 2020, Vol. 40
2020, Vol. 40文章信息
- 王延芳, 张永香, 勾晓华, 高琳琳, 王放
- WANG Yanfang, ZHANG Yongxiang, GOU Xiaohua, GAO Linlin, WANG Fang
- 祁连山中部低海拔地区青海云杉径向生长的气候响应机制
- Climate response mechanism of radial growth of Picea crassifolia in low altitude area of middle Qilian Mountains
- 生态学报. 2020, 40(1): 161-169
- Acta Ecologica Sinica. 2020, 40(1): 161-169
- http://dx.doi.org/10.5846/stxb201811022366
-
文章历史
- 收稿日期: 2018-11-02
- 网络出版日期: 2019-10-25





2. 中国气象局国家气候中心, 北京 100081
2. National Climate Center, China Meteorological Administration, Beijing 100081, China
森林是全球重要的生态系统, 对改善区域气候, 防止水土流失、维持生态系统多样性等有重要作用[1-2]。目前, 以变暖为主要特征的全球气候变化正在加剧, 森林对此响应强烈, 并引发了一系列的生态问题[3-5]。其中, 树木径向生长对气候变化的响应为该领域研究的热点问题之一。气候变化使得树木生长的物候改变, 生长季延长或缩短, 光合峰值时间变化, 树木基面积生长量改变等[6-8];此外, 树木对气候要素的敏感性也会随之改变[9-10]。对不同地区, 不同树种的径向生长而言, 气候变化对其的影响差异较大, 作用机制也不尽相同。因此, 开展特定地区、特定树种径向生长对气候要素的响应机制研究将有助于更好地掌握树木生长对气候的响应机理, 更好地理解和预估气候变化情景下森林生态系统的变化。
祁连山位于青藏高原东北缘, 是我国典型的寒冷干旱地区, 具有典型的大陆性干旱半干旱气候。同时, 该区域也是季风和西风系统的交汇作用区, 区域气候变化对全球气候变化响应敏感[11-12]。青海云杉是祁连山森林的主要建群树种, 也是干旱半干旱区典型的针叶树种, 具有抗旱、耐寒、对气候敏感等特性[13]。因此, 研究祁连山中部青海云杉径向生长的气候响应机制有重要意义。
目前, 针对青海云杉径向生长-气候要素响应关系的研究大部分是在近线性关系的基础上对树轮序列和器测数据进行统计分析[14-16], 缺乏生理机制的解释。Vaganov-Shashkin (VS)模型综合考虑了外界环境与形成层细胞生长之间的关系, 通过各类参数, 模拟外界环境对树木径向生长的影响[17-22]。因其参数相对容易获得, 有涉及细胞尺度的模拟, VS模型在国内外被广泛应用, 并取得较好的模拟效果[17-18, 20-26]。本文以祁连山地区青海云杉为研究对象, 利用VS模型, 分析祁连山青海云杉径向生长对气候要素的响应, 探究祁连山中部青海云杉径向生长的气候响应机制, 以期为研究气候变化情景下森林生态系统的变化奠定基础, 为未来祁连山自然保护区森林管理提供理论依据。
1 资料与方法 1.1 树轮资料青海云杉林是祁连山区主要森林类型, 其面积约占乔木林总面积的四分之三, 海拔跨度自2500—3300 m[27-28]。本研究所利用的青海云杉样本采于张掖市肃南裕固族自治县康乐乡大草滩村, 样点代号为P1(东经99°55′ 59″, 北纬38°49′ 2″, 海拔2660m, 图 1)。采样点海拔较低, 坡度适中, 整体比较干旱, 草本植物稀疏, 生境条件相对较差。将采集的树木年轮样芯带回实验室, 对样芯进行固定、风干、打磨、交叉定年、轮宽测量、COFECHA程序校正等处理。共计55根样芯, 26棵树, 无样芯被剔除。利用ARSTAN程序建立年表, 主要采用负指数函数和线性函数方法进行年轮序列趋势拟合, 个别采用样条函数拟合, 最终得出去趋势后的树轮宽度指数年表。

