 2020, Vol. 40
2020, Vol. 40文章信息
- 王楠, 潘小承, 白尚斌
- WANG Nan, PAN Xiaocheng, BAI Shangbin
- 模拟酸雨对我国亚热带毛竹林土壤呼吸及微生物多样性的影响
- Effects of simulated acid rain on soil respiration and microbial diversity in Moso bamboo forest in subtropical China
- 生态学报. 2020, 40(10): 3420-3430
- Acta Ecologica Sinica. 2020, 40(10): 3420-3430
- http://dx.doi.org/10.5846/stxb201905241073
-
文章历史
- 收稿日期: 2019-05-24
- 修订日期: 2020-03-18






2. 浙江农林大学浙江省森林生态系统碳循环与固碳减排重点实验室, 杭州 311300
2. Zhejiang Provincial Key Laboratory of Carbon Cycling in Forest Ecosystems and Carbon Sequestration, Zhejiang Agricultural and Forestry University, Hangzhou 311300, China
酸雨作为全球性环境问题之一, 已受到国际社会普遍关注[1]。我国酸雨区面积仍不断扩大, 危害程度逐渐加重, 已超过国土面积的40%, 成为继欧美之后的第三大酸雨区域, 其中亚热带地区酸雨污染尤为严重[2]。浙江位于我国中亚热带, 全省平均酸雨率超过80%, 是酸雨发生典型区域[3]。酸雨给森林生态系统带来严重影响和破坏, 不仅对植物叶片造成直接损害, 使叶片变黄甚至脱落, 影响植物光合作用, 而且可导致土壤酸化, 造成钾、钙、镁等盐基离子淋溶, 从而使土壤贫瘠, 影响植物正常生长发育[4]。同时, 土壤酸化使Al3+增加, 引起铝中毒, 毒害植物根系[5]。土壤酸化还能促进H+浓度增加, 引起土壤微生物群落组成和结构发生变化, 不可避免地影响着土壤碳循环过程[6-7]。
土壤呼吸作为陆地生态系统碳循环的一个主要组分, 在调控大气CO2浓度和气候变化方面起着关键作用[8]。土壤碳库是大气碳库的4倍, 土壤呼吸释放碳的速率比人类活动释放碳的速率要大一个数量级, 其微小变化能极大改变大气CO2浓度[9]。森林是陆地生态系统的主体, 其碳库占全球总碳库的46%, 占土壤呼吸总量的69%, 因此, 森林土壤呼吸成为陆地生态系统土壤呼吸的重要组分, 其动态变化也对全球碳平衡影响深远[10]。土壤呼吸主要包括植物根系呼吸和土壤微生物分解两个过程, 呼吸速率受植被特征、气候、水热条件、环境变化等诸多因素影响。酸雨可通过影响植物根系生长及影响微生物组成和活性变化来改变森林土壤呼吸速率[7]。一些研究认为酸雨促进了土壤呼吸[11], 也有研究认为酸雨会抑制或不影响土壤呼吸速率[12]。目前关于酸雨影响森林土壤呼吸还未有明确定论, 有待进一步深入研究。
竹林是世界重要的森林类型之一, 蕴含着巨大的固碳增汇潜力。我国竹子有39属, 500余种, 面积约601万公顷, 是竹子资源最丰富的国家, 其中毛竹林(Phyllostachys edulis)面积约占73.8%。毛竹具有较高的社会和经济效益, 是应对全球变化的重要树种, 已得到广泛推广种植[13-14]。影响毛竹林土壤呼吸的环境因子诸多, 其中酸雨胁迫是一个主要因素, 然而关于酸雨对毛竹林土壤呼吸的影响及其微生物学响应机制研究却相对较少[15]。因此, 本文以我国热带区域分布较多的毛竹林为研究对象, 采用PCR-DGGE技术, 探讨酸雨对毛竹林土壤细菌和真菌群落结构及多样性的影响, 及其与土壤呼吸的关联, 揭示土壤碳释放模式的变化, 从而为进一步研究亚热带毛竹林土壤碳释放对环境问题的响应提供理论基础。
1 研究地区与研究方法 1.1 研究地区概况研究区域位于浙江省杭州临安天目山国家级自然保护区(30°18′30″—30°21′37″N, 119°24′11″—119°27′11″ E), 保护区海拔300—1500 m, 属北亚热带季风气候, 四季分明, 温暖湿润, 年均气温14.5℃, 年年平均降雨量1400 mm。其森林植被大体分为6种类型:常绿阔叶林, 常绿落叶阔叶混交林, 落叶阔叶林、针阔混交林, 针叶林、毛竹林。毛竹林主要分布在海拔350—950 m。林冠层树种主要有青冈(Cyclobalanopsis glance)、木荷(Schima superba)、苦槠(Castanopsis sclerophylla)、毛竹(Phyllostachys pubscens)等;灌木层有毛花连蕊茶(Camellia fraternal)、山矾(Symplocos caudate)和山胡椒(Linder glauce)等;草本层有菊科(Compositae)和禾本科(Gramineae)等植物[16]。
1.2 试验样地设计实验样地位于海拔485 m的毛竹林内, 共设置3个样地, 在每个实验样地分别设置2个模拟酸雨小区和1个对照小区, 在处理1(T1)小区喷洒pH 4.0的模拟酸雨;在处理2(T2)小区中喷洒pH 2.0的模拟酸雨;在对照(CK)小区喷洒pH值约为5.8的当地湖水。3个样地呈三角形分布, 间隔距离100 m, 同一样地的各处理在同一水平地上, 缓冲间隔5 m, 每个小区面积为1 m×1 m, 3个样地共计9个小区。参照临安市以往的常规酸雨监测资料[6], 按照H2SO4 :HNO3=8:1的摩尔比配置模拟酸雨母液, 再用母液与去离子水配成相应pH值的模拟酸雨。自2016年9月开始模拟酸雨处理, 喷淋频率为每两周一次(如遇雨天或土壤湿度过大情况则顺延, 另择时间进行喷淋, 以防止喷洒的酸雨被地表径流输出)。每个小区每次喷洒酸雨量为1 mm, 对照小区等量喷洒pH 5.8的湖水。
