3 | |
| 方形尖额溞Alono quadranyularis | 0.1 | 0.1 | 0.1 | 0.1 | 0.3 | 0.1 | |||||||
| 盔形溞Daphnia galeata | 0.1 | < 0.1 | 0.1 | 0.1 | |||||||||
| 微型裸腹溞Moina micrura | 0.3 | 0.2 | 0.1 | 0.1 | |||||||||
| 角突网纹溞Ceriodaphnia cornuta | 0.1 | ||||||||||||
| 卵形盘肠溞Chydorus ovalis | 0.1 | 0.1 | 0.2 | < 0.1 | 0.1 | ||||||||
| 老年低额溞Simocephalus vetulus | 0.1 | ||||||||||||
| 短尾秀体溞Diaphanosoma brachyurum | 3.4 | 6.5 | 32.1 | 43.3 | 27.7 | 3.8 | 0.1 | ||||||
| 汤匙华哲水蚤Sinocalanus dorrii | 0.2 | 0.1 | 0.3 | 0.3 | 0.1 | 0.1 | 0.3 | ||||||
| 近邻剑水蚤Cyclops vicinus | 2.0 | 0.2 | 1.5 | 0.6 | 3.2 | ||||||||
| 广布中剑水蚤Mesocyclops leuckarti | 0.1 | 0.1 | 2.4 | 9.2 | 11.3 | 15.3 | 38.9 | 9.8 | 4.4 | 0.1 | 0.1 | ||
| 真剑水蚤Eucyclops sp. | 0.1 | 0.1 | |||||||||||
| 猛水蚤Harpacticoida | 0.1 | 0.1 | 2.0 | 0.1 | |||||||||
| 无节幼体Nauplius | 7.2 | 0.8 | 2.9 | 4.8 | 3.3 | 6.6 | 9.4 | 10.4 | 4.9 | 5.5 | 6.5 | 6.9 | |
象鼻溞(Bosmina sp.)、短尾秀体溞(D. brachyurum)和广布中剑水蚤(M. leuckarti)的年平均密度分别为9.8、9.7、7.6个/L, 分别占总密度的35%、34%和27%。象鼻溞(Bosmina sp.)的出现频率为100%, 全年都有分布, 最大密度(40.6个/L)出现在9月;短尾秀体溞(D. brachyurum)的出现频率为58%, 最大密度(43.3个/L)出现在8月;广布中剑水蚤(M. leuckarti)的出现频率为92%, 最大密度(38.9个/L)出现在9月(表 2)。
2.3 浮游甲壳动物密度和生物量的季节动态临涣湖浮游甲壳动物密度和生物量呈现出明显的季节变化, 且变化趋势相一致, 年平均密度和生物量分别为28.3个/L和0.33 mg/L, 最小密度和生物量(0.53个/L和0.01 mg/L)均出现在4月。随着水温的升高, 浮游甲壳动物开始大量繁殖, 到9月达到最大值(107.2个/L和1.07 mg/L), 10月开始逐渐下降, 12月—2月维持在较低水平(图 2)。

|
| 图 2 临涣湖浮游甲壳动物密度和生物量的季节变化 Fig. 2 Seasonal variations of density and biomass of crustacean zooplankton in Lake Linhuan |
临涣湖浮游甲壳动物的密度与水温(P<0.001)、总磷浓度(P<0.01)、透明度(P<0.001)、pH(P<0.01)和叶绿素a浓度(P<0.001)均呈极显著正相关, 与总氮浓度无相关性(表 1)。
2.4 浮游甲壳动物物种多样性指数临涣湖浮游甲壳动物的Shannon指数、Pielou指数和Simpson指数的季节差异均显著(P<0.05)。Shannon指数和Simpson指数的最大值均出现在6月, 最小值均出现在1月, 年平均值分别为0.86和0.49。Pielou指数的变化范围为0.58—0.96, 年平均值为0.74(表 3)。
| 多样性指数 Diversity index |
2017 | 2018 | |||||||||||
| 3月 March |
4月 April |
5月 May |
6月 June |
7月 July |
8月 August |
9月 September |
10月 October |
11月 November |
12月 December |
1月 January |
2月 February |