|
| 图 1 采样点分布图 Fig. 1 Locations of sampling sites |
一般选用样本间相关系数R1、树间相关系数R2、同一棵树不同树芯间相关系数R3、信噪比SNR、样本解释信号强度EPS、年表第一主成分解释方差PC1等参数来评估年表的质量[29-32]。EPS为共同信号与年表总变异(共同信号+噪音)的比值, EPS值越大, 共同信号越强。气候信号与非气候因素形成的噪音的比值即为信噪比, SNR越大, 年表包含的气候信息越多。PC1越大, 说明年表中各样本序列的同步性越大, 气候因子对树木年轮宽度生长的限制作用越强, 年表中的气候信息含量就越多。鉴于差值年表(Residual Chronology, 简称RES)各项参数高于标准年表(Standard Chronology, 简称STD), 且考虑到树木种内竞争引起的低频干扰, 因此研究分析选用差值年表。
| 采样点Sampling point | P1 | |
| 年表类型Chronologies | 标准年表 | 差值年表 |
| 可靠时段Common period | 1859—2016 | 1859—2016 |
| 样本间相关系数Mean correlations among all radii | 0.631 | 0.699 |
| 树间相关系数Mean correlations between trees | 0.668 | 0.735 |
| 同一棵树不同树芯间相关系数Mean correlations within trees | 0.627 | 0.695 |
| 信噪比Signal-to-noise ratio | 78.755 | 106.786 |
| 样本解释信号强度Expressed population signal | 0.987 | 0.991 |
| 年表第一主成分解释方差Variance in first eigenvector | 65.1% | 71.0% |
| P1:采样点代号 | ||
采样点距离张掖气象站(100°15′ 36″E, 38°33′ 36″N)较近, 直线距离是34.91km, 附近无其他距离较近的气象站, 故选用张掖气象站的气温和降水量数据。张掖站器测资料显示, 研究区雨热同期, 蒸发量高于降水量, 日温差和年温差均较大, 年内最高温出现在6—8月, 降水集中在5—9月, 降水量占全年80%以上, 尤其是7—8月, 降水量接近全年的50%(图 2)。统计表明, 月最高气温为32.9℃, 出现在7月;月最低气温为-21.1℃, 出现在1月;月均最大降水量为90.3mm, 出现在8月。自1951年以来, 年均温显著增加(R=0.81, P<0.01), 年降水量略有增加趋势, 相对湿度显著降低(R=0.46, P<0.01), 研究区气候趋于干热。野牛沟站、祁连站、山丹站、张掖站、高台站器测数据的计算结果表明研究区气温随海拔升高显著递减, 垂直递减率为5.4℃/km (R=0.99, P<0.01)。数据分析时, 根据张掖站和采样点的海拔差, 将温度插值到采样点海拔处的温度, 降水量采用张掖气象站原始器测数据。干旱数据选用的是距离采样点较近的格点数据帕尔默干旱指数(Palmer drought severity index, PDSI), 分辨率0.25°×0.25°。时间尺度是1953年到2016年。

|
| 图 2 张掖气象站多年月平均降水和月均温分布图 Fig. 2 Distribution of monthly mean precipitation and monthly mean temperature over Zhangye meteorological station |
在VS模型中, 影响树木生长的因子主要有3个:降水、温度和光照。VS模型就是以这3个参数为主要输入条件进行树木生长模拟, 还考虑到土壤层厚度、采样点坡度、植物主要根系深度以及土壤最大含水率等因素的影响[17-18, 20-26]。
树木的生长速率由上面提到的3个最主要的因子决定, 即:
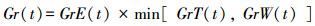
|
(1) |
式中, Gr(t)为树木的日生长速率, GrE(t)、GrT(t)和GrW(t)为假定其他因子不影响生长时, 分别受光照、温度和土壤湿度限制时得到的生长速率。
温度对树木生长的影响遵循温度三基点原则。气温达到Tbeg时, 树木开始生长;气温低于Tmin, 树木不生长;气温大于Tmin而低于Topt1时, 树木生长随着气温的升高以线性速度增加;气温在Topt1和Topt2之间时, 生长速度维持在最高水平;气温高于Topt2而小于Tmax时, 生长速度随着气温升高以线性速度下降(Tbeg代表生长积温;Tmin代表最低生长温度;Topt1代表最适生长温度的下限;Topt2代表最适生长温度上限;Tmax代表最高生长温度)。
土壤湿度对树木生长的影响类似于温度。包含了Wmin、Wopt1、Wopt2和Wmax等4个参数, 含义同上。每日土壤含水量的变化(dW)由土壤水动态平衡方程计算得到:
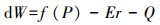
|
(2) |
式中, f (P)为日降水量, Er是日蒸腾量, Q为土壤中水的日径流量。其中,
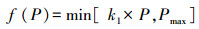
|
(3) |
式中, P表示实际的日降水量, k1为渗透系数, Pmax为土壤饱和的最大日降水量。
光照影响的树木生长速率GrE(t) (忽略地球绕太阳运行时的离心率以及大气的透射率), 可用纬度φ、太阳入射角(θ)和日长(φ)表示:
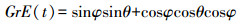
|
(4) |
模拟青海云杉径向生长情况, 首先要选择合适参数。因此本文先参照已有模拟研究确定了部分参数的范围[18, 20, 22, 26], 如Zhang等[26]结合观测和模拟数据研究了祁连山中东西部祁连圆柏的生理过程, Yang等[20]基于观测数据、遥感数据及VS模型对整个青藏高原的针叶树种进行了生理过程研究。其中, Zhang等文章中的祁连县采样点、Yang等文章中的寺大隆采样点与本文中采样点距离较近, 且树木生境较为相似。因此Zhang等文章中的使土壤饱和的最大日降水量Pmax, Yang等文章中的树木开始生长的积温Tbeg, 根深Droot, 降水渗透到土壤的系数k1, 植物蒸腾量k2、k3被用于本文的模拟。其他少数参数由VS模型给出的参数范围模拟调整获得。
另外, 本研究利用软件Matlab R2014b进行数据模拟, SPSS 16.0进行Pearson相关分析, Excel进行标准差分析, 采用ArcMap 10.3和Origin 8.6做图。
2 结果 2.1 青海云杉轮宽-气候要素响应分析青海云杉树轮宽度年表与上一年9月和当年5月的降水量存在显著正相关关系, 与上年9月到当年6月的降雨累积量也存在显著正相关关系, 与上一年8月、12月和当年6月、7月、8月的降水量存在正相关关系但未达到显著水平(图 3)。青海云杉树轮宽度年表与月均温无显著相关关系。青海云杉树轮宽度年表与上一年9—12月和当年1—7月的PDSI均存在显著正相关关系, 与5月、6月PDSI的相关系数最大。