1.3 土壤呼吸速率及温度测定从2016年9月开始, 进行了为期2年的土壤呼吸观测试验。预先把直径20 cm、高10 cm的土壤呼吸底座埋入每个小区, 将PVC环的一端沿坡面压入土中约5 cm深, 尽量减少布置PVC环对土壤的镇压作用, 基座埋设过程中尽量不破坏土壤, 以减少土壤扰动及根系损伤对测量结果的影响, PVC环在整个测量期间位置不变[17]。试验期间, 定期去除掉底座内的植物, 以避免测定的土壤呼吸包含植株呼吸作用。
于每个观测日8:00—12:00采用Li-8100土壤碳通量观测系统(Li-COR, Lincoln, Nebraska, USA)测定土壤呼吸, 测定频率为每月2次, 每次每个PVC环重复测定3次, 取其平均值代表该次测定值, 土壤呼吸测定在每次模拟酸雨喷洒前进行。在测定土壤呼吸的同时, 采用Li-8100土壤碳通量观测系统自带的温度探头测5 cm深处的土壤温度。
1.4 土壤生化指标的测定于2018年8月(模拟酸雨二年后)进行土壤采集。分别在每个小区的四角与中心布设5个取样点, 采用无菌土钻取表层大约5—20 cm深度的土壤样品, 更换样地取样前对工具采取清洗杀菌措施。每个取样点中大约取出0.5 kg的土壤样品, 然后将每个土样分为2份, 一份土样装入无菌自封样品袋中并置于4℃电子恒温箱中, 9个小区中共获取45份土样, 用于测定土壤化学性质。将同一小区剩余5份土样充分混合, 过2 mm筛网, 去除杂草与石块后装入无菌自封样品袋中, 并储存与-80℃的干冰保温箱中, 用于提取DNA, 共获取9个土壤微生物样品。
土壤pH值采用1:2.5土水比, 电位法测定;土壤溶解性有机碳(Soil dissolved organic carbon, DOC)使用高纯度水浸提, 取上清液过0.45 μm滤膜后使用岛津TOC-VCPH分析仪测定;碱解氮(Available nitrogen, AN)采用碱解蒸馏法测定;有效磷(Available phosphorous, AP)采用钼锑抗分光光度法测定;速效钾(Available potassium, AK)采用火焰原子吸收法测定;土壤微生物生物量碳(Soil microbial biomass of carbon, MBC)采用氯仿熏蒸-K2SO4浸提法测定[18]。
1.5 土壤细菌和真菌的变性梯度电泳(DGGE)采用DGGE方法对土壤细菌和真菌进行检测, 细菌采用16S rDNA-PCR扩增所用一对通用引物338F-GC和518R, 扩增片段长约250 bp;真菌18S rDNA-PCR扩增所用一对通用引物FR1-GC和FF390, 扩增片段长约390 bp。20 μL反应体系如下:10×Premix Tap 10 L, 引物(10 μmol/L)各0.2 μL, 模板DNA 0.3 μL, 牛血清蛋白(BSA, 20 mg/mL)0.2 μL。细菌的PCR反应参数:94℃预变性5 min, 94℃变性1 min, 50℃退火1 min, 72℃延伸1 min, 30个循环, 最后72℃延伸10 min。真菌的PCR反应参数:95℃预变性8 min, 94℃变性30 s, 50℃退火45 s, 72℃延伸2 min, 30个循环, 最后72℃延伸10 min。取3 μL PCR产物经1.0%琼脂糖凝胶电泳检测产物及其浓度[19]。
采用The DCodeTM Universal Mutation Detection System(Bio-Rad), 细菌扩增产物在变性梯度为45%—60%的6%聚丙烯酰胺凝胶上进行电泳。真菌扩增产物在变性梯度为25%—40%的6%聚丙烯酰胺凝胶上进行电泳。细菌和真菌的电泳过程条件一致:在60℃、80 V条件下电泳13 h, 使用SYBR greenⅠ染色0.5 h, 染色结果于Gel Doc TM EQ(Bio-Rad)凝胶成像系统成像, 使用Quantity One 4.4软件(Bio-Rad)进行图像分析。
1.6 数据分析土壤呼吸速率与土壤5 cm深温度的关系采用如下指数模型:
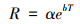
|
式中, R为土壤呼吸速率;T为土壤5 cm深温度;α是温度为0℃时土壤呼吸速率;b为温度反应系数。
Q10值的计算公式[20]:
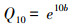
|
Shannon多样性指数(H)、Margalef丰富度指数(D)以及Pielou均匀度指数(E)对土壤细菌微生物群落多样性进行分析[21], 其计算公式为:

|
式中, ni为每条电泳条带光密度峰值, N为同一泳道中所有条带光密度峰值总和。电泳条带光密度的峰值通过分析软件Quantity one进行读取。
采用Origin Pro8.0软件对数据进行处理和绘图, 采用Pearson法进行相关性分析, 采用t检验进行不同处理土壤呼吸速率的均值比较, 采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)对3种处理间土壤呼吸速率进行方差分析和多重比较, 用Curve Estimation对土壤呼吸速率与土壤温度关系进行回归分析, R语言vegan包进行冗余分析(Redundancy Analysis, RDA)和作图。
2 结果与分析 2.1 不同酸雨处理土壤呼吸的季节变化土壤呼吸速率具有明显的季节动态规律, 最高值出现在7—8月, 最低值出现在12月份至次年2月期间。如图 1所示, 观测期可分为5个阶段:阶段1(2016年9月—2017年2月)模拟酸雨对土壤呼吸速率的影响没有显著差异;阶段2(2017年3月—2017年6月)模拟酸雨对土壤呼吸速率产生了促进作用, 模拟强酸降雨样地与对照之间具有显著差异(P < 0.05);阶段3 (2017年7月—2017年10月)模拟强酸降雨与对照样地、模拟弱酸降雨样地之间具有显著差异(P < 0.05), 而对照样地与模拟弱酸降雨样地之间的差异不显著, 阶段4(2017年11月—2018年2月)模拟酸雨对土壤呼吸速率的影响没有显著差异;阶段5(2018年3月—2018年8月)模拟酸雨对土壤呼吸速率产生了抑制作用, 且模拟强酸降雨样地与对照之间具有及显著差异(P < 0.05)。