||
| Shannon指数Shannon index | 0.86 | 1.01 | 0.97 | 1.13 | 0.87 | 0.76 | 1.05 | 0.83 | 0.73 | 0.92 | 0.55 | 0.64 | |
| Pielou指数Pielou index | 0.67 | 0.90 | 0.74 | 0.87 | 0.79 | 0.63 | 0.96 | 0.69 | 0.66 | 0.68 | 0.75 | 0.58 | |
| Simpson指数Simpson index | 0.45 | 0.59 | 0.54 | 0.66 | 0.51 | 0.44 | 0.63 | 0.48 | 0.44 | 0.49 | 0.33 | 0.33 | |
临涣湖的TSIM值季节差异极显著(P<0.001), 在冬季较低, 秋季较高, 变化范围为54.2—67.1, 年平均值为62.6, 临涣湖水体处于富营养化状态。Pearson相关分析表明, 临涣湖的TSIM 值与浮游甲壳动物密度、主要优势种和Simpson指数均呈极显著正相关, 与Shannon指数和Pielou指数均呈显著正相关(表 4)。由此可见, 临涣湖水体富营养化初期明显提高了浮游甲壳动物的密度、均匀度和多样性, 对浮游甲壳动物群落具有重要的影响。
| 参数 Parameters |
Shannon指数 Shannon index |
Pielou指数 Pielou index |
Simpson指数 Simpson index |
象鼻溞 Bosmina sp. |
短尾秀体 D. brachyurum |
广布中剑水蚤 M. leuckarti |
浮游甲壳动物 Crustacean zooplankton |
| 相关性Correlation | 0.314 | 0.317 | 0.383 | 0.649 | 0.569 | 0.661 | 0.655 |
| 显著性Significance | 0.015 | 0.014 | 0.003 | < 0.001 | < 0.001 | < 0.001 | < 0.001 |
去趋势对应分析(DCA)显示, 第1轴梯度长为2.334, 可见选择线性模型比选择单峰模型更为合理。因此, 选择冗余分析(RDA)进行约束性排序。RDA分析结果显示, 第1轴和第2轴特征值分别为0.598和0.094, 前2轴共解释了95.0%的物种与环境的关系(表 5)。
| 排序轴Axes | 1 | 2 | 3 | 4 |
| 特征值Eigenvalues | 0.598 | 0.094 | 0.031 | 0.003 |
| 物种-环境相关系数Species-environment correlations | 0.889 | 0.866 | 0.736 | 0.484 |
| 累积变异百分比Cumulative percentage variance | ||||
| 物种Species data | 59.8 | 69.2 | 72.4 | 72.7 |
| 物种-环境Species-environment relation | 82.1 | 95.0 | 99.3 | 99.8 |
| 全部特征值总和Sum of all eigenvalues | 1.000 | |||
| 全部典范特征值总和Sum of all canonical eigenvalues | 0.729 |
由表 6可见, 两物种排序轴的相关系数为0.0006, 近似垂直, 两环境因子排序轴的相关系数为0, 表明排序结果可靠[25]。水温和叶绿素a浓度与第一排序轴呈极显著负相关, 相关系数分别为-0.7474和-0.6552;pH和透明度也与第一排序轴有相关性, 相关系数分别为-0.3676和0.4960;TN与第二排序轴正相关, 相关系数为0.3340;TP浓度与第一排序轴负相关, 与第二排序轴极显著正相关, 相关系数分别为-0.3424和0.6772;TSIM与第一排序轴极显著负相关, 相关系数为-0.6642。从第一、第二排序轴的相关性分析可知, TSIM、水温、叶绿素a浓度和TP浓度对浮游甲壳动物的密度变化有较大影响, 而透明度、TN浓度和pH的影响较小。
| 项目Item | SPEC AX1 | SPEC AX2 | ENVI AX1 | ENVI AX2 |
| TN | 0.0183 | 0.3340 | 0.0206 | 0.3856 |
| TP | -0.3424 | 0.6772 | -0.3852 | 0.7818 |
