 2020, Vol. 40
2020, Vol. 40文章信息
- 唐玥, 童春富, 刘毛亚, 朱宜平, 陈蓓蓓
- TANG Yue, TONG Chunfu, LIU Maoya, ZHU Yiping, CHEN Beibei
- 上海金泽水库典型挺水植物碳、氮、磷化学计量特征的季节变化
- Seasonal variations of carbon, nitrogen, phosphorus stoichiometry of four emergent hydrophytes in Jinze Reservoir, Shanghai
- 生态学报. 2020, 40(13): 4528-4537
- Acta Ecologica Sinica. 2020, 40(13): 4528-4537
- http://dx.doi.org/10.5846/stxb201912102682
-
文章历史
- 收稿日期: 2019-12-10
- 修订日期: 2020-05-08





2. 上海城投原水有限公司, 上海 200125
2. Shanghai Chengtou Raw Water Co., Ltd., Shanghai 200125, China
生态化学计量比为研究生态系统代谢和营养循环提供了一个强有力的方法[1-2]。碳(C)、氮(N)、磷(P)因其在生物化学功能中的强相互作用, 是生态化学计量学中研究最多的3种元素[3]。其中, 氮和磷是限制初级生产者生长的两种主要营养物质[4]。目前, 国内外针对水生生态系统中浮游植物、浮游动物和鱼类的化学计量特征已有广泛的研究[5-9], 而水生高等植物所受关注相对较少[1, 10]。
水生高等植物主要从沉积物和水体中摄取营养物质, 满足其对氮、磷等营养物质的需求[11]。然而, 外部氮、磷有效性与植物组织氮、磷含量之间的相关性有时并不强[12], 这意味着除外部氮、磷有效性外的其他因素也会导致植物氮、磷含量的变化。研究表明, 与生长速率、养分分配和贮藏有关的植物特性[13-15], 以及植株大小、植被类型和植物生活型[16-18]等均会引起植物氮、磷含量及其化学计量比的巨大变化。目前, 有关水生高等植物化学计量比的研究多集中在沉水植物[3, 19], 针对挺水植物的研究相对较少, 且大多数研究主要针对单一物种, 如芦苇或香蒲等[20-22], 缺乏多物种间的对比;涉及年周期、不同季节间对比分析的则更少[23-24]。甚至有学者认为水生植物的内稳态机制可能使主要元素的化学计量比维持某一动态平衡[1, 18, 25]。根据现有研究, 随着植物的生长, 其碳、氮、磷含量会随季节发生变化[23-24, 26], 那么其化学计量比是否会发生相应变化?不同物种其碳、氮、磷化学计量比的季节变化是否存在显著差异?针对这些问题, 本文选取了四种典型挺水植物作为研究对象, 通过固定样地的取样监测与分析, 探讨了碳、氮、磷化学计量比的变化特征, 进一步揭示了水生高等植物体内元素互相作用及变化规律, 为更好的研究水生生态系统的物质循环特征提供了重要基础。
1 材料与方法 1.1 研究区域与材料实验分析样品采自金泽水库。金泽水库位于上海市青浦区金泽镇, 太浦河北岸, 黄浦江上游, 平均海拔约3 m, 占地面积约2.7km2, 其中70%为水域, 为上海市西南五区提供水源。上海市青浦区2017年平均气温17.4℃, 年降水量1082.0 mm, 2018年平均气温17.5℃, 年降水量1280.6 mm[27-28]。最高平均气温出现在7、8月间, 而降水量2017年最大值出现在6月, 2018年出现在7月, 如图 2所示。金泽水库总库容910万m3, 近期供水量为351万m3/d, 远期将达500万m3/d, 实现黄浦江上游原水的集中供应。水库每日供水量较高, 水流速度快, 水库内不同区域水质较为均匀。作为新建水库, 不同区域滨岸带土壤质地基本一致。水库滨岸带植被主要为人工种植的挺水植物, 包括旱伞草(Cyperus alternifolius), 芦苇(Phragmites australis), 千屈菜(Lythrum salicaria), 水葱(Scirpus validus)等。其中, 芦苇主要分布在生态净化区, 其余3种植物主要分布在强制净化区, 如图 1所示。旱伞草为多年生常绿草本植物, 芦苇为多年生高大禾草, 千屈菜和水葱为多年生宿根草本植物, 四种植物均为地下芽植物, 耐寒性不强。芦苇的株高为1.5—3 m, 花期为8—12月;其他3种植物的花期为6—9月, 旱伞草的株高为0.6—1.5 m, 千屈菜为0.3—1.0 m, 水葱为1—2 m。

|
| 图 1 金泽水库采样站点示意图(JC4—JC9为生态净化区, JC1—JC3, JC10—JC12为强制净化区) Fig. 1 Sampling sites in Jinze Reservoir (JC4—JC9 are in the ecological purification area, JC1—JC3, JC10—JC12 are in the compulsory purification area) |

如图 1所示, 在金泽水库的生态净化区(JC4—JC9)和强制净化区(JC1—JC3, JC10—JC12)各设置6块样地, 分别于2017年10月、12月和2018年3月、6月(秋、冬、春、夏)4个不同季节采集植物地上部分样品。采样时均采集特定时期生长良好的代表性植株, 且不同样地采集的同种植物样本成熟度基本一致。其中, 芦苇在生态净化区采集, 其余3种植物在强制净化区采集。每个样地每种植物设置两个0.5 m×0.5 m的平行样方。采集样方内所有植物的地上部分, 并按物种分类后带回实验室做进一步处理。