|
| 图 1 不同酸雨处理下毛竹林土壤呼吸动态变化 Fig. 1 Dynamic change of soil respiration in moso bamboo under different acid rain treatments during 2016—2018 CK:对照处理, control treatment;T1:处理一, treatment 1;T2:处理二, treatment 2 |
根据对土壤温度检测数据分析(图 2), 土壤呼吸速率与土壤温度的季节变化趋势相近, 呈显著正相关关系, 方差分析表明不同模拟酸雨处理的土壤温度无显著差异, 年均土壤温度为16.6℃。

|
| 图 2 土壤呼吸与土壤温度的关系 Fig. 2 Relationship between soil respiration and soil temperature |
回归分析表明, CK、T1、T2处理的土壤呼吸与土壤温度之间呈极显著的指数回归关系(表 1), 这3个小区的回归方程的决定系数R2分别为0.960、0.937、0.845, 这表明土壤温度可分别解释这3个小区土壤呼吸96.0%、93.7%、84.5%的季节变异。根据指数回归方程估算得到的CK、T1、T2处理的土壤呼吸的温度敏感系数(Q10)分别为2.14、2.10、2.86。T2与T1和CK间差异显著, T1与CK间无显著性差异, 这一结果表明, 重度酸雨处理增加了温度敏感性。
| 处理 Treatment |
拟合方程 Fitted equation |
R2 | Q10 |
| CK | y = 1.129e0.076T | 0.960** | 2.14b |
| T1 | y = 1.129e0.074T | 0.937** | 2.10b |
| T2 | y = 1.193e0.105T | 0.845** | 2.86a |
| 不同字母表示同一列内各均值存在显著差异(P < 0.05);*表示显著相关(P < 0.05), **表示极显著相关(P < 0.01); CK:对照处理, control treatment;T1:处理一, treatment 1;T2:处理二, treatment 2 | |||
模拟酸雨对土壤pH值、养分及碳、氮含量的影响差异显著(表 2)。T2处理土壤的平均pH值比CK处理低17.31%, 比T1处理低14.00%;T2处理土壤有机质含量比CK低47.55%, 比T1处理低30.67%;CK处理土壤DOC、AK、MBC的含量分别是T1处理的1.22、1.16、1.01倍, 分别是T2处理的1.39、1.36、1. 06。T2处理土壤AN、AP的含量分别是CK处理的1.09、1.67倍, T1的1.05、1.62倍。由此可见, 不同强度的酸雨对土壤理化性质和微生物量具有显著的影响。
| 处理 Treatment |
pH | 碱解氮 Available N/(mg/kg) |
有效磷 Available P/(mg/kg) |
速效钾 Available K/(mg/kg) |
可溶性有机碳 Dissolved organic carbon/(mg/kg) |
微生物量碳 Microbial biomass carbon/(mg/kg) |
| CK | 5.2±0.04a | 221.82±3.14c | 3.50±0.07b | 44.68±3.40a | 138.38±2.04a | 607.71±0.38a |
| T1 | 5.0±0.08b | 230.45±2.68b | 3.59±0.09b | 38.52±0.59b | 113.57±3.96b | 598.78±2.42b |
| T2 | 4.3±0.03c | 242.83±4.49a | 5.83±0.10a | 32.78±2.20b | 99.55±1.27c | 573.08±2.02c |
| 不同字母表示同一列内各均值存在显著差异(P < 0.05); CK:对照处理, control treatment;T1:处理一, treatment 1;T2:处理二, treatment 2 | ||||||
模拟酸雨实验2年后, 对不同处理的土壤微生物群落采用DGGE技术检测, 确定了共30条DGGE带。结果表明, 模拟酸雨处理的所有样品中均含有8个DGGE谱带(3、7、11、13、14、23、28和30)。在这些条带中, 3、30条带被鉴定为细菌, 7、11、13、14、23、28条带被分别鉴定为放线菌、分枝杆菌、硝化螺、酸性微生物和鞘氨醇菌。样品中含有一些共有菌群, 如细菌、放线菌、厚壁菌门、硝化螺旋菌门和α-变形菌。酸雨也造成了某些菌群的减少或消失, 如带4(茎菌属)、9(孢囊杆菌属)、12(慢生根瘤菌属)和25(念珠菌属), 同时, 酸性条件也导致某些种类的增加, 如带5(肉食杆菌属)、6(丙酸杆菌属)、8(梭形芽孢杆菌属)。这说明在酸雨作用下这类微生物对土壤有机质降解和释放土壤二氧化碳中起着更大作用。
不同模拟酸雨处理的土壤细菌和土壤真菌多样性具有显著差异(表 3), 模拟酸雨处理的土壤细菌群落多样性表现为降低, 说明酸性胁迫抑制了土壤细菌群落的多样性。T2处理的土壤细菌多样性与CK处理相比存在显著差异(P < 0.05), 说明重度酸雨处理下土壤细菌群落对酸性环境的耐受性最低。而模拟酸雨处理的土壤真菌群落多样性表现相反, T1处理的土壤真菌多样性与CK处理和T2处理存在显著差异(P < 0.05), 说明中度酸雨处理条件下土壤真菌群落对酸性环境的耐受性最高, 弱酸可促进土壤真菌群落的多样性。
| 处理 Treatment |
土壤细菌Soil bacterial community | 土壤真菌Soil fungal community | |||||
| Shannon多样性指数 Shannon diversity index(H) |
Margalef丰富度指数 Margalef richness index (D) |