| Chla | -0.6552 | -0.0901 | -0.7371 | -0.1041 |
| T | -0.7474 | -0.1951 | -0.8408 | -0.2251 |
| pH | -0.3676 | -0.0122 | -0.4135 | -0.0141 |
| SD | 0.4960 | -0.0750 | 0.5580 | -0.0866 |
| TSIM | -0.6642 | 0.1002 | -0.7472 | 0.1157 |
| TN:总氮浓度Total nitrogen concentration;TP:总磷浓度Total phosphorus concentration;T:水温Water temperature;pH:酸碱度Potential of hydrogen;SD:透明度Secchi depth;Chla:叶绿素a浓度Chlorophyll a concentration;TSIM:营养指数Trophic state index | ||||
由图 3可知, 象鼻溞和广布中剑水蚤与叶绿素a浓度、TSIM、TP浓度和pH呈正相关关系, 与透明度负相关;短尾秀体溞与TSIM、水温、叶绿素a浓度和pH呈正相关关系;近邻剑水蚤和汤匙华哲水蚤与透明度呈正相关关系。

|
| 图 3 临涣湖浮游甲壳动物与环境因子的冗余分析 Fig. 3 Redundancy analysis between crustacean zooplankton and environmental factors in Lake Linhuan TN:总氮浓度Total nitrogen concentration;TP:总磷浓度Total phosphorus concentration;T:水温Water temperature;SD:透明度Secchi depth;Chla:叶绿素a浓度Chlorophyll a concentration;TSI:营养指数Trophic state index;Db:短尾秀体溞D. brachyurum;Ml:广布中剑水蚤M. leuckarti;Cv:近邻剑水蚤C. vicinus;Bo:象鼻溞Bosmina sp;Co:卵形盘肠溞C. ovalis;Aq:方形尖额溞A. quadranyularis;Ha:猛水蚤Harpacticoida;Sd:汤匙华哲水蚤Sinocalanus dorrii;Nu:无节幼体Nauplius |
水温、总氮和总磷是影响浮游甲壳动物群落结构的重要因素[26-28]。总氮和总磷能促进浮游植物的生长, 为浮游甲壳动物提供更多的食物来源, 从而间接的促进浮游甲壳动物的生长和繁殖[29-30]。在食物充足的条件下, 随着水温的升高, 浮游甲壳动物的发育时间缩短, 使浮游甲壳动物种群密度迅速增加, 进而影响浮游甲壳动物群落结构的变化[18, 31-32]。从第一、第二排序轴的相关性可知, 水温和TP对浮游甲壳动物的影响较大, 而TN的影响较小。夏秋季水温和总磷浓度相对较高, 浮游甲壳动物具有较高的密度, 而冬春季水温和总磷浓度相对较低时, 浮游甲壳动物密度也较低。Pearson分析表明, 临涣湖浮游甲壳动物密度与水温和总磷浓度均呈极显著正相关。RDA分析也表明, 水温和总磷浓度对浮游甲壳动物群落结构影响较大。因此, 水温和总磷浓度是影响临涣湖浮游甲壳动物群落结构的上行效应因子。这与于婷婷等的研究结果相一致[33]。
浮游甲壳动物主要以浮游植物为食, 浮游植物生物量的高低对浮游甲壳动物群落结构具有重要的影响[34]。叶绿素a浓度是衡量浮游植物生物量的重要指标, 在一定程度上可以体现出浮游甲壳动物的食物量[3, 35]。李娟等研究表明, 叶绿素a浓度与浮游甲壳动物呈正相关性, 对浮游甲壳动物群落结构的季节变化影响较大[8]。本研究中, 2017年7月—10月, 临涣湖叶绿素a浓度的含量较高, 为浮游甲壳动物提供更多的食物, 导致浮游甲壳动物的密度也较高。Pearson分析表明, 叶绿素a浓度与浮游甲壳动物密度呈显著正相关性。RDA分析也表明, 叶绿素a浓度与短尾秀体溞、象鼻溞和广布中剑水蚤呈正相关关系, 对浮游甲壳动物群落结构的影响较大。因此, 叶绿素a浓度是影响临涣湖浮游甲壳动物群落结构的重要因素。
鲢、鳙鱼对浮游动物的捕食具有明显的选择性, 主要以大型浮游甲壳动物为食, 导致浮游动物群落结构小型化[36]。但短尾秀体溞身体较透明, 且行动敏捷, 不易为鱼类察觉, 有较强的逃避能力[37]。杨威等(2019)研究表明, 鲢、鳙鱼的捕食压力和选择性捕食对浮游甲壳动物种类和密度具有重要的影响[38]。临涣湖主要养殖鲢、鳙鱼, 且以小型浮游甲壳动物(象鼻溞和广布中剑水蚤)及身体较透明的短尾秀体溞为优势种, 而盔形溞和近邻剑水蚤等大型浮游甲壳动物密度均较低。因此, 鲢、鳙鱼的选择性捕食是造成临涣湖浮游甲壳动物群落结构小型化的下行效应因子。