1.3 样品处理植物样品在实验室于60℃烘干至恒重后称重, 获取生物量干重。烘干样品粉碎后, 过100目筛。使用元素分析仪(Vario Marco CNS)测定植物总碳、总氮含量。部分样品采用硫酸-过氧化氢消解后, 用钼锑抗分光光度法测定总磷含量[29]。
1.4 数据处理使用Origin 8.6作图, 采用统计软件SPSS 20.0进行数据的统计分析。采用单因素方差分析(One-way ANOVA)分别对不同植物C、N、P含量以及C/N, C/P和N/P不同季节间的差异进行显著性分析, 选用最小显著差法(LSD)进行均数间多重比较。用基于物种和季节两因素的双因素方差分析(Two-way ANOVA)的离差平方和(Sum of Squares of Deviations, SS)来表示它们对C、N、P含量以及化学计量比的方差贡献[12]。分析前对数据进行log10变换。方差分析的总离差平方和(SS)分解为:
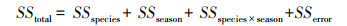
|
式中, SStotal, SSspecies, SSseason, SSspecies×season, SSerror分别表示总离差平方和、物种、季节、物种与季节交互作用以及误差项的离差平方和;将各因子的方差贡献表示为总离差平方和的百分比, 则SSspecies%+SSseason %+SSspecies×season%+SSerror%=1。各参数间的相关性分析采用Spearman相关分析[3]。
2 结果与分析 2.1 碳、氮、磷含量、化学计量比及生物量的变化特征植物地上生物量的变化特征如图 3所示, 不同植物存在明显差异。旱伞草的生物量季节变化不显著(P>0.05), 芦苇的生物量在夏季显著升高(P < 0.05), 其春季生物量显著低于夏秋两季(P < 0.05)。千屈菜的生物量在冬季显著降低(P < 0.05), 其秋季生物量显著高于冬季与春季(P < 0.05)。水葱冬季的生物量显著低于夏季(P < 0.05), 其他季节间并无显著差异(P>0.05)。

|
| 图 3 植物地上生物量的季节变化(平均值±标准误差) Fig. 3 Seasonal variations of above ground biomass of the plants (Mean ± SE) 图中标注不同的小写字母表示同一物种在不同季节间存在显著差异(P < 0.05) |
四种植物的碳、氮、磷含量及其化学计量比的变化特征如图 4所示。不同植物碳、氮、磷含量的季节变化特征并不相同;相较氮、磷含量而言, 植物碳含量的变化较小。其中, 旱伞草、芦苇的碳含量在4个季节间无显著差异(P>0.05), 千屈菜和水葱碳含量仅在冬季较其他季节显著偏低(P < 0.05)。旱伞草的氮含量在4个季节间均无显著差异(P>0.05), 千屈菜和芦苇冬季氮含量显著低于其他三个季节(P < 0.05)。水葱的氮含量在秋、冬、夏三个季节无显著差异(P>0.05)。旱伞草的磷含量仅在夏季显著低于其他三个季节(P < 0.05)。芦苇磷含量在冬季与夏季无显著差异(P>0.05), 但显著低于秋、春两季(P < 0.05)。水葱的磷含量在四季内均无显著差异(P>0.05), 而千屈菜秋季磷含量显著高于其他三个季节(P < 0.05)。

|
| 图 4 植物C、N、P含量及化学计量比的季节变化(平均值±标准误差) Fig. 4 Seasonal variations of C, N, P contents and stoichiometric ratios of the four plants (Mean ± SE) 图中标注不同的小写字母表示同一物种在不同季节间存在显著差异(P < 0.05) |
就植物化学计量比而言, 不同植物、以及同种植物的不同化学计量比随季节的变化规律并不一致。四种植物的N/P随季节变化特征较为一致, 均在冬季处于最高值。旱伞草的C/P在夏季显著高于其他三季(P < 0.05);芦苇的C/P在春季显著低于冬夏两季(P < 0.05);千屈菜C/P在秋季显著低于其他季节(P < 0.05), 而冬季C/P显著高于其他三季(P < 0.05);水葱C/P在4个季节间均无显著差异(P>0.05)。旱伞草的C/N和N/P在四季内均无显著性差异(P>0.05)。芦苇、千屈菜和水葱的C/N变化特征相似, 冬季处于最低水平, 在春季显著升高;但它们冬季的N/P均显著高于其他季节(P < 0.05)。
2.2 变动来源分析基于物种与季节变化的双因素方差分析的结果如表 1所示, 植物碳含量受季节变化的影响(SSseason%=32.29)远高于物种差异(SSspecies%=5.58)。植物氮含量受季节变化(SSseason%=23.35)和物种差异(SSspecies%=24.30)的影响程度相似。季节变化(SSseason%=34.78)对磷含量的影响大于物种差异作用(SSspecies%=17.55)。对于化学计量比而言, 物种差异对C/N和C/P的影响都略大于季节变化, 但对N/P来说, 季节变化(SSseason%=45.86)对其的影响程度远高于物种变化(SSspecies%=0.66)。
