 2020, Vol. 40
2020, Vol. 40文章信息
- 唐志红, 尉秋实, 刘虎俊, 姜生秀, 何芳兰, 张莹花, 王芳琳, 张裕年, 赵赫然, 赵鹏
- TANG Zhihong, YU Qiushi, LIU Hujun, JIANG Shengxiu, HE Fanglan, ZHANG Yinghua, WANG Fanglin, ZHANG Yunian, ZHAO Heran, ZHAO Peng
- 祁连山东段高寒植被群落特征及其与地形气候因子关系研究
- Characteristics of alpine vegetation community and its relationship to topographic climate factors in the eastern Qilian mountain
- 生态学报. 2020, 40(1): 223-232
- Acta Ecologica Sinica. 2020, 40(1): 223-232
- http://dx.doi.org/10.5846/stxb201812192760
-
文章历史
- 收稿日期: 2018-07-10
- 网络出版日期: 2019-10-25





2. 甘肃省治沙研究所, 甘肃省荒漠化与风沙灾害防治重点实验室-省部共建国家重点实验室培育基地, 兰州 730070;
3. 甘肃民勤荒漠草地生态系统国家野外科学观测研究站, 民勤 733000;
4. 甘肃河西走廊森林生态系统国家定位观测研究站, 武威 733000
2. State Key Laboratory Breeding Base of Desertification and Aeolian Sand Disaster Combating, Gansu Desert Control Research Institute, Lanzhou 730070, China;
3. Gansu Minqin National Field Observation & Research Station on Ecosystem of Desert Grassland, Minqin 733000, China;
4. Gansu Hexi Corridor Forest Ecosystem National Research Station, Wuwei 733000, China
山地植被空间分布格局是气候和地形影响的直接结果, 也是山地生态学研究的主要内容[1]。生境和气候带的多样使我国成为北半球最富植物多样性的国家之一[2]。地形是植被时空异质性的重要来源, 山地森林植被存在多重尺度和方向的分异格局[3]。海拔、坡度、坡向等地形因子通过对热量、光照辐射、水分等资源的再分配影响着山地植物种的分布范围[4]、群落动态[5]、植被类型[6], 其中海拔是最重要的影响因子[7-8]。作为植物多样性的演化和维持的重要因素之一[9], 气候通过局域生物因素(如盖度、生活型)间接作用于多样性格局[10]。山地生态系统森林植物种的空间分布对气候变化的响应十分复杂, 尤其在降雨量小于400 mm的干旱区。青藏高原树线位置主要受7月最低气温的驱动, 且受到坡向的影响[11]。降雨量是影响祁连山青海云杉径向生长的最重要因子[12]。能量和水分共同作用是影响大尺度森林乔木层物种多样性空间分布格局[13]、潜在自然植被NPP [14]、植被覆盖[15]的主要因素。干旱区森林植物群落的分布取决于降水、温度等气候因子, 同时也受局地生境和地形空间异质性的制约, 准确理解气候地形因子对群落结构的共同影响机制, 有助于在变化环境条件下预测植物群落的空间分布[16], 实现山地物种多样性变化与模拟[17]。
祁连山东段高寒植被群落位于石羊河上游中、浅山地带, 其水源涵养功能对于改善整个流域水资源状况、维持流域生态平衡具有重要作用。目前祁连山植被群落的研究集中在高寒草甸[18], 高山灌丛[19], 青海云杉[20, 21]等单一植被群落, 且空间范围以北坡居多[22-25], 多以遥感数据源为主[26-27], 关于祁连山东段植被群落特征及其与地形气候因子关系的研究未见报道。为此, 本研究基于大范围野外植被样方调查和地形气候数据资料, 旨在摸清高寒植被群落的物种组成, 阐明主要群丛类型及其分布格局, 揭示地形气候因子对物种分布的驱动机制, 为祁连山东段高寒植被群落物种多样性保护提供理论依据。
1 材料与方法 1.1 研究区概况研究区位于祁连山中东段的哈溪-祁连-大黄山地区, 地理位置为纬度37°22′13.4″—38°25′52.9″, 经度102°15′42.9″—103°14′48.5″。该区域属高寒半干旱气候区, 因受大陆性荒漠气候和高大山体的影响, 气候的水平、垂直性差异较大。地势西南高、东北低, 海拔在2400—4146 m之间。年平均气温1.5℃, 年均降水量约400 mm。森林类型构成简单, 主要乔木树种有青海云杉Picea crassifolia、祁连圆柏Sabina chinensis, 局部有落叶松Larix gmelinii、山杨Populus davidiana等。灌木有金露梅Potentilla fruticosa、银露梅Potentilla glabra、鬼箭锦鸡儿Caragana jubata 、唐古特瑞香Daphne tangutica等。草本植物以蓼科、禾本科、毛茛科、龙胆科等科植物居多。