3.2 富营养化对浮游甲壳动物群落结构和物种多样性的影响水体富营养化与浮游甲壳动物群落结构的季节变化密切相关[9, 39]。鲁敏等研究表明, 营养水平是导致武汉东湖浮游甲壳动物群落结构在不同湖区具有不同分布格局的重要因素[40]。水体富营养化使一些敏感种类消失, 耐受种类大量繁殖[41-42], 导致浮游甲壳动物密度增加。临涣湖主要优势种为象鼻溞、短尾秀体溞和广布中剑水蚤, 均为富营养化指示种类。随着营养水平的升高, 其密度增大[43-45]。本研究中, Pearson分析结果表明, 营养状态指数(TSIM)与浮游甲壳动物密度呈极显著正相关, 水体富营养化提高了浮游甲壳动物的密度。RDA分析结果也表明, TSIM与短尾秀体溞、广布中剑水蚤和象鼻溞呈正相关关系, 对浮游甲壳动物的群落结构具有重要的影响。这与李娟等的研究结果一致[8]。
富营养化对浮游动物物种多样性的季节变化也具有重要的影响[46-48]。王颖等研究表明, 长期的水体富营养化降低了浮游甲壳动物的物种多样性[5]。本研究中, 营养状态指数(TSIM)与浮游甲壳动物的多样性指数呈正相关。随着水体富营养化的加剧, 浮游甲壳动物的多样性指数增大。湖泊富营养化的过程中, 多样性一般先升后降, 在中营养水平时, 群落结构复杂, 多样性高;在超富营养水平时, 群落结构简单, 多样性低[49]。氮、磷等营养盐水体显示, 临涣湖水体呈现中营养水平, 并向富营养化演变。因此, 随着营养状态指数(TSIM)的升高, 浮游甲壳动物的多样性呈上升趋势。
3.3 与淮南和淮北采煤塌陷湖泊的比较由表 7可知, 淮南和淮北采煤塌陷湖泊营养盐浓度、浮游甲壳动物群落结构及密度具有一定的空间差异。本次调查结果显示, 临涣湖浮游甲壳动物密度高于淮南和淮北塌陷湖泊, 且淮南和淮北塌陷湖泊浮游甲壳动物密度均较低。就种类而言, 淮南和淮北塌陷湖泊浮游甲壳动物种类少, 群落结构小型化。从营养盐来看, 淮北采煤塌陷湖泊总磷浓度均高于淮南塌陷湖泊, 而总氮浓度均低于淮南塌陷湖泊, 水体均呈现出不同程度的富营养化。因此, 要加强对淮南和淮北采煤塌陷湖泊水环境的保护, 减少鱼类养殖活动, 调整鱼类养殖结构, 可以降低鲢、鳙鱼对浮游甲壳动物的捕食压力, 促进生态系统的良性循环。
| 湖泊 Lakes |
总氮浓度 (TN)/(mg/L) |
总磷浓度 (TP)/(mg/L) |
物种数 Species number |
密度 Density/(个/L) |
叶绿素a (Chl a)/(μg/L) |
主要鱼类 Major fish species |
鱼产量 Fish yield/(kg/km2) |
优势种 Dominant species |
营养状态 Trophic state |
参考文献 References |
| 淮南潘集站 | 2.0 | 0.08 | 13 | 5.5 | 16.8 | 鲢鱼 鳙鱼 |
17×104 | 象鼻溞、短尾秀体溞 | 轻度富营养化 | 易齐涛等[50] |
| 淮南谢桥站 | 1.9 | 0.07 | 12 | 4.9 | 14.9 | 鲢鱼 鳙鱼 |
13×104 | 象鼻溞、多刺秀体溞 | 轻度富营养化 | 易齐涛等[50] |
| 淮南淮西湖 | 5.28 | 0.94 | 14 | 8.5 | 68.3 | 鲢鱼 鳙鱼 |
40×104 | 象鼻溞、近邻剑水蚤 | 重度富营养化 | 余艳霞[51] |
| 淮北东湖 | 1.52 | 0.14 | 8 | 12.8 | 55.8 | 鲢鱼 鳙鱼 |
2.5—3×104 | 广布中剑水蚤、短尾秀体溞、近邻剑水蚤 | 中度富营养化 | 杨威等[38, 52] |
| 淮北刘桥湖 | 1.35 | 0.138 | 10 | 4.4 | 59.0 | 鲢鱼 鳙鱼 |
2.5—3×104 | 广布中剑水蚤、短尾秀体溞、近邻剑水蚤 | 中度富营养化 | 杨威等[38, 52] |
| 淮北临涣湖 | 1.19 | 0.12 | 13 | 28.3 | 25.7 | 鲢鱼 鳙鱼 |
2.7×104 | 象鼻溞、广布中剑水蚤、短尾秀体溞 | 轻度富营养化 | 本研究 |
(1) 临涣湖共记录浮游甲壳动物13种, 其中枝角类8种, 桡足类5种。浮游甲壳动物密度和生物量的季节差异显著, 年平均密度和生物量分别为28.3个/L和0.33 mg/L。