Pielou均匀度指数 Pielou evenness index(E) |
Shannon多样性指数 Shannon diversity index(H) |
Margalef丰富度指数 Margalef richness index(D) |
Pielou均匀度指数 Pielou evenness index(E) |
||
| CK | 3.28±0.03a | 3.54±0.02a | 0.998±0.015a | 2.22±0.04b | 2.29±0.03b | 0.999±0.014a | |
| T1 | 2.97±0.02ab | 3.16±0.06ab | 0.991±0.024a | 2.56±0.06a | 2.79±0.07a | 0.989±0.022a | |
| T2 | 2.79±0.05b | 2.88±0.03b | 0.993±0.013a | 2.24±0.02b | 2.15±0.05b | 0.992±0.025a | |
| 不同字母表示同一列内各均值存在显著差异(P < 0.05) | |||||||
土壤性质与土壤细菌、土壤真菌α多样性指数的相关性分析表明(表 4), 土壤细菌的Shannon多样性指数、Margalef丰富度指数与碱解氮和有效磷呈显著或极显著负相关(P < 0.01), 与pH值、速效钾、可溶性有机碳和微生物量碳呈显著或极显著正相关(P < 0.01);土壤真菌的Shannon多样性指数与碱解氮显著正相关(P < 0.05), 与可溶性有机碳和微生物量碳呈显著或极显著负相关(P < 0.01), Margalef丰富度指数与pH值、可溶性有机碳和微生物量碳呈显著负相关(P < 0.05), 与碱解氮为显著正相关(P < 0.05)。
| 多样性指数 Diversity index |
pH | 碱解氮 Available N |
有效磷 Available P |
速效钾 Available K |
可溶性有机碳 Dissolved organic carbon |
微生物量碳 Microbial biomass carbon |
|
| 土壤细菌 Soil bacteria |
Shannon多样性指数(H) | 0.720* | -0.817** | -0.676* | 0.627* | 0.730* | 0.771* |
| Margalef丰富度指数(D) | 0.834** | -0.873** | -0.709* | 0.686* | 0.801** | 0.889** | |
| Pielou均匀度指数(E) | 0.458 | -0.352 | 0.366 | 0.479 | 0.567 | -0.248 | |
| 土壤真菌 Soil fungi |
Shannon多样性指数(H) | -0.551 | 0.631* | 0.533 | -0.482 | -0.742* | -0.898** |
| Margalef丰富度指数(D) | -0.638* | 0.724* | 0.479 | -0.553 | -0.687* | -0.791* | |
| Pielou均匀度指数(E) | -0.332 | 0.103 | 0.325 | 0.417 | -0.257 | 0.228 | |
| *表示显著相关(P < 0.05), **表示极显著相关(P < 0.01) | |||||||
为了解释酸雨与土壤微生物群落结构及环境因子之间的关系, 对不同模拟酸雨样地土壤微生物群落与环境因子进行冗余分析, 可以看出CK与T1、T2之间具有明显地距离, 这证实了之前的分析, 即不同模拟酸雨处理样地的土壤微生物群落在差异很大程度上与环境变量有关。两轴累计贡献率为97.74%, T1和T2分布在第二排序轴正方向, CK分布在第二排序轴负方向, 表明不同模拟酸雨处理是第二排序轴的主要影响因素。pH、AK、DOC和MBC与模拟酸雨处理的土壤微生物群落结构关系紧密且正相关, AN、AP与土壤微生物群落结构负相关。排列检验显示pH、DOC、AP、AN和MBC对土壤细菌群落结构的影响最显著(P < 0.05)(图 3)。

|
| 图 3 土壤微生物群落与环境影响因子的冗余分析 Fig. 3 RDA of soil microbial community and environmental factors pH:酸碱度;AN:碱解氮, Available N;AP:有效磷, Available P;AK:速效钾, Available K;DOC:可溶性有机碳, Dissolved organic carbon;MBC:微生物量碳, Microbial biomass carbon |
土壤呼吸与土壤理化性质、土壤微生物α多样性指数的相关性分析表明(表 4), 土壤呼吸与pH值、速效钾、可溶性有机碳、微生物量碳和细菌Shannon指数呈显著正相关(P < 0.05), 与碱解氮和有效磷为显著负相关(P < 0.05)。
3 讨论 3.1 不同酸雨处理土壤呼吸的差异性及控制因子由于各地区的气候、土壤养分及植被类型等条件的不同, 模拟酸雨对土壤呼吸的影响结果存在较大差异。Reth等[22]研究发现模拟酸雨抑制了土壤呼吸, 这种抑制作用可能是氢离子高载荷对土壤微生物产生毒性所致。Fangueiro等[23]报道, 低模拟酸雨强度反而促进了土壤呼吸。本研究结果表明, 实验初期模拟酸雨对土壤呼吸无明显影响, 这主要是由于土壤拥有巨大的缓冲体系, 对酸性物质的输入具有很强的缓冲作用, 毛竹林根系短期内对酸雨的耐受性较强。随着时间推进, 在模拟酸雨作用下, 土壤呼吸呈现出先增大后减小的趋势, 酸雨的促进土壤呼吸作用可能是根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 那么根系呼吸也不会发生变化, 而酸雨中的NO3-的施肥作用恰恰增加了毛竹林根系细胞的氮含量, 使得地下的土壤呼吸速率加强[12]。模拟酸雨下森林的土壤酸化是个逐渐累积的过程, 随着酸雨强度的增大和时间的推移, 模拟酸雨对土壤呼吸产生抑制作用, 这与其胁迫下土壤酸化从而导致土壤微生物活性及土壤养分流失有关。相关性分析表明(表 4), 土壤呼吸与土壤细菌α多样性指数呈显著正相关(P < 0.05), 土壤微生物活性下降, 从而延缓了土壤有机物的矿化和分解速率, 抑制了土壤CO2的释放, 并呈现逐渐增强的趋势[24]。