1.2 样地布设与调查2013年7月—8月在祁连山北坡东段哈溪-祁连-大黄山一带选择典型植被群落类型设置样地(图 1), 坡面植被根据海拔设定上、中、下, 3个梯度, 每个梯度沿等高线布设3个20 m×20 m乔木大样方。乔木样方内沿对角线布设3个5 m×5 m灌木样方和随机布设5个1 m×1 m草本样方。调查统计样方内乔木层的种类、胸径、树高、冠幅, 多度;灌木层的种类、高度、盖度、多度;草本层种类、高度、多度、盖度。共调查乔木样方65个, 灌木样方195个, 草本样方325个。

|
| 图 1 植物群落样地调查点 Fig. 1 Overview of plant samples sites distribution in Qilian Mountains |
采用GPS记录样地的经纬度、海拔。基于Climatic Research Unit (CRU), 即英国东英格利亚大学气候研究所1901—2012年月平均地表 0.5°×0.5°气候要素数据集(http://www.geodata.cn)和DEM数据(http://www.gscloud.cn/), 借助Arcgis10.2软件Spatial analyst工具下Extraction功能, 参考样地经纬度获得降水、温度、坡度和坡向数据。
1.3 数据处理(1) 重要值
作为各种植物在群落中的优势度指标, 重要值表示不同植物在群落中的功能地位, 计算公式如下:
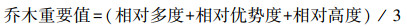
|
(1) |
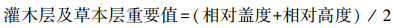
|
(2) |
(2) 群落分类与排序
采用双向指示种分析(Two-way indicator species analysis, TWINSPAN)对高山植被群落进行群丛类型划分, 选择0、2、5、10、20共5个分类层次水平, 剔除出现次数小于2的植物种, 分类软件采用WINTWIN 2.3。DCA(Detrend Correspondence Analysis)及CCA(Canonical Correspondence Analysis)排序分析采用国际标准生态学软件Canoco 5, 排序分析方法如下:先对物种重要值-样方数据进行除趋势对应分析, 根据DCA排序轴的梯度长度选择适宜的排序方法。一般排序轴梯度长度<3时, 宜采用线性模型排序;梯度长度> 4时, 宜采用单峰模型排序;梯度长度介于3和4之间, 两种模型都适合。排序前采用前项选择对所环境因子进行RDA筛选, 若某环境因子的变异膨胀因子> 20, 表明与其他环境因子有多重共线性, 对模型的贡献很小可以舍去。
2 结果 2.1 植被群落物种组成野外调查共记录种子植物181种, 隶属40科, 124属。科物种组成及占总物种数比例分别为菊科30个种, 占16.57%;蔷薇科17个, 占9.44%;禾本科13个种, 占7.22%;豆科11个种, 占6.11%。毛茛科10个种, 占5.56%。单属种82个, 占总属数的66.13%。植被群落层片由乔木层、灌木层和草本层组成, 乔木8种, 灌木25种, 草本148种。根据表 1群落不同层片物种重要值排序可知, 乔木层优势种有青海云杉、祁连圆柏、红桦、落叶松、山杨、白桦。灌木层优势种有金露梅、山生柳、匙叶小檗、高山绣线菊、沙棘。草本层优势种有甘肃薹草、珠芽蓼、早熟禾、唐松草、甘青蒿(表 1)。
| 科名 Family |
属名 Genus |
种名 Species |
生活型 Life form |
重要值 Important value |
| 松科 | 云杉属 | 青海云杉Picea crassifolia | 乔木 | 718.5 |
| 柏科 | 圆柏属 | 祁连圆柏Sabina chinensis | 乔木 | 159.07 |
| 桦木科 | 桦木属 | 红桦Betula albosinensis | 乔木 | 60.2 |
| 松科 | 落叶松属 | 落叶松Larix gmelinii | 乔木 | 44.87 |
| 杨柳科 | 杨属 | 山杨Populus davidiana | 乔木 | 43.84 |
| 桦木科 | 桦木属 | 白桦Betula platyphylla | 乔木 | 42.44 |
| 芸香科 | 花椒属 | 花椒Zanthoxylum bungeanum | 乔木 | 15.49 |
| 松科 | 松属 | 油松Pinus tabuliformis | 乔木 | 4.25 |
| 蔷薇科 | 委陵菜属 | 金露梅Potentilla fruticosa | 灌木 | 131.94 |
| 杨柳科 | 柳属 | 山生柳Salix oritrepha | 灌木 | 114.59 |
| 小檗科 | 小檗属 | 匙叶小檗Berberis vernae | 灌木 | 113.01 |
| 蔷薇科 | 绣线菊属 | 高山绣线菊Spiraea alpina | 灌木 | 109.79 |
| 胡颓子科 | 沙棘属 | 沙棘Hippophae rhamnoides | 灌木 | 105.05 |
| 蔷薇科 | 委陵菜属 | 银露梅Potentilla glabra | 灌木 | 69.93 |
| 豆科 | 锦鸡儿属 | 鬼箭锦鸡儿Caragana jubata | 灌木 | 43.7 |
| 瑞香科 | 瑞香属 | 唐古特瑞香Daphne tangutica | 灌木 | 42.53 |
| 蔷薇科 | 委陵菜属 | 小叶金露梅Potentilla parvifolia | 灌木 | 42.34 |
| 杜鹃花科 | 杜鹃属 | 烈香杜鹃Rhododendron anthopogonoides | 灌木 | 38.17 |