(2) 临涣湖主要优势种为短尾秀体溞、广布中剑水蚤和象鼻溞。
(3) TN、TP和营养状态指数(TSIM)表明, 临涣湖水体处于富营养状态。TSIM值与浮游甲壳动物主要优势种、均匀度和多样性指数均呈显著正相关。
(4) 冗余分析结果表明, 水温、总磷浓度和叶绿素a浓度是影响临涣湖浮游甲壳动物群落结构变化的上行效应因子。
| [1] |
Feniova I Y, Razlutskij V I, Gladyshev M I, Kostrzewska-Szlakowska I, Majsak N N, Rzepecki M, Sushchik N N, Zilitinkevich N S. Factors of dynamics of plankton crustacean communities under eutrophic conditions. Russian Journal of Ecology, 2019, 50(1): 50-57. |
| [2] |
Piscia R, Mazzoni M, Bettinetti R, Caroni R, Cicala D, Manca M M. Stable isotope analysis and persistent organic pollutants in crustacean zooplankton:the role of size and seasonality. Water, 2019, 11(7): 1490. |
| [3] |
孙颖, 舒婷婷, 李静, 戴曦, 吕志均, 陈非洲. 南京玄武湖浮游甲壳动物的水平分布及其与环境的关系. 湖泊科学, 2012, 24(3): 422-428. |
| [4] |
林海, 王源, 李冰. 北京市妫水河浮游动物群落结构与水质评价. 生态学报, 2019, 39(20): 7583-7591. |
| [5] |
田雪, 付宏臣, 李宇, 姜明, 姜云垒. 2015年富锦国家湿地公园水体中浮游甲壳动物群落结构及多样性季节动态. 湿地科学, 2017, 15(1): 73-79. |
| [6] |
赵永宏, 邓祥征, 战金艳, 席北斗, 鲁奇. 我国湖泊富营养化防治与控制策略研究进展. 环境科学与技术, 2010, 33(3): 92-98. |
| [7] |
朱颖, 马骏, 陈睿哲. 富营养化的湖泊. 生态经济, 2018, 34(12): 6-9. |
| [8] |
李娟, 温周瑞, 李庚辰, 张敏. 太湖梅梁湾和贡湖湾浮游甲壳动物群落结构及其与环境因子的相互关系. 长江流域资源与环境, 2014, 23(S1): 81-90. |
| [9] |
García-Chicote J, Armengol X, Rojo C. Zooplankton species as indicators of trophic state in reservoirs from Mediterranean river basins. Inland Waters, 2019, 9(1): 113-123. |
| [10] |
王文侠, 陈非洲, 谷孝鸿. 南京市5座中型水库浮游动物群落结构及其与环境因子的关系. 湖泊科学, 2017, 29(1): 216-223. |
| [11] |
范廷玉, 程方奎, 严家平, 刘杰, 喻怀君, 张冰, 陈威. 淮北临涣煤矿塌陷水域叶绿素a与相关因子分析. 环境化学, 2015, 34(6): 1168-1176. |
| [12] |
任梦溪, 郑刘根, 程桦, 喻怀君. 淮北临涣采煤沉陷区水域水体污染源解析. 中国科学技术大学学报, 2016, 46(8): 680-688. |
| [13] |
汪晨琛.淮北临涣采煤沉陷湿地浮游植物群落结构研究[D].合肥: 安徽大学, 2018. http://cdmd.cnki.com.cn/Article/CDMD-10357-1018111979.htm
|
| [14] |
孔令健, 姜春露, 郑刘根, 程桦, 任梦溪, 闵飞虎, 方刘兵. 淮北临涣矿采煤沉陷区不同水体水化学特征及其影响因素. 湖泊科学, 2017, 29(5): 1158-1167. |
| [15] |
纪磊, 李晓明, 邓道贵. 淮北煤矿区塌陷湖大型底栖动物群落结构及水质生物学评价. 水生生物学报, 2016, 40(1): 147-156. |
| [16] |
蒋燮治, 堵南山. 中国动物志-淡水枝角类. 北京: 科学出版社, 1979.