| 项目 Project |
土壤呼吸 Soil respiration |
pH | 碱解氮 Available N |
有效磷 Available P |
速效钾 Available K |
可溶性有机碳 Dissolved organic carbon |
微生物量碳 Microbialbiomass carbon |
细菌多样性 Bacteria diversity |
真菌多样性 Fungi diversity |
| 土壤呼吸Soil respiration | 1 | ||||||||
| pH | 0.752* | 1 | |||||||
| 碱解氮Available N | -0.778* | -0.724* | 1 | ||||||
| 有效磷Available P | -0.783* | -0.755* | 0.777* | 1 | |||||
| 速效钾Available K | 0.773* | -0.893* | 0.422 | -0.735* | 1 | ||||
| 可溶性有机碳 Dissolved organic carbon |
0.741* | -0.748* | 0.767* | -0.750* | 0.776* | 1 | |||
| 微生物量碳 Microbial biomass carbon |
0.725* | 0.980** | -0.750* | -0.715* | 0.706* | 0.727* | 1 | ||
| 细菌多样性 Bacteria diversity |
0.739* | 0.720* | -0.817** | -0.676* | 0.627* | 0.730* | 0.771* | 1 | |
| 真菌多样性 Fungi diversity |
0.412 | 0.631* | 0.533 | -0.482 | -0.742* | -0.898** | 0.631* | 0.533 | 1 |
| *表示显著相关(P < 0.05), **表示极显著相关(P < 0.01) | |||||||||
酸雨对土壤微生物量碳的影响与酸雨胁迫下土壤持续酸化有关, 通常pH值较低的土壤中微生物量碳含量相应较低[25]。本研究在模拟酸雨的持续作用下, 土壤微生物量碳的含量及土壤呼吸速率呈显著下降特征, 这是由于酸雨导致土壤pH改变, 引起土壤微生物活性、功能及土壤微生物量碳的变化。该结果与王国兵等[26]研究结论一致, 土壤呼吸产生的CO2部分是由于微生物的自养呼吸和分解有机质和植物凋落物的异养呼吸而产生的, 微生物生物量碳的减少间接地导致了土壤呼吸速率的减少[27]。另外, 酸雨使土壤pH值降低, 有机质矿化速率下降, 土壤腐殖质层DOC的释放量减少[28]。本研究中毛竹林土壤溶解性有机碳呈随着酸雨强度增加而下降的趋势, 可能由于DOC来源于凋落物的分解以及微生物代谢, 微生物活减少, 产生的水溶性有机物质也减少[29-30]。因此, 酸雨对土壤微生物活性的影响被认为是影响DOC释放的关键因素。
土壤温度是影响土壤呼吸季节性动态变化的最主要的因素, 土壤呼吸为酶促反应, 而土壤温度是影响酶促反应的重要因素, 温度升高往往会导致酶促反应加快, 从而促进土壤呼吸。本研究表明, 酸雨各处理的土壤呼吸季节变化具有明显规律性, 在不受土壤水分胁迫的条件下酸雨各处理的土壤呼吸与土壤温度之间遵循指数回归关系, 这与Nikolova等[31]以及Karhu等[32]的研究结果一致。梁国华等[33]发现酸雨降低了土壤呼吸的温度敏感性, 并且酸性越强抑制作用越显著。Chen等[34]研究表明酸雨对土壤呼吸的温度敏感性没有影响。本研究发现模拟酸雨改变了土壤呼吸对温度的响应, 表现为Q10值在酸处理下有上升的趋势, 说明模拟酸雨增加了土壤呼吸的温度敏感性, 这可能是因为DOC随酸处理的增加而减少, 不同生态系统土壤呼吸季节变异的Q10与DOC呈负相关[35]。毛竹林生态系统中毛竹林凋落物分解因酸雨而促进, 凋落物有机碳的质量逐渐下降, 土壤呼吸的Q10值越来越高[36]。Q10值是在整个测定期间进行计算的, 分高温阶段或低温不同阶段分析可能有更为具体直观的结果, 今后可进一步研究酸雨胁迫下土壤呼吸在不同温度时段的温度敏感性, 以及进一步测定土壤呼吸的日变化也是今后研究值得考虑的方面。
3.2 不同模拟酸雨条件下土壤微生物群落多样性的比较及控制因子土壤呼吸与土壤有机物的转化与循环密切相关, 而土壤有机物的转化与循环有赖于微生物活化才能完成。酸化的土壤由于H+的毒害作用, 从而改变了土壤分解者微生物的种类, 结构以及生物活性[37]。不同pH的模拟酸雨均能抑制细菌、放线菌、氮素生理类群的生长, 其数量随着酸雨pH的降低而不断减少, 真菌、纤维素分解菌的数量随酸雨pH的降低呈现先升高后下降的趋势[38]。本研究结果表明, 模拟酸雨对毛竹林土壤细菌和真菌群落多样性、丰富度具有显著影响, 通过比较指数值的大小可以发现, 酸雨能够在一定程度上降低土壤细菌群落多样性, 而弱酸能够显著提高土壤细菌群落多样性, 同时对酸雨具有抗性或耐性的微生物种群会表现出更高的活性。