| 蔷薇科 | 蔷薇属 | 红花蔷薇Rosa moyesii | 灌木 | 26.69 |
| 豆科 | 锦鸡儿属 | 狭叶锦鸡儿Caragana stenophylla | 灌木 | 26.02 |
| 杜鹃花科 | 杜鹃属 | 千里香杜鹃Rhododendron thymifolium | 灌木 | 21.96 |
| 杜鹃花科 | 杜鹃属 | 头花杜鹃Rhododendron capitatum | 灌木 | 21.69 |
| 杜鹃花科 | 杜鹃属 | 青海杜鹃Rhododendron qinghaiense | 灌木 | 20.25 |
| 蔷薇科 | 栒子属 | 红花栒子Cotoneaster rubens | 灌木 | 17.65 |
| 蔷薇科 | 蔷薇属 | 密刺蔷薇Rosa spinosissima | 灌木 | 16.94 |
| 蔷薇科 | 栒子属 | 黑果栒子Cotoneaster melanocarpus | 灌木 | 12.79 |
| 蔷薇科 | 悬钩子属 | 紫色悬钩子Rubus irritans | 灌木 | 11.62 |
| 柏科 | 圆柏属 | 叉子圆柏Sabina vulgaris | 灌木 | 10.82 |
| 杜鹃花科 | 北极果属 | 北极果Arctous alpinus | 灌木 | 9.76 |
| 蔷薇科 | 栒子属 | 西北栒子Cotoneaster zabelii | 灌木 | 8.55 |
| 虎耳草科 | 茶藨子属 | 长果茶藨子Ribes stenocarpum | 灌木 | 8.28 |
| 忍冬科 | 忍冬属 | 忍冬Lonicera japonica | 灌木 | 8.26 |
| 小檗科 | 小檗属 | 黄芦木Berberis amurensis | 灌木 | 5.36 |
| 莎草科 | 薹草属 | 甘肃薹草Carex kansuensis | 多年生草本 | 660.09 |
| 蓼科 | 蓼属 | 珠芽蓼Polygonum viviparum | 多年生草本 | 434.90 |
| 禾本科 | 早熟禾属 | 早熟禾Poa annua | 一年生草本 | 218.06 |
| 毛茛科 | 唐松草属 | 唐松草Thalictrum aquilegiifolium var. sibiricum | 多年生草本 | 112.62 |
| 菊科 | 蒿属 | 甘青蒿Artemisia tangutica | 多年生草本 | 104.90 |
| 莎草科 | 球柱草属 | 丝叶球柱草Bulbostylis densa | 一年生草本 | 104.21 |
| 毛茛科 | 乌头属 | 高乌头Aconitum sinomontanum | 多年生草本 | 103.04 |
| 蔷薇科 | 草莓属 | 东方草莓Fragaria orientalis | 多年生草本 | 99.02 |
| 菊科 | 火绒草属 | 火绒草Leontopodium leontopodioides | 多年生草本 | 94.23 |
| 禾本科 | 披碱草属 | 垂穗披碱草Elymus nutans | 多年生草本 | 88.60 |
| 毛茛科 | 银莲花属 | 小花草玉梅Anemone rivularis | 多年生草本 | 82.67 |
| 玄参科 | 马先蒿属 | 藓生马先蒿 Pedicularis muscicola | 多年生草本 | 80.72 |
| 菊科 | 苦荬菜属 | 苦荬菜Ixeris polycephala | 多年生草本 | 78.47 |
| 堇菜科 | 堇菜属 | 堇菜Viola verecunda | 多年生草本 | 73.89 |
| 菊科 | 风毛菊属 | 风毛菊Saussurea japonica | 二年生草本 | 73.01 |
| 荨麻科 | 冷水花属 | 冷水花Pilea notata | 多年生草本 | 65.88 |
| 堇菜科 | 堇菜属 | 双花堇菜Viola biflora | 多年生草本 | 60.63 |
| 伞形科 | 独活属 | 裂叶独活Heracleum millefolium | 多年生草本 | 55.28 |
| 毛茛科 | 毛茛属 | 高原毛茛Ranunculus tanguticus | 多年生草本 | 51.55 |
| 菊科 | 风毛菊属 | 小花凤毛菊Saussurea parviflora | 多年生草本 | 51.17 |
| 豆科 | 棘豆属 | 黄花棘豆Oxytropis ochrocephala | 多年生草本 | 50.60 |
| 鸭跖草科 | 鸭跖草属 | 节节草Commelina diffusa | 一年生草本 | 48.33 |
| 禾本科 | 针茅属 | 针茅Stipa capillata | 多年生草本 | 45.59 |
| 禾本科 | 芨芨草属 | 醉马草 Achnatherum inebrians Keng | 多年生草本 | 45.08 |
| 菊科 | 橐吾属 | 箭叶橐吾Ligularia sagitta | 多年生草本 | 41.75 |
| 物种重要值系65个样地的累加, 包括乔木层、灌木层及草本重要值排序的前25个种 | ||||
采用TWINSPAN第三层次水平的划分结果, 依据群落生境特征的指示种和优势种命名群丛类型, 最终得到高寒植被植物群落7个群丛(图 2):

|