|
| [17] |
沈嘉瑞. 中国动物志·淡水桡足类. 北京: 科学出版社, 1979.
|
| [18] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991.
|
| [19] |
国家环境保护总局《水和废水监测分析方法》编委会. 水和废水监测分析方法(第四版). 北京: 中国环境科学出版社, 2002.
|
| [20] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949.
|
| [21] |
Pielou E C. Ecological Diversity. New York: John Wiley, 1975.
|
| [22] |
徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13-15, 19-19. |
| [23] |
Aizaki M, Otsuki A, Fukushima T. Application of Modified Carlson's Trophic State Index to Japanese Lakes and Its Relationships to Other Parameters Related to Trophic State. Japan:Research Report of National Institute of Environmental Study, 1981, 13-31. |
| [24] |
杨梅玲, 胡忠军, 刘其根, 任丽萍, 陈来生, 李培培. 利用综合营养状态指数和修正的营养状态指数评价千岛湖水质变化(2007年-2011年). 上海海洋大学学报, 2013, 22(2): 240-245. |
| [25] |
Ter Braak C J F. Canonical correspondence analysis:a new eigenvector technique for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167-1179. |
| [26] |
刘宝贵, 刘霞, 吴瑶, 钟正, 陈宇炜. 鄱阳湖浮游甲壳动物群落结构特征. 生态学报, 2016, 36(24): 8205-8213. |
| [27] |
Yin J H, Zhang G T, Li C L, Wang S W, Zhao Z X, Wan A Y. Community composition, abundance and biomass of zooplankton in Zhangzi Island waters, Northern Yellow Sea. Chinese Journal of Oceanology and Limnology, 2017, 35(5): 1144-1151. |
| [28] |
赵会, 张坤, 彭水秀, 张萌, 邓道贵. 军山湖、青山湖和瑶湖春夏季节浮游甲壳动物的群落结构及其与环境因子的关系. 生态学杂志, 2018, 37(4): 1197-1203. |
| [29] |
杨亮杰, 吕光汉, 竺俊全, 徐镇, 金春华. 横山水库浮游动物群落结构特征及水质评价. 水生生物学报, 2014, 38(4): 720-728. |
| [30] |
陈星, 熊飞, 刘红艳, 张繁荣. 武汉三角湖浮游甲壳动物群落结构. 动物学杂志, 2017, 52(5): 824-834. |
| [31] |
Maier G. The effect of temperature on the development times of eggs, naupliar and copepodite stages of five species of cyclopoid copepods. Hydrobiologia, 1989, 184(1): 79-88. DOI:10.1007/BF00014304 |
| [32] |
Deng D G, Xie P. Effect of food and temperature on the growth and development of Moina irrasa (Cladocera:Moinidae). Journal of Freshwater Ecology, 2003, 18(4): 503-513. |
| [33] |