|
| 图 3 青海云杉轮宽指数与月均温、月降水量和PDSI的的响应关系 Fig. 3 Correlations between chronology and temperature、precipitation、PDSI 横坐标中, P代表上年, C代表当年, PxCy代表上年x月到当年y月的累计量;0.01代表相关系数达到了99%的置信水平, 0.05代表相关系数达到了95%的置信水平 |
在确定了部分参数的基础上, 对Tmin、Topt1、Topt2、Tmax、Wmin、Wopt1、Wopt2、Wmax进行调试, 模拟参数如表 2所示。
| 参数Parameter | 描述(单位) Description (units) | 取值Value |
| Tmin | 最低生长温度(℃) | 4.5 |
| Topt1 | 最适生长温度下限(℃) | 12.8 |
| Topt2 | 最适生长温度上限(℃) | 20.0 |
| Tmax | 最高生长温度(℃) | 31.0 |
| Wmin | 生长的最低土壤湿度(v/v) | 0.034 |
| Wopt1 | 生长的最适土壤湿度下限(v/v) | 0.055 |
| Wopt2 | 生长的最适土壤湿度上限(v/v) | 0.80 |
| Wmax | 生长的最高土壤湿度(v/v) | 0.90 |
| Tbeg | 开始生长的积温(℃) | 100 |
| Droot | 根深(mm) | 750 |
| Pmax | 使土壤饱和的最大日降水量(mm) | 20 |
| k1 | 降水渗透到土壤的系数 | 0.60 |
| k2 | 植物蒸腾量的第一系数, 和降水有关(mm/day) | 0.035 |
| k3 | 植物蒸腾量的第二系数, 和温度有关(1/℃) | 0.18 |
1953—2016年, 模拟轮宽与实测轮宽的相关系数为0.565(P<0.01);1953—2003年模拟轮宽与实测轮宽的相关系数为0.668(P<0.01), 2004—2016年模拟轮宽与实测轮宽相关系数较小, 且不存在显著相关关系(图 4)。1953—2003年模拟轮宽与实测值拟合较好, 而2004—2016年模拟轮宽与实测值无显著相关, 可能是21世纪以来, 人为活动对树木生长影响较大导致的。因此, 选用1953—2003年的模拟结果进行生理机制分析。

|
| 图 4 实测和模拟的径向生长序列对比 Fig. 4 Comparison of observed and simulated radial growth sequences |
我们将差值年表平均值加差值年表的标准差定义为宽轮界限, 将差值年表平均值减差值年表的标准差定义为窄轮界限[33]。利用EXCEL软件对差值年表进行标准差分析以及宽轮界限和窄轮界限的计算, 得到特征窄年共9年, 分别是1957、1962、1966、1968、1981、1985、1995、2001、2003年;得到特征宽年共9年, 分别为1955、1956、1964、1972、1979、1980、1983、1998、2002年。
为更好地理解气候要素如何影响青海云杉树轮宽度, 本文利用VS模型来研究树木形成宽轮和窄轮时的年内生长差异。如图 5所示, 在宽轮和窄轮的模拟对比中, 温度影响的青海云杉生长速率无显著差异;土壤湿度影响的树木生长速率存在不同, 主要表现为, 形成宽轮时, 青海云杉5—8月份生长速率较高, 且5月下旬至6月份土壤湿度影响的树木生长速率最高。

|
| 图 5 特征宽(黑实线)窄(红实线)年土壤湿度和温度决定的树木径向生长速率 Fig. 