| 图 2 祁连山植被群落双向指示种分析树状图 Fig. 2 Dendrogram with the Two-way indicator species analysis for the plant communities in Qilian mountain N(Number)代表样方的数量,Number of samples;Eig代表特征值Eigenvalue, |
群丛Ⅰ红桦-红花蔷薇-甘肃薹草, 包括样方54、55、56、57, 伴生种有白桦、银露梅、垂穗披碱草、扁蕾、凤毛菊、小花草玉梅、唐松草、高乌头, 平均海拔2532 m, 群丛层片由乔木层、灌木层和草本层组成, 主要分布在朱岔峡二甲皮。
群丛Ⅱ青海云杉-匙叶小檗-甘肃薹草, 包括样方9、10、13、22、23、27、28、29、30、31、37、38、39、41、42、51、52、62、63、64、65, 伴生种有高山绣线菊、银露梅、密刺蔷薇、红花蔷薇、长果茶藨子、西北栒子、唐古特瑞香、小叶金露梅、紫色悬钩子、珠芽蓼、早熟禾、高乌头、堇菜、藓生马先蒿、甘肃马先蒿、小花凤毛菊、拉拉藤、扁蕾、小花草玉梅、唐松草、双花堇菜、紫苞风毛菊、黄耆、羌活、黄精、六叶葎、裂叶独活, 海拔2662 m, 群丛层片由乔木层、灌木层和草本层组成, 主要分布在哈溪猫坨落、清水沟、直河沟, 祁连西沟河大山阳洼、林缘造林区。
群丛Ⅲ祁连圆柏-高山绣线菊-珠芽蓼, 包括样方14、15、16、17、20、21、24、25、26、32、33、34、40、46、47、48、49、50、53, 伴生种有青海云杉、山生柳、金露梅、银露梅、匙叶小檗、唐古特瑞香、小叶金露梅、早熟禾、中国马先蒿、球穗藨草、高山紫菀、黄花棘豆、堇菜、高原毛茛、东方草莓、裂叶独活、藓生马先蒿、双花堇菜、龙胆、肉果草、针茅、小花草玉梅、蒲公英、甘肃薹草、扁蕾、小米草, 海拔2917 m, 群丛层片由乔木层、灌木层和草本层组成, 主要分布在哈溪双龙沟、慢坡垭豁、火烧台、清水沟口, 祁连甘沙河锅层掌、天池。
群丛Ⅳ高山绣线菊-珠芽蓼+青藏蓼, 包括样方35、36、43、44、45、58、59、60、61, 伴生种有鬼箭锦鸡儿、山生柳、青海杜鹃、千里香杜鹃、头花杜鹃、火绒草、甘肃薹草、早熟禾、湿地繁缕、矮大黄、大花嵩草、唐松草、高原毛茛、丝叶球柱草、垂穗披碱草, 海拔3332 m, 群丛层片由灌木层和草本层组成, 主要分布在古城五台岭、祁连冰沟河、西沟河大山阳洼上坡、半阳坡。
群丛Ⅴ沙棘+甘青蒿+鼠掌老鹳草, 包括样方3、5、6、7、8、11、12, 伴生种有丝叶球柱草、箭叶橐吾、拂子茅、早熟禾、披碱草、甘露子、节节草、赖草、小米草、苦荬菜, 群丛层片由灌木层和草本层组成, 海拔2703 m, 主要分布在哈溪大南泥沟退耕地中上坡。
群丛Ⅵ沙棘+苦荬菜+苦荞麦, 包括样方1、2、4, 伴生种有风毛菊、蕨麻、甘青蒿、甘露子、羊茅、丝叶球柱草、拉拉藤, 群丛层片由灌木层和草本层组成, 海拔2527 m, 主要分布在哈溪大南泥沟退耕地坡底。
群丛Ⅶ沙棘-冰草+西北沼委陵菜, 包括样方18、19, 伴生种有金露梅、垂穗披碱草、湿地繁缕、芥叶蒲公英、平车前, 群丛层片由灌木层和草本层组成, 海拔2917 m, 主要分布在哈溪双龙沟河沟砾石地。
2.3 植被群落的排序65个样方-重要值的DCA分析结果表明, 4个排序轴的特征值分别为0.53、0.28、0.20、0.16, 因其前两个排序轴的特征值较大, 显示出重要的生态意义, 所以采用前2个排序轴的数据分别作二维排序图(图 3)。TWINSPAN分类群丛类型在DCA排序图上的聚集分布, 反映了较好的环境梯度。群丛Ⅰ海拔2532 m至群丛Ⅳ海拔3332 m沿DCA第二轴由下往上依次分布, 反映了海拔的梯度变化。群丛Ⅴ和Ⅵ交叉分布在排序图左端上中位置, 群丛Ⅶ沙棘-冰草+西北沼委陵菜单独分布在排序图左上角, 代表哈溪双龙沟河沟砾石地。

|
| 图 3 祁连山植被群落样方的DCA排序图 Fig. 3 DCA two-dimensional ordination diagram for samples of plant community in Qilian mountain DCA,即除趋势对应分析,Detrend Correspondence Analysis; S1—S65代表样品编号,S1—S65 represents sample number。 |
祁连山高寒植被群落物种重要值-样方矩阵的DCA排序所有轴的梯度长度最大为4.6, 故选择单峰模型(CCA)。蒙特卡罗检验(Monte Carlo)结果显示, 所有典范排序轴通过统计检验(F = 1.8, P = 0.002), 说明所选环境因子对祁连山高寒植被群落植物种分布具有显著影响。CCA排序前4轴代表不同的环境因子梯度变化, 根据环境因子与排序轴的相关性大小, 海拔、降水、坡度与坡向第一轴呈正相关关系(P<0.05), 相关性大小排序为海拔>降水>坡向, 表明第一轴主要反映海拔的梯度变化。降水、海拔、坡度与第二轴呈正相关关系(P <0.05), 相关性大小排序为降水>海拔>坡度, 表明第二轴主要反映降水的梯度变化(表 2)。前向选择与蒙特卡罗检验(Monte Carlo)检验结果表明, 海拔是祁连山高寒植被群落植物种分布的最重要环境因子, 其次是降水、温度、坡向、坡度(表 3)。
| 排序轴 Axis |
第Ⅰ轴 AxisⅠ |
第二轴 Axis Ⅱ |
第三轴 Axis Ⅲ |
第四轴 Axis Ⅳ |
| 环境因子Environmental variables | ||||
| 坡向Aspect | 0.27* | 0.13 | 0.55** | 0.02 |
| 坡度Slope | 0.43** | 0.28* | 0.50** | 0.47** |
| 海拔Altitude | 0.89** | 0.36** | 0.22 | 0.14 |