| SS物种% SSspecies% | SS季节% SSseason% | SS物种×季节% SSspecies×season% | SS误差% SSerror% | |
| C | 5.58 | 32.29** | 3.40 | 58.73 |
| N | 24.30** | 23.35** | 15.40* | 36.95 |
| P | 17.55** | 34.78** | 8.32 | 39.35 |
| C/N | 22.02** | 21.87** | 15.11 | 41.00 |
| C/P | 26.20** | 23.97** | 12.63* | 37.20 |
| N/P | 0.66 | 45.86** | 15.08* | 38.40 |
| *P < 0.05; **P < 0.01 SS:离差平方和Sum of Squares of Deviations | ||||
相关分析结果如表 2至表 5所示。不同季节植物碳氮磷含量的相关关系是不同的。秋季(2017年10月)时, 3种元素含量之间的相关关系均不显著(P>0.05);冬季(2017年12月)时, 除了C和N含量有显著负相关关系(P < 0.05)外, 其他元素含量间均无显著相关性(P>0.05);春季(2018年3月)时, C含量分别与N、P含量呈显著负相关(P < 0.01), 且N含量与生物量呈显著正相关(P < 0.01);夏季(2018年6月)时, 只有N与P含量呈显著正相关(P < 0.01), C含量及生物量与其他参数均无显著相关性(P>0.05)。
| C | N | P | C/N | C/P | N/P | 生物量Biomass | |
| C | 1 | ||||||
| N | -0.182 | 1 | |||||
| P | 0.143 | -0.082 | 1 | ||||
| C/N | 0.355* | -0.959** | 0.142 | 1 | |||
| C/P | 0.057 | -0.214 | -0.415** | 0.174 | 1 | ||
| N/P | -0.085 | 0.251 | -0.529** | -0.284 | 0.804** | 1 | |
| 生物量Biomass | 0.348 | 0.676* | 0.548* | -0.310 | 0.110 | -0.529* | 1 |
| *P < 0.05; **P < 0.01 | |||||||
| C | N | P | C/N | C/P | N/P | 生物量Biomass | |
| C | 1 | ||||||
| N | -0.742** | 1 | |||||
| P | -0.159 | 0.246 | 1 | ||||
| C/N | 0.827** | -0.955** | -0.238 | 1 | |||
| C/P | -0.028 | -0.287 | -0.509** | 0.234 | 1 | ||
| N/P | -0.741** | 0.652** | -0.164 | -0.675** | 0.407** | 1 | |
| 生物量Biomass | 0.643* | 0.029 | 0.587 | 0.024 | -0.595 | -0.677* | 1 |
| *P < 0.05; **P < 0.01 | |||||||
| C | N | P | C/N | C/P | N/P | 生物量Biomass | |
| C | 1 | ||||||
| N | -0.585** | 1 | |||||
| P | -0.517** | 0.371* | 1 | ||||
| C/N | 0.663** | -0.991** | -0.397** | 1 | |||
| C/P | 0.378* | -0.277 | -0.341* | 0.299* | 1 | ||
| N/P | -0.172 | 0.576** | -0.011 | -0.558** | 0.558** | 1 | |
| 生物量Biomass | 0.125 | 0.863** | 0.212 | -0.758** | -0.022 | 0.659* | 1 |
| *P < 0.05; **P < 0.01 | |||||||
| C | N | P | C/N | C/P | N/P | 生物量Biomass | |
| C | 1 | ||||||
| N | 0.001 | 1 | |||||
| P | 0.014 | 0.714** | 1 | ||||
| C/N | 0.163 | -0.969** | -0.694** | 1 | |||
| C/P | 0.179 | -0.657** | -0.655** | 0.683** | 1 | ||
| N/P | -0.042 | 0.113 | -0.201 | -0.105 | 0.571** | 1 | |
| 生物量Biomass | 0.236 | 0.465 | 0.125 | -0.658** | 0.235 | 0.597** | 1 |
| *P < 0.05; **P < 0.01 | |||||||
不同季节各元素含量与化学计量比之间的关系也不相同。除夏季外的三个季节中, C含量均与C/N呈显著正相关(P < 0.05)。所有季节中, N含量与C/N以及P含量与C/P均分别呈显著负相关(P < 0.05)。植物生物量在秋冬两季与N/P呈显著负相关(P < 0.05), 春夏两季与N/P呈显著正相关(P < 0.05)。所有季节N/P均与C/P呈极显著正相关(P < 0.01)。