有关不同地区土壤微生物多样性的研究表明, 土壤pH值是影响土壤微生物多样性的重要因子[39]。Pearson相关性分析表明, 土壤细菌的多样性指数和丰富度指数与pH值呈显著或极显著正相关, 这与孙岩和全炳武[40]的研究结果一致, 这可能是酸雨胁迫会抑制大多数细菌的生长和繁殖, 使土壤细菌的多样性和丰富度下降。同时, 我们利用冗余分析可以揭示不同酸雨处理条件下土壤性质对微生物结构的影响, 发现其结果与土壤细菌相近, 这也说明土壤细菌群落是毛竹林土壤环境中的主导微生物。本研究中, 土壤微生物与pH值、有效钾、可溶性有机碳和微生物量碳呈显著正相关, 同时表现出与土壤呼吸呈正相关关系。这可能是与南方酸性土壤有关, 酸雨淋溶增强, 土壤的分解与矿化过程较快, 土壤元素的迁移较强, 尤其是钾的流失量较大, 土壤含钾量低, 抑制土壤微生物的生长。土壤微生物与土壤碱解氮和有效磷含量呈显著负相关关系, 可能原因pH值降低, 土壤对氮、磷酸根的吸附量降低。酸雨对土壤生态系统的影响是个长期累积的过程, 开展酸雨胁迫下毛竹林土壤呼吸的长期定位观测具有较高研究价值, 尽管本次研究经历两年的实验, 但酸雨处理区域面积偏小, 今后可增大实验区面积, 以更好地深入研究酸雨胁迫引起的土壤酸化对土壤生态系统过程影响, 这些工作开展将有利于我国毛竹林的科学经营及亚热带常绿阔叶林的生态保护。
4 结论(1) 模拟酸雨对土壤呼吸速率的影响表现为先促进后抑制趋势, 土壤呼吸速率呈季节性规律变化, 不同模拟酸雨处理改变了土壤呼吸对温度的敏感性。
(2) 模拟酸雨改变土壤微生物群落结构及多样性, 重度酸雨处理抑制了土壤细菌群落生长, 而中度酸雨对土壤真菌多样性和丰富度具有促进作用。
(3) pH值、速效钾、可溶有机碳等土壤理化性质与土壤微生物群落结构、土壤呼吸密切相关。
| [1] |
Menz F C, Seip H M. Acid rain in Europe and the United States:an update. Environmental Science & Policy, 2004, 7(4): 253-265. |
| [2] |
Zhang X Y, Jiang H, Jin J X, Xu X H, Zhang Q X. Analysis of acid rain patterns in northeastern China using a decision tree method. Atmospheric Environment, 2012, 46: 590-596. |
| [3] |
牟永铭, 朱光良. 基于GIS技术的浙江省酸雨区分布研究. 科技通报, 2005, 21(3): 356-359. |
| [4] |
Ramlall C, Varghese B, Ramdhani S, Pammenter N W, Bhatt A, Berjak P, Sershen. Effects of simulated acid rain on germination, seedling growth and oxidative metabolism of recalcitrant-seeded Trichilia dregeana grown in its natural seed bank. Physiologia Plantarum, 2015, 153(1): 149-160. |
| [5] |
许第发, 钟天祥, 徐冬梅, 刘广深. 酸沉降对土壤地球化学过程的影响. 地质地球化学, 2002, 30(4): 68-74. |
| [6] |
刘源月, 江洪, 李雅红, 原焕英. 模拟酸雨对杉木幼苗-土壤复合体系土壤呼吸的短期效应. 生态学报, 2010, 30(8): 2010-2017. |
| [7] |
Liang G H, Liu X Z, Chen X M, Qiu Q Y, Zhang D Q, Chu G W, Liu S X, Liu S Z, Zhou G Y. Response of soil respiration to acid rain in forests of different maturity in southern china. PLoS One, 2013, 8(4): e62207. |
| [8] |
Mitchell P J, Simpson A J, Soong R, Simpson M J. Shifts in microbial community and water-extractable organic matter composition with biochar amendment in a temperate forest soil. Soil Biology and Biochemistry, 2015, 81: 244-254. |
| [9] |
Davidson E A, Savage K, Verchot L V, Navarro R. Minimizing artifacts and biases in chamber-based measurements of soil respiration. Agricultural and Forest Meteorology, 2002, 113(1/4): 21-37. |
| [10] |
Janssens I A, Lankreijer H, Matteucci G, Kowalski A S, Buchmann N, Epron D, Pilegaard K, Kutsch W, Longdoz B, Grünwald T, Montagnani L, Dore S, Rebmann C, Moors E J, Grelle A, Rannik V, Morgenstern K, Oltchev S, Clement R, Guðmundsson J, Minerbi S, Berbigier P, Ibrom A, Moncrieff J, Aubinet M, Bernhofer C, Jensen N O, Vesala T, Granier A, Schulze E D, Lindroth A, Dolman A J, Jarvis P G, Ceulemans R, Valentini R. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Global Change Biology, 2001, 7(3): 269-278. |
| [11] |