于婷婷, 朱雅, 徐珑, 赵磊, 钱文杰, 常青, 王国祥, 陈建琴. 苏州工业园区水体浮游甲壳动物群落特征及其与环境因子的关系. 应用生态学报, 2015, 26(8): 2534-2542. |
| [34] |
Yang W, Deng D G, Zhang S, Hu C L. Seasonal dynamics of crustacean zooplankton community structure in Erhai Lake, a plateau lake, with reference to phytoplankton and environmental factors. Chinese Journal of Oceanology and Limnology, 2014, 32(5): 1074-1082. |
| [35] |
王庆, 刘丽君, 杨宇峰. 南方某水库轮虫群落特征及其对环境因子的响应. 生态学报, 2010, 30(13): 3385-3395. |
| [36] |
杨宇峰, 黄祥飞. 鲢鳙对浮游动物群落结构的影响. 湖泊科学, 1992, 4(3): 78-86. |
| [37] |
李纯厚, 黄祥飞. 略论武汉东湖枝角类种类演替及其与生态因子的关系. 水生生物学报, 1992, 16(2): 101-112. |
| [38] |
杨威, 张婷婷, 孙雨琛, 刘琪, 葛茜, 邓道贵. 淮北采煤塌陷区两个小型湖泊浮游甲壳动物群落结构及其与环境因子的关系. 生态学杂志, 2019, 38(1): 188-196. |
| [39] |
邓道贵, 杨威, 孟小丽, 葛茜, 金显文, 胡振东. 淮河中游浮游甲壳动物群落结构的季节动态. 水生生物学报, 2013, 37(5): 869-875. |
| [40] |
鲁敏, 谢平. 武汉东湖不同湖区浮游甲壳动物群落结构的比较. 海洋与湖沼, 2002, 33(2): 174-181. |
| [41] |
钱方平, 席贻龙, 温新利, 黄林. 湖泊富营养化对轮虫群落结构及物种多样性的影响. 生物多样性, 2007, 15(4): 344-355. |
| [42] |
胡艺, 李秋华, 何应, 孟纯兰, 韩孟书. 贵州高原水库浮游动物分布特征及影响因子——以阿哈水库为例. 中国环境科学, 2020, 40(1): 227-236. |
| [43] |
徐杭英, 于海燕, 韩明春, 黄平沙, 李共国. 浙江饮用水源地浮游动物群落特征及环境响应. 生态学报, 2015, 35(21): 7219-7228. |
| [44] |
林青, 由文辉, 徐凤洁, 俞秋佳, 余华光. 滴水湖浮游动物群落结构及其与环境因子的关系. 生态学报, 2014, 34(23): 6918-6929. |
| [45] |
林志, 万阳, 徐梅, 汪晨琛, 吴奇丽, 周忠泽. 淮南迪沟采煤沉陷区湖泊后生浮游动物群落结构及其影响因子. 湖泊科学, 2018, 30(1): 171-182. |
| [46] |
郭凯, 赵文, 殷守仁, 李艳颍, 徐立蒲, 徐锋, 陈立斌. 北京官厅水库轮虫群落结构与水体富营养化状况. 湖泊科学, 2010, 22(2): 256-264. |
| [47] |
丁瑞睿, 郭匿春, 马友华. 巢湖流域双桥河底泥疏浚对浮游甲壳动物群落结构的影响. 湖泊科学, 2019, 31(3): 714-723. |
| [48] |
胡菊香, 吴生桂, 陈金生, 胡望斌, 唐会元, 邹清. 巢湖富营养化对轮虫的影响研究. 环境科学与技术, 2007, 30(12): 16-18, 68-68. |
| [49] |
日本生态学会环境问题专门委员会编.环境和指示生物﹒水域分册.卢全章, 译.北京: 中国环境科学出版社, 1987.
|
| [50] |
易齐涛, 汪琪, 谢凯, 张鑫. 淮南采煤塌陷湖泊浮游甲壳动物群落结构特征及其影响因素. 水生态学杂志, 2018, 39(3): 37-43. |
| [51] |
余艳霞.渔业捕捞方式对采煤塌陷湖泊生态系统营养结构和功能的影响[D].淮南: 安徽理工大学, 2018. https://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CMFD&filename=1018177286.nh
|
| [52] |
杨威, 张婷婷, 孙雨琛, 刘琪, 邓道贵, 金显文. 淮北采煤塌陷区两个小型湖泊轮虫的群落结构特征. 水资源保护, 2020, 36(1): 82-88, 96-96. |