3 讨论 3.1 植物C、N、P含量及其化学计量比变化特征已有研究表明, 不同季节植物体内碳、氮、磷含量及化学计量比会存在不同程度的动态变化[23-24], 植物氮、磷含量在不同生长阶段有较大的差异[30]。Li等人[20]的研究结果显示, 植物生长速率与叶片磷含量呈显著正相关, 在发育初期, 植物细胞生长迅速, 需要大量核酸装配进行快速分裂, 此时植物磷含量会有上升趋势。本次研究中植物地上生物量在6月普遍升高, 但植物磷元素含量并未显著升高, 可能由于此时挺水植物正处于快速生长时期时, 养分主要集中在根部, 而茎叶的快速增长可能稀释了体内的磷含量, 导致植物旺盛生长时期体内磷含量偏低[24, 31]。李川等人[32]的研究结果显示, 溱湖湿地的芦苇春季时氮含量低于冬季, 这与本文的研究结果一致;其夏季氮、磷含量均有显著升高, 而本研究中夏季芦苇氮含量有显著升高, 但磷含量有所下降。前人对芦苇、海三棱藨草等草本植物在杭州湾以及九段沙的研究结果均显示植物夏季磷含量降低[23, 26], 与本文结果相似。植物的氮、磷含量可以在一定程度上反映植物的生境条件[33], 如果在营养不够充足的环境, 植物的生长有可能会受到外界环境氮和磷有效性的抑制, 本次实验中植物夏季磷含量的偏低是否与土壤有效磷的供应有关, 还需结合土壤养分状况做进一步研究。
夏成星[34]在关于中国东部内陆水域水生植物的研究中发现, 挺水植物C/N和C/P均显著高于沉水植物和浮叶植物, 而不同生活型的水生植物N/P并没有显著差异。主要原因可能是挺水植物含有更多富含碳的结构性物质支撑其生长需要, 因而相比于沉水和浮叶植物具有更高的碳含量[34]。与陆生植物相比, 本文挺水植物平均N/P为29.9, 而我国东部地区陆生植物N/P为15.4[35], 西北地区主要草地植物N/P为15.3[36], 均低于本研究中挺水植物N/P。本文挺水植物C/N变化范围为9.58—92.45, 其最高值远低于阿拉善荒漠地区的灌木和草本植物(C/N变化范围9.52—396.99)[33]以及典型草原植物(C/N变化范围80.60—143.55)[37], 但高于亚热带常绿阔叶林典型植物的C/N(变化范围28.96—37.97)[38];本文挺水植物C/P变化范围为316.54—1830.92, 而阿拉善荒漠地区植物C/P变化范围为88.03—2685.35[33], 亚热带常绿阔叶林典型植物C/P变化范围为628.54—1035.7[38], 典型草原植物的C/P变化范围为342.37—1491.91[37]。说明不同地区、不同类型植物C/N、C/P存在较大差异, 而本研究中挺水植物的N/P高于大部分陆生植物。研究表明, 陆生植物主要从土壤中汲取营养, 植物地上部分的养分含量通常与土壤养分含量有一定相关性, 而土壤的成土特征易受到土壤母质、气候、地貌等因素影响, 导致不同环境下土壤养分元素差别较大, 进而影响植物的化学计量比[39]。水生植物相当一部分营养物质来自于水体, 水环境与土壤环境中养分的差异性同样会造成水生植物与陆生植物化学计量比的差异。而陆生植物相比水生植物含有更高的木质素、纤维素等富含碳的结构性物质, 这可能导致了其C/N与C/P相对水生植物更高[34]。本研究显示植物C/N, C/P, N/P存在季节变化, 这与王维琦等人[21]的研究结果一致。氮和磷在植物细胞生长中起着关键作用:氮主要存在于核酸、氨基酸和蛋白质中, 磷主要存在于核酸和三磷酸腺苷(酶的合成与细胞内能量传递系统的基础)中[1]。一般认为, 生态化学计量与RNA分配以及细胞生长速率有很大相关性, 植物高生长速率需要更多的rRNA以产生更多用于生长所需的蛋白质。由于rRNA是磷的主要赋存形式之一, rRNA含量的增加会导致细胞中磷浓度不成比例的增加, C/P和N/P会在植物快速生长时有所降低[13, 40-41]。植物体内N/P会在相对增长率较低时升高, 通过最大值, 然后在相对增长率较高时下降[13]。本研究中四种植物的C/P和N/P在冬季均出现了高值, 其中芦苇和千屈菜冬季的C/P显著高于秋季, 除旱伞草外, 其他3种植物冬季时N/P显著高于其他三个季节, 此时生物量与N/P呈显著负相关, 地上生物量也普遍下降, 其中千屈菜的地上生物量显著降低。主要原因可能是冬季气温偏低, 植物代谢过程减弱, 细胞分裂和分化能力降低, 这说明植物的低生长速率与高N/P和C/P值有一定联系, 与前人的研究结果相一致[13, 40-41]。而这两个指标在春季植物生长初期显著下降, 但此时植物地上生物量并未有明显升高, 与冬季无显著差异, 这与许多研究规律有所不同[13, 38, 41], 但植物的生长受多种因素影响, 化学计量比与生长季节间的联系还有待结合更多因子做一进步研究。
C/N和C/P可以在一定程度上反映植物对养分的吸收利用效率以及固碳效率[38], 本文结果显示春、夏季节植物C/N和C/P较高, 说明此时植物吸收营养元素和同化碳的能力均略高于其他季节。旱伞草的C/N、N/P在四季间均无显著差异, C/P只有在夏季时显著高于其他季节, 其生物量在一年四季均无显著差异, 这可能因为旱伞草是四季常绿植物, 其代谢强度变化不大的缘故。