Chen S T, Shen X S, Hu Z H, Chen H S, Shi Y H, Liu Y. Effects of simulated acid rain on soil CO2 emission in a secondary forest in subtropical China. Geoderma, 2012, 189-190: 65-71. |
| [12] |
陈书涛, 孙鹭, 桑琳, 胡正华. 模拟酸雨对次生林土壤呼吸及异养呼吸的影响. 环境科学, 2017, 38(3): 1235-1244. |
| [13] |
Li P H, Zhou G M, Du H Q, Lu D S, Mo L F, Xu X J, Shi Y J, Zhou Y F. Current and potential carbon stocks in Moso bamboo forests in China. Journal of Environmental Management, 2015, 156: 89-96. |
| [14] |
Zhou G M, Xu X J, Du H Q, Ge H L, Shi Y J, Zhou Y F. Estimating aboveground carbon of Moso bamboo forests using the k nearest neighbors technique and satellite imagery. Photogrammetric Engineering and Remote Sensing, 2011, 77(11): 1123-1131. |
| [15] |
Du H Q, Zhou G M, Ge H L, Fan W Y, Xu X J, Fan W L, Shi Y J. Satellite-based carbon stock estimation for bamboo forest with a non-linear partial least square regression technique. International Journal of Remote Sensing, 2012, 33(6): 1917-1933. |
| [16] |
白尚斌, 周国模, 王懿祥, 梁倩倩, 陈娟, 程艳艳, 沈蕊. 天目山保护区森林群落植物多样性对毛竹入侵的响应及动态变化. 生物多样性, 2013, 21(3): 288-295. |
| [17] |
李雅红, 江洪, 原焕英, 刘源月, 周国模, 余树全. 西天目山毛竹林土壤呼吸特征及其影响因子. 生态学报, 2010, 30(17): 4590-4597. |
| [18] |
杨文佳, 李永夫, 姜培坤, 周国模, 刘娟. 亚热带毛竹人工林土壤呼吸组分动态变化及其影响因素. 应用生态学报, 2015, 26(10): 2937-2945. |
| [19] |
武志华, 夏冬双, 王雪寒, 马强, 唐凯, 刘惠荣. 利用PCR-DGGE技术分析内蒙古西部地区土壤细菌的多样性. 生态学报, 2019, 39(7): 2545-2557. |
| [20] |
Davidson E A, Janssens I A, Luo Y Q. On the variability of respiration in terrestrial ecosystems:moving beyond Q10. Global Change Biology, 2006, 12(2): 154-164. |
| [21] |
赵天心, 毛新伟, 程敏, 陈俊辉, 秦华, 李永春, 梁辰飞, 徐秋芳. 毛竹种植对土壤细菌和真菌群落结构及多样性的影响. 应用生态学报, 2017, 28(11): 3740-3750. |
| [22] |
Reth S, Reichstein M, Falge E. The effect of soil water content, soil temperature, soil pH-value and the root mass on soil CO2 efflux-A modified model. Plant and Soil, 2005, 268(1/2): 21-33. |
| [23] |
Fangueiro D, Surgy S, Coutinho J, Vasconcelos E. Impact of cattle slurry acidification on carbon and nitrogen dynamics during storage and after soil incorporation. Journal of Plant Nutrition and Soil Science, 2013, 176(4): 540-550. |
| [24] |
Walna B, Siepak J, Drzymala S. Soil degradation in the Wielkopolski National Park (Poland) as an effect of acid rain simulation. Water, Air, and Soil Pollution, 2001, 130(1): 1727-1732. |
| [25] |
Araújo A S F, Monteiro R T R. Microbial biomass and activity in a Brazilian soil amended with untreated and composted textile sludge. Chemosphere, 2006, 64(6): 1043-1046. |
| [26] |
王国兵, 郝岩松, 王兵, 阮宏华. 土地利用方式的改变对土壤呼吸及土壤微生物生物量的影响. 北京林业大学学报, 2006, 28(S2): 74-79. |
| [27] |