| 降水Precipitation | 0.48** | 0.85** | 0.10 | 0.03 |
| 温度Temperature | 0.19 | 0.06 | 0.18 | 0.95** |
| 特征值Eigenvalues | 0.28 | 0.15 | 0.11 | 0.08 |
| 物种-环境相关性Species-environment correlations | 0.87 | 0.85 | 0.70 | 0.73 |
| 物种数据累计百分比方差 Cumulative percentage variance of species |
5.48 | 8.35 | 10.43 | 12.02 |
| 物种-环境因子关系数据累计百分比方差 Cumulative percentage variance relation between species and environment factors |
41.83 | 63.71 | 79.61 | 91.78 |
| 所有典范轴的显著性测验 Test of significance of all canonical axes |
F=1.8 | P=0.002 | ||
| ** P<0.01; * P<0.05;CCA,即典范对应分析Canonical correspondence analysis | ||||
| 环境因子 Environmental factor |
重要性排序 Importance rank |
环境因子所占的解释量 Variance explains of environmental factors |
F | P |
| 海拔Altitude | 1 | 4.4 | 2.9 | 0.002 |
| 降水Precipitation | 2 | 3.7 | 2.5 | 0.002 |
| 温度Temperature | 3 | 1.8 | 1.2 | 0.242 |
| 坡向Aspect | 4 | 1.7 | 1.2 | 0.192 |
| 坡度Slope | 5 | 1.6 | 1.1 | 0.316 |
样方-环境因子CCA排序直观的呈现了高寒植被群落群丛类型分布与气候地形因子的关系(图5), CCA排序第一轴代表海拔环境梯度, 从左往右海拔逐渐升高。第二轴代表降水梯度, 从下往上降雨量逐渐增大。群丛Ⅱ青海云杉-匙叶小檗-甘肃薹草的分布与降水、温度、坡向的关系较为密切。群丛Ⅲ祁连圆柏-高山绣线菊-珠芽蓼受海拔和坡度的影响较大。群丛Ⅳ高山绣线菊-珠芽蓼+青藏蓼分布的海拔最高, 群丛Ⅰ红桦-红花蔷薇-甘肃薹草分布区降雨量较高。群丛Ⅴ沙棘+甘青蒿+鼠掌老鹳草与群丛Ⅵ沙棘+苦麦菜+苦荞麦主要分布在海拔较低的河流沟谷地带, 温度较高, 受坡度和坡向影响较小。

|
| 图 4 祁连山高寒植被群落样方与环境因子的CCA排序图 Fig. 4 CCA two-dimensional ordination diagram for samples and environmental variables of alpine communities in Qilian mountain CCA,即典范对应分析,Canonical Correspondence Analysis; S1—S65代表样品编号,S1—S65 represents sample number |
祁连山植被覆盖总体上自西向东递增, 呈现东多西少的分布格局[28]。植被类型主要包括温带落叶阔叶林、寒温性针叶林、温性灌丛、高寒灌丛、高原河谷灌丛、温性草原、高寒草原、荒漠、高寒草甸、沼泽草甸[29], 本研究调查区域获得的7个群丛主要对应森林、灌丛。祁连山东段高寒植被群落包括181个植物种, 隶属40科, 124属, 而北坡植物分属84科399属1 044种, 物种丰富度较东段高, 两个区域植物种的优势科都为菊科、禾本科、蔷薇科[30]。研究区乔木以青海云杉为主, 且占绝对优势, 在部分近阳坡的山地分布大量祁连圆柏, 在峡谷陡坡地带有桦木科的红桦和白桦, 河谷地段有斑块状山杨分布。乔木层植被组成稳定、个体生长缓慢, 而灌木层和草本层植被组成随乔木层的演替出现阶段性变化[31]。灌木是祁连山东段重要的植物类群, 从低山到高山都有灌木分布, 共有25种, 占植物种类的13.81%。近阳坡上分布有瑞香科的甘肃瑞香, 在部分阴坡或半阴坡上, 杜鹃花科的植物占优势, 形成高寒灌丛。群丛Ⅳ高山绣线菊+鬼箭锦鸡儿-珠芽蓼属于高寒落叶灌丛, 主要分布在山地阴坡及沟谷地带等。本研究调查发现金露梅是灌木层优势度最高的植物种, 也是祁连山最具代表性的高寒落叶灌丛[32], 在滩地或缓坡形成群落。
物种丰富度海拔梯度格局及其形成机制一直是宏观生态学研究的重要议题[33]。太白山木本物种组成取决于海拔主导的温度和湿度梯度[34]。神农架南坡植物多样性的垂直格局基本符合“单峰”模式, 峰值出现在海拔1400 —1500 m[35]。长白山北坡不同林层的乔、灌、草各生活型类群物种丰富度及多样性随海拔上升均表现出较明显的线性下降趋势[36]。随着海拔的升高, 滇西北玉龙雪山2900—4200 m间木本和草本的物种丰富度先增加后下降, 而灌木层的物种丰富度呈明显下降格局[37]。海拔是贺兰山灌丛群落的物种组成及其多样性垂直分布格局的主要限制因素[38]。祁连山区海拔2701—2800 m带是物种多样性的一个重要转折点, 是物种多样性较丰富的地带[39], 本研究也证明海拔是祁连山东段植被群落分布的最重要环境因子。由此可见, 不同地理气候带海拔对群落尺度山地植被分布具有决定性作用。