3.2 季节变化对植物碳、氮、磷化学计量比特征的影响许多对植物化学计量比的研究表明, 氮、磷化学计量比受土壤肥力、温度、发育阶段和食草动物等多种因素的影响[25, 40, 42-44], 这就可能导致不同区域、季节及不同物种间植物氮、磷含量及化学计量比的差异。已有研究表明, 不同水生植物碳、氮、磷含量通常存在差异, 其中氮、磷的变异性更大, 碳含量的变异系数最小[45], 这与本文结果相似, 植物碳含量因物种差异而产生的变化远小于季节对其的影响。
旱伞草生物量在四季内无显著变化, 作为四季常绿植物, 其碳、氮含量全年无显著差异, 磷含量只在夏季时有所降低, 说明旱伞草的生长与代谢情况都较其他植物更为稳定, 体内营养元素的变化不显著。本文显示, 相比于碳含量而言, 物种差异对氮、磷含量的影响更为明显。植物花、叶的氮、磷含量一般会高于茎, 这些营养成分对植物生长很重要, 并且与器官功能有关, 茎和根茎的主要功能是水分和养分的输送和机械支持, 其氮、磷含量相对较低[46], 花和种子中富含线粒体, 其中含有大量氮和磷。植物的不同植物学特征可能导致物种间营养元素及化学计量比的差异, 本研究显示花期在6—8月的千屈菜在夏季氮、磷含量升高, 而芦苇的茎叶比通常在70%以上, 茎的氮、磷含量低可能导致芦苇整体氮、磷含量低于其他植物。本文显示季节变化对植物氮含量的影响程度与物种差异相当。除旱伞草外, 其他3种植物地上生物量在三月普遍偏低, 说明早春气温回升但植物并未处于快速生长阶段, 生长较为缓慢, 此时植物氮含量也偏低, 生物量与氮含量呈显著正相关。随着温度升高, 降水增多, 植被的生长发育开始加快[47], 物质积累迅速, 植物地上生物量较春季有所升高, 此时植物处于快速生长阶段, 氮含量也显著升高。
本研究显示, 相比物种差异而言, N/P主要受季节变化的影响, 这与Güeswell等人[12]的报道一致。前人的许多研究结果认为植物N/P与生长速率有一定联系[13, 40-41], 不同季节间N/P的较大差异可能反映了植物在不同季节生长速率的差异。常绿植物旱伞草N/P在四季间无显著差异, 其生物量也不随季节产生显著变化, 这表明其生长与代谢速率在全年都较稳定, 在适宜条件下不随气温或降水的变化而产生显著差异。其他3种植物N/P普遍在冬季升高, 此时气温处于全年最低值, 地上生物量普遍较低, 植物暂停生长, 这符合前人所说的高N/P与低生长速率相对应的规律[13, 40-41], 也说明冬季较低的气温影响了植物生长速率。本次研究中, 三月气温相较十二月已明显回升, 植物N/P在三月显著下降, 而此时地上生物量并不高, 说明早春虽然气温回升但植物并非处于快速生长阶段。随着气温持续上升, 植物生长速率加快, 但除芦苇外, 其他3种植物六月N/P并无显著变化, 这也表明植物化学计量比与气温的变化规律并非一致。本研究中, 植物在三月的低N/P值并未对应高的生长速率, 这与许多研究所得规律不一致[13, 40-41], 但也有研究显示N/P在植物生长初期处于最低值, 生长旺季先升高后降低, 叶片成熟不再生长时逐渐回升[23], 这与本次研究结果相符。
本研究中, C/N和C/P分别与N和P含量表现出显著的负相关关系, 其中C/N与N含量的负相关系数均大于0.95, 表明植物C/N和C/P分别受到氮和磷主导, 这与Demars等人[1]的研究结果相符。有很多相关研究的结果显示, 植物N与P含量具有较强的相关性, 但在本次研究中, 只有春季和夏季, 即植物生长季节显示出N和P的显著正相关, 而秋冬季节二者的相关性并不显著, 这也许与水库的特殊环境有关, 水库特殊的运行方式会带来一定的人为干扰因素, 同时, 有研究显示水环境中的氮、磷浓度与水生植物体内氮、磷含量有一定相关性[20], 而水源湖库作为集中式饮用水源地, 其水体氮、磷浓度相对偏低, 水体中的养分浓度对于植物生长发育、养分吸收以及体内生理调控会产生一定影响, 可能会使得植物体内氮磷比产生一定变化。
植物生态化学计量比的变化是多种非生物及生物因素共同作用的综合反映, 不同物种在不同季节下其体内碳、氮、磷及化学计量比的变化特征不尽相同。本研究就四种典型挺水植物不同生长季节碳、氮、磷及其化学计量比的变化特征及影响因素做了初步探究, 但植物的生长及养分的利用是一个复杂的过程, 受多种因素综合影响。在今后的研究中可结合更多环境因子以准确地分析植物的养分利用情况以及其与生境间C、N、P的交换过程。
| [1] |
Demars B O L, Edwards A C. Tissue nutrient concentrations in freshwater aquatic macrophytes:High inter-taxon differences and low phenotypic response to nutrient supply. Freshwater Biology, 2007, 52(11): 2073-2086. DOI:10.1111/j.1365-2427.2007.01817.x |
| [2] |
Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L W. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540-550. DOI:10.1046/j.1461-0248.2000.00185.x |
| [3] |