Lim S M, Cha S S, Shim J K. Effects of simulated acid rain on microbial activities and litter decomposition. Journal of Ecology and Environment, 2011, 34(4): 401-410. |
| [28] |
Lu S B, Chen C R, Zhou X Q, Xu Z H, Bacon G, Rui Y C, Guo X M. Responses of soil dissolved organic matter to long-term plantations of three coniferous tree species. Geoderma, 2012, 170: 136-143. |
| [29] |
Huang J S, Hu B, Qi K B, Chen W J, Pang X Y, Bao W K, Tian G L. Effects of phosphorus addition on soil microbial biomass and community composition in a subalpine spruce plantation. European Journal of Soil Biology, 2016, 72: 35-41. |
| [30] |
张慧玲, 吴建平, 熊鑫, 褚国伟, 周国逸, 张德强. 南亚热带森林土壤碳库稳定性与碳库管理指数对模拟酸雨的响应. 生态学报, 2018, 38(2): 657-667. |
| [31] |
Nikolova P S, Raspe S, Andersen C P, Mainiero R, Blaschke H, Matyssek R, Häberle K H. Effects of the extreme drought in 2003 on soil respiration in a mixed forest. European Journal of Forest Research, 2009, 128(2): 87-98. |
| [32] |
Karhu K, Fritze H, Tuomi M, Vanhala P, Spetz P, Kitunen V, Liski J. Temperature sensitivity of organic matter decomposition in two boreal forest soil profiles. Soil Biology and Biochemistry, 2010, 42(4): 72-82. |
| [33] |
梁国华, 吴建平, 熊鑫, 吴小映, 褚国伟, 周国逸, 曾任森, 张德强. 模拟酸雨对鼎湖山季风常绿阔叶林土壤呼吸的初期影响. 广西植物, 2016, 36(2): 145-153. |
| [34] |
Chen S T, Zhang X, Liu Y F, Hu Z H, Shen X S, Ren J Q. Simulated acid rain changed the proportion of heterotrophic respiration in soil respiration in a subtropical secondary forest. Applied Soil Ecology, 2015, 86: 148-157. |
| [35] |
Zheng Z M, Yu G R, Fu Y L, Wang Y S, Sun X M, Wang Y H. Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content:a trans-China based case study. Soil Biology and Biochemistry, 2009, 41(7): 1531-1540. |
| [36] |
Jia B R, Zhou G S, Wang F Y, Wang Y H, Weng E S. Effects of grazing on soil respiration of Leymus chinensis steppe. Climatic Change, 2007, 82(1): 211-223. |
| [37] |
Xu H Q, Zhang J E, Qu Y, Lin L, Quan G M, Zhao B L, Yu J Y. Effects of simulated acid rain on microbial characteristics in a lateritic red soil. Environmental Science and Pollution Research, 2015, 22(22): 18260-18266. |
| [38] |
Chen D M, Wang Y, Lan Z C, Li J J, Xing W, Hu S J, Bai Y F. Biotic community shifts explain the contrasting responses of microbial and root respiration to experimental soil acidification. Soil Biology and Biochemistry, 2015, 90: 139-147. |
| [39] |
Chu H Y, Fierer N, Lauber C L, Caporaso J G, Knight R, Grogan P. Soil bacterial diversity in the Arctic is not fundamentally different from that found in other biomes. Environmental Microbiology, 2010, 12(11): 2998-3006. |
| [40] |
孙岩, 全炳武. 模拟酸雨对玉米幼苗生理与根际微生物的影响. 延边大学农学学报, 2009, 31(3): 165-171. |