在区域至全球尺度上, 水热条件是决定植物种、生活型或植被类型分布的主导因素, 而在景观及更小的尺度上, 区域环境因子则主导着植被的分布格局[40]。区域尺度上, 降水和温度是影响祁连山东段植被种类和数量的主要因素, 也是影响植被NDVI变化的主要因子[41], 植被空间分布与降水空间分布一致[42]。以400 mm年降水线为界, 在阴坡、阳坡分别形成不同的演替格局[43]。景观群落尺度, 地形是造成土壤及植物分布格局分异的重要驱动因子[44]。生境的热量、水分(养分)和光照条件等小气候条件, 土壤厚度、养分空间差异可主要由海拔、坡位和坡度、坡向等地形特征分别反映[45]。随海拔的升高, 年均温度降低, 降雨量增大[16], 土壤pH和盐分减小, 有机质、土壤水分、全氮增大[46]。南坡、上坡位、坡度缓的山地生境土壤肥力条件较好, 海拔是影响土壤有机质的重要因子[47]。从南坡到北坡土壤水分增大, 温度和pH降低, 有机碳增大, 但容重减小。草本主要分布在南坡、西南坡、西北坡, 北坡主要生长乔木[48]。由海拔、坡向主导的降水量、温度等小气候和土壤质地的综合作用是山地植被分布的决定性因素[49]。与上述结论一致, 本研究证明海拔、降水、温度共同影响着祁连山高寒植被群落的物种分布。
4 结论(1) 祁连山东段高寒植被群落包括181个植物种, 隶属40科, 124属。群落层片由乔木层、灌木层和草本层组成, 乔木8种, 灌木25种, 草本148种。乔木层优势种有青海云杉、祁连圆柏、红桦。灌木层优势种有金露梅、山生柳、匙叶小檗、高山绣线菊。草本层优势种有甘肃薹草、珠芽蓼、早熟禾、唐松草、甘青蒿。
(2) 祁连山东段高寒植被群落主要包括7个群丛类型:群丛Ⅰ红桦-红花蔷薇-甘肃薹草, 群丛Ⅱ青海云杉-匙叶小檗-甘肃薹草, 群丛Ⅲ祁连圆柏-高山绣线菊-珠芽蓼, 群丛Ⅳ高山绣线菊+鬼箭锦鸡儿-珠芽蓼, 群丛Ⅴ沙棘+甘青蒿+鼠掌老鹳草, 群丛Ⅵ沙棘+苦荬菜+苦荞麦, 群丛Ⅶ沙棘-冰草+西北沼委陵菜。
(3) 海拔是祁连山东段高寒植被群落植物种分布的最重要环境因子, 其次是降水、温度、坡向、坡度。
| [1] |
方精云, 沈泽昊, 崔海亭. 试论山地的生态特征及山地生态学的研究内容. 生物多样性, 2004, 12(1): 10-19. DOI:10.3321/j.issn:1005-0094.2004.01.003 |
| [2] |
应俊生. 中国种子植物物种多样性及其分布格局. 生物多样性, 2001, 9(4): 393-398. DOI:10.3321/j.issn:1005-0094.2001.04.011 |
| [3] |
沈泽昊, 张新时, 金义兴. 地形对亚热带山地景观尺度植被格局影响的梯度分析. 植物生态学报, 2000, 24(4): 430-435. DOI:10.3321/j.issn:1005-264X.2000.04.008 |
| [4] |
Pinder Ⅲ J E, Kroh G C, White J D, Basham May A M. The relationships between vegetation type and topography in lassen volcanic national park. Plant Ecology, 1997, 131(1): 17-29. DOI:10.1023/A:1009792123571 |
| [5] |
胡志伟, 沈泽昊, 吕楠, 赵俊, 李道兴, 陈华, 王功芳. 地形对森林群落年龄及其空间格局的影响. 植物生态学报, 2007, 31(5): 814-824. |
| [6] |
李英年, 沈振西, 周华坤. 寒冻雏形土不同地形部位土壤湿度及其与主要植被类型的对应关系. 山地学报, 2001, 19(3): 220-225. DOI:10.3969/j.issn.1008-2786.2001.03.005 |
| [7] |
熊斌梅, 雷耘, 汪正祥, 李亭亭, 熊秀海, 李中强, 张娥, 彭宗林. 地形对七姊妹山自然保护区植物丰富度及分布格局的影响. 西北植物学报, 2016, 36(11): 2307-2313. DOI:10.7606/j.issn.1000-4025.2016.11.2307 |
| [8] |
钟娇娇, 陈杰, 陈倩, 姬柳婷, 康冰. 秦岭山地天然次生林群落MRT数量分类、CCA排序及多样性垂直格局. 生态学报, 2019, 39(1): 277-285. |
| [9] |
刘杰, 罗亚皇, 李德铢, 高连明. 青藏高原及毗邻区植物多样性演化与维持机制:进展及展望. 生物多样性, 2017, 25(2): 163-174. |
| [10] |
张树斌, 王襄平, 吴鹏, 孙晗, 李巧燕, 吴玉莲, 韩威, 武娴. 吉林灌木群落物种多样性与气候和局域环境因子的关系. 生态学报, 2018, 38(22): 7990-8000. |
| [11] |
王亚锋, 刘波. 气候和地形对青藏高原树线位置的影响. 生态学杂志, 2018, 37(12): 3508-3515. |
| [12] |
Tian Q Y, He Z B, Xiao S C, Peng X M, Ding A J, Lin P F. Response of stem radial growth of Qinghai spruce (Picea crassifolia) to environmental factors in the Qilian Mountains of China. Dendrochronologia, 2017, 44: 76-83. DOI:10.1016/j.dendro.2017.04.001 |