Xing W, Wu H P, Hao B B, Liu G H. Stoichiometric characteristics and responses of submerged macrophytes to eutrophication in lakes along the middle and lower reaches of the Yangtze River. Ecological Engineering, 2013, 54: 16-21. DOI:10.1016/j.ecoleng.2013.01.026 |
| [4] |
Elser J J, Bracken M E S, Cleland E E, Gruner D S, Harpole W S, Hillebrand H, Ngai J T, Seabloom E W, Shurin J B, Smith J E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 2007, 10(12): 1135-1142. DOI:10.1111/j.1461-0248.2007.01113.x |
| [5] |
Cáceres C E, Tessier A J, Andreou A, Duffy M A. Stoichiometric relationships in vernal pond plankton communities. Freshwater Biology, 2008, 53(7): 1291-1302. DOI:10.1111/j.1365-2427.2008.01962.x |
| [6] |
Elser J J, Andersen T, Baron J S, Bergström A K, Jansson M, Kyle M, Nydick K R, Steger L, Hessen D O. Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition. Science, 2009, 326(5954): 835-837. DOI:10.1126/science.1176199 |
| [7] |
Striebel M, Spörl G, Stibor H. Light-induced changes of plankton growth and stoichiometry:Experiments with natural phytoplankton communities. Limnology and Oceanography, 2008, 53(2): 513-522. DOI:10.4319/lo.2008.53.2.0513 |
| [8] |
Van de Waal D B, Verspagen J M H, Lürling M, Donk E V, Visser P M, Huisman J. The ecological stoichiometry of toxins produced by harmful cyanobacteria:An experimental test of the carbon-nutrient balance hypothesis. Ecology Letters, 2009, 12(12): 1326-1335. DOI:10.1111/j.1461-0248.2009.01383.x |
| [9] |
Vanni M J, Renwick W H, Bowling A M, Horgan M J, Christian A D. Nutrient stoichiometry of linked catchment-lake systems along a gradient of land use. Freshwater Biology, 2011, 56(5): 791-811. DOI:10.1111/j.1365-2427.2010.02436.x |
| [10] |
王芳, 国先涛, 董双林. 水域生态系统生态化学计量学研究进展. 中国海洋大学学报, 2015, 45(12): 16-23, 71-71. |
| [11] |
Cao T E, Ni L Y, Xie P, Xu J, Zhang M. Effects of moderate ammonium enrichment on three submersed macrophytes under contrasting light availability. Freshwater Biology, 2011, 56(8): 1620-1629. DOI:10.1111/j.1365-2427.2011.02601.x |
| [12] |
Güsewell S, Koerselman W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology, Evolution and Systematics, 2002, 5(1): 37-61. DOI:10.1078/1433-8319-0000022 |
| [13] |
Ågren G I. The C:N:P stoichiometry of autotrophs-theory and observations. Ecology Letters, 2004, 7(3): 185-191. |
| [14] |
Yu Q, Wu H H, He N P, Lü X T, Wang Z P, Elser J J, Wu J G, Han X G. Testing the growth rate hypothesis in vascular plants with above-and below-ground biomass. PLoS One, 2012, 7(3): e32162. DOI:10.1371/journal.pone.0032162 |
| [15] |
Ågren G I. Stoichiometry and nutrition of plant growth in natural communities. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 153-170. DOI:10.1146/annurev.ecolsys.39.110707.173515 |
| [16] |
He J S, Fang J Y, Wang Z H, Guo D L, Flynn D F B, Geng Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia, 2006, 149(1): 115-122. DOI:10.1007/s00442-006-0425-0 |
| [17] |
Zhang H Y, Wu H H, Yu Q, Wang Z W, Wei C Z, Long M, Kattge J, Smith M, Han X G. Sampling date, leaf age and root size:Implications for the study of plant C:N:P stoichiometry. PLoS One, 2013, 8(4): e60360. DOI:10.1371/journal.pone.0060360 |
| [18] |
Elser J J, Fagan W F, Kerkhoff A J, Swenson N G, Enquist B J. Biological stoichiometry of plant production:Metabolism, scaling and ecological response to global change. New Phytologist, 2010, 186(3): 593-608. DOI:10.1111/j.1469-8137.2010.03214.x |
| [19] |
Li W, Zhong J Y, Yuan G X, Fu H, Fan H B, Ni L Y, Xie P, Cao T. Stoichiometric characteristics of four submersed macrophytes in three plateau lakes with contrasting trophic statuses. Ecological Engineering, 2017, 99: 265-270. DOI:10.1016/j.ecoleng.2016.11.059 |
| [20] |
Li L P, Zerbe S, Han W X, Thevs N, Li W P, He P, Schmitt A O, Liu Y N, Ji C J. Nitrogen and phosphorus stoichiometry of common reed (Phragmites australis) and its relationship to nutrient availability in northern China. Aquatic Botany, 2014, 112: 84-90. DOI:10.1016/j.aquabot.2013.08.002 |
| [21] |
王维奇, 徐玲琳, 曾从盛, 仝川, 张林海. 河口湿地植物活体-枯落物-土壤的碳氮磷生态化学计量特征. 生态学报, 2011, 31(23): 7119-7124. |
| [22] |
屈凡柱, 孟灵, 付战勇, 孙景宽, 刘京涛, 宋爱云. 不同生境条件下滨海芦苇湿地C、N、P化学计量特征. 生态学报, 2018, 38(5): 1731-1738. |
| [23] |
吴统贵, 吴明, 刘丽, 萧江华. 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化. 植物生态学报, 2010, 34(1): 23-28. DOI:10.3773/j.issn.1005-264x.2010.01.005 |
| [24] |
牛得草, 李茜, 江世高, 常佩静, 傅华. 阿拉善荒漠区6种主要灌木植物叶片C:N:P化学计量比的季节变化. 植物生态学报, 2013, 37(4): 317-325. |
| [25] |
Yu Q, Elser J J, He N P, Wu H H, Chen Q S, Zhang G M, Han X G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia, 2011, 166(1): 1-10. DOI:10.1007/s00442-010-1902-z |
| [26] |
刘长娥, 杨永兴, 杨杨. 九段沙上沙湿地植物N、P、K的分布特征与季节动态. 生态学杂志, 2008, 27(11): 1876-1882. |
| [27] |
上海市统计局. 青浦区统计年鉴-2018. 北京: 中国统计出版社, 2018.