| [13] |
吴安驰, 邓湘雯, 任小丽, 项文化, 张黎, 葛蓉, 牛忠恩, 何洪林, 何立杰. 中国典型森林生态系统乔木层群落物种多样性的空间分布格局及其影响因素. 生态学报, 2018, 38(21): 7727-7738. |
| [14] |
任正超, 朱华忠, 史华, 柳小妮. 中国潜在自然植被NPP时空分布格局变化及其对气候和地形的响应. 草地学报, 2017, 25(3): 474-485. |
| [15] |
李卓, 张继超, 孙然好, 张翀. 气候和地形对植被覆盖动态演化的影响研究. 生态科学, 2017, 36(6): 35-43. |
| [16] |
Yang W J, Wang Y H, Webb A A, Li Z Y, Tian X, Han Z T, Wang S L, Yu P T. Influence of climatic and geographic factors on the spatial distribution of Qinghai spruce forests in the dryland Qilian Mountains of Northwest China. Science of the Total Environment, 2018, 612: 1007-1017. DOI:10.1016/j.scitotenv.2017.08.180 |
| [17] |
王根绪, 刘国华, 沈泽昊, 王文志. 山地景观生态学研究进展. 生态学报, 2017, 37(12): 3967-3981. |
| [18] |
柳小妮, 孙九林, 张德罡, 蒲小鹏, 徐广平. 东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究. 草业学报, 2008, 17(4): 1-11. DOI:10.3321/j.issn:1004-5759.2008.04.001 |
| [19] |
雷蕾, 刘贤德, 王顺利, 李毅, 张学龙. 祁连山高山灌丛生物量分配规律及其与环境因子的关系. 生态环境学报, 2011, 20(11): 1602-1607. DOI:10.3969/j.issn.1674-5906.2011.11.003 |
| [20] |
孟好军, 刘建泉, 刘贤德, 赵维俊. 祁连山青海云杉群落物种组成及α多样性垂直分布. 生态环境学报, 2011, 28(3): 435-440. DOI:10.3969/j.issn.1674-5906.2011.03.008 |
| [21] |
赵维俊, 刘贤德, 金铭, 车宗玺, 敬文茂, 马钰. 祁连山青海云杉林群落结构特征分析. 干旱区研究, 2012, 29(4): 615-620. |
| [22] |
王国宏. 祁连山北坡中段植物群落多样性的垂直分布格局. 生物多样性, 2002, 10(1): 7-14. DOI:10.3321/j.issn:1005-0094.2002.01.002 |
| [23] |
贾文雄, 陈京华, 张禹舜, 曹文侠. 祁连山北坡草地植物群落特征与土壤水热因子的关系. 生态学杂志, 2016, 35(3): 661-667. |
| [24] |
汪有奎, 杨全生, 郭生祥, 李进军, 王零, 袁虹. 祁连山北坡森林资源变迁. 干旱区地理, 2014, 37(5): 966-979. |
| [25] |
刘贤德, 张学龙, 赵维俊, 车宗玺, 杨逍虎, 苗毓鑫, 张克海. 祁连山西水林区亚高山灌丛水文功能的综合评价. 干旱区地理, 2016, 39(1): 86-94. |
| [26] |
葛丽娟, 党虹, 戎战磊, 高云飞, 赵传燕, 王红义. 祁连山中段植被群落的时空格局变化. 干旱区研究, 2018, 35(2): 346-353. |
| [27] |
贾文雄, 赵珍, 俎佳星, 陈京华, 王洁, 丁丹. 祁连山不同植被类型的物候变化及其对气候的响应. 生态学报, 2016, 36(23): 7826-7840. |
| [28] |
武正丽, 贾文雄, 刘亚荣, 张禹舜. 近10a来祁连山植被覆盖变化研究. 干旱区研究, 2014, 31(1): 80-87. |
| [29] |
陈桂琛, 彭敏, 黄荣福, 卢学峰. 祁连山地区植被特征及其分布规律. 植物学报, 1994, 36(1): 63-72. DOI:10.3321/j.issn:1000-4025.1994.01.012 |
| [30] |
易玉媛, 王红义. 祁连山北坡植被群落垂直分布特征. 防护林科技, 2013(10): 6-9, 16-16. DOI:10.3969/j.issn.1005-5215.2013.10.003 |
| [31] |
何芳兰, 徐先英, 尉秋实, 刘虎俊, 贺访印, 马俊梅, 金红喜. 祁连山青海云杉人工林与天然林群落结构特征及物种多样性比较研究. 西北林学院学报, 2016, 31(5): 1-7. DOI:10.3969/j.issn.1001-7461.2016.05.01 |
| [32] |
乔雨, 赵传燕, 戎占磊, 解欢欢, 高婵婵, 高云飞, 王方圆. 祁连山中部金露梅种群生长过程中的植被光谱特征. 生态学杂志, 2017, 36(1): 11-20. |
| [33] |
刘开明, 郑智, 龚大洁. 物种丰富度的垂直分布格局及其形成机制. 生态学杂志, 2017, 36(2): 541-554. |
| [34] |
唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释. 生物多样性, 2004, 12(1): 115-122. DOI:10.3321/j.issn:1005-0094.2004.01.014 |
| [35] |
沈泽昊, 胡会峰, 周宇, 方精云. 神农架南坡植物群落多样性的海拔梯度格局. 生物多样性, 2004, 12(1): 99-107. DOI:10.3321/j.issn:1005-0094.2004.01.012 |
| [36] |
郝占庆, 于德永, 杨晓明, 丁之慧. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 2002, 13(7): 785-789. DOI:10.3321/j.issn:1001-9332.2002.07.005 |
| [37] |
冯建孟, 王襄平, 徐成东, 杨元合, 方精云. 玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局. 山地学报, 2006, 24(1): 110-116. DOI:10.3969/j.issn.1008-2786.2006.01.016 |
| [38] |
苏闯, 张芯毓, 马文红, 赵利清, 梁存柱. 贺兰山灌丛群落物种多样性海拔格局及环境解释. 山地学报, 2018, 36(5): 699-708. |
| [39] |
常学向, 赵文智, 赵爱芬. 祁连山区不同海拔草地群落的物种多样性. 应用生态学报, 2004, 15(9): 1599-1603. DOI:10.3321/j.issn:1001-9332.2004.09.022 |
| [40] |
王敏, 周才平. 山地植物群落数量分类和排序研究进展. 南京林业大学学报:自然科学版, 2011, 35(4): 126-130. |
| [41] |
陈京华, 贾文雄, 赵珍, 张禹舜, 刘亚荣. 1982-2006年祁连山植被覆盖的时空变化特征研究. 地球科学进展, 2015, 30(7): 834-845. |
| [42] |
蒋友严, 杜文涛, 黄进, 赵慧珍, 王成福. 2000-2015年祁连山植被变化分析. 冰川冻土, 2017, 39(5): 1130-1136. |
| [43] |
张永虎. 祁连山东段植被分布特征及演替规律. 人民黄河, 2010, 32(8): 94-95. DOI:10.3969/j.issn.1000-1379.2010.08.041 |
| [44] |
牛钰杰, 周建伟, 杨思维, 王贵珍, 刘丽, 花立民. 坡向和海拔对高寒草甸山体土壤水热和植物分布格局的定量分解. 应用生态学报, 2017, 28(5): 1489-1497. |
| [45] |
McDonald D J, Cowling R M, Boucher C. Vegetation-environment relationships on a species-rich coastal mountain range in the fynbos biome (South Africa). Vegetatio, 1996, 123(2): 165-182. DOI:10.1007/BF00118269 |
| [46] |
Zhang H Y, Qian Y B, Wu Z N, Wang Z C. Vegetation-environment relationships between northern slope of Karlik mountain and Naomaohu Basin, East Tianshan Mountains. Chinese Geographical Science, 2012, 22(3): 288-301. DOI:10.1007/s11769-012-0536-y |
| [47] |
Fu B J, Liu S L, Ma K M, Zhu Y G. Relationships between soil characteristics, topography and plant diversity in a heterogeneous deciduous broad-leaved forest near Beijing, China. Plant and Soil, 2004, 261(1/2): 47-54. DOI:10.1023/B:PLSO.0000035567.97093.48 |
| [48] |
Qin Y Y, Nicholas H, Feng Q, Zhu M. Influence of slope aspect on plant community composition and its implications for restoration of a Chinese Mountain Range. Polish Journal of Environmental Studies, 2017, 26(1): 375-383. DOI:10.15244/pjoes/64458 |
| [49] |
Burke A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. Journal of Vegetation Science, 2001, 12(1): 53-60. DOI:10.1111/j.1654-1103.2001.tb02616.x |