|
| [28] |
上海市统计局. 青浦区统计年鉴-2019. 北京: 中国统计出版社, 2019.
|
| [29] |
鲍士旦. 土壤农化分析 (第3版). 北京: 中国农业出版社, 2000.
|
| [30] |
Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385. DOI:10.1111/j.1469-8137.2005.01530.x |
| [31] |
Cronin G, Hay M E. Effects of light and nutrient availability on the growth, secondary chemistry, and resistance to herbivory of two brown seaweeds. Oikos, 1996, 77(1): 93-106. DOI:10.2307/3545589 |
| [32] |
李川, 朱陈名, 葛之葳, 李琪, 李昱, 薛建辉. 芦苇与土壤间氮磷化学计量的灰色关联分析. 南京林业大学学报:自然科学版, 2016, 40(2): 16-20. |
| [33] |
张珂, 何明珠, 李新荣, 谭会娟, 高艳红, 李刚, 韩国君, 吴杨杨. 阿拉善荒漠典型植物叶片碳、氮、磷化学计量特征. 生态学报, 2014, 34(22): 6538-6547. |
| [34] |
夏成星.中国东部内陆水域水生植物生态化学计量学研究[D].武汉: 武汉大学, 2014. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CDFD&filename=1015565974.nh
|
| [35] |
卢同平, 史正涛, 牛洁, 张文翔. 我国陆生生态化学计量学应用研究进展与展望. 土壤, 2016, 48(1): 29-35. |
| [36] |
He J S, Wang L, Flynn D F B, Wang X P, Ma W H, Fang J Y. Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biome. Oecologia, 2008, 155(2): 301-310. DOI:10.1007/s00442-007-0912-y |
| [37] |
陆颖, 王保林, 沈艳. 宁夏典型草原区不同退耕年限草地植物群落及优势植物C、N、P化学计量特征. 草业科学, 2019, 36(5): 1200-1206. |
| [38] |
柯立, 杨佳, 余鑫, 李鹏, 徐小牛. 北亚热带常绿阔叶林三优势树种叶水平碳氮磷化学计量及季节变化特征. 土壤通报, 2014, 45(5): 1170-1174. |
| [39] |
刘岑薇, 郑向丽, 王俊宏, 徐国忠, 黄毅斌. 陆生和水域生态系统植物的C、N、P生态化学计量特征研究综述. 中国农学通报, 2017, 33(17): 70-75. DOI:10.11924/j.issn.1000-6850.casb16090094 |
| [40] |
Matzek V, Vitousek P M. N:P stoichiometry and protein:RNA ratios in vascular plants:an evaluation of the growth-rate hypothesis. Ecology Letters, 2009, 12(8): 765-771. DOI:10.1111/j.1461-0248.2009.01310.x |
| [41] |
Vrede T, Dobberfuhl D R, Kooijman S A L M, Elser J J. Fundamental connections among organism C:N:P stoichiometry, macromolecular composition, and growth. Ecology, 2004, 85(5): 1217-1229. |
| [42] |
Güsewell S. N:P ratios in terrestrial plants:Variation and functional significance. New Phytologist, 2004, 164(2): 243-266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [43] |
Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006. DOI:10.1073/pnas.0403588101 |
| [44] |
Zhang G M, Han X G, Elser J J. Rapid top-down regulation of plant C:N:P stoichiometry by grasshoppers in an Inner Mongolia grassland ecosystem. Oecologia, 2011, 166(1): 253-264. DOI:10.1007/s00442-011-1904-5 |
| [45] |
Duarte C M. Nutrient concentration of aquatic plants:Patterns across species. Limnology and Oceanography, 1992, 37(4): 882-889. DOI:10.4319/lo.1992.37.4.0882 |
| [46] |
Baldantoni D, Alfani A, Di Tommasi P, Bartoli G, De Santo A V. Assessment of macro and microelement accumulation capability of two aquatic plants. Environmental Pollution, 2004, 130(2): 149-156. |
| [47] |
苏嫄, 焦菊英, 马祥华. 黄土丘陵沟壑区主要群落地上生物量季节变化及其与土壤水分的关系. 水土保持研究, 2012, 19(6): 7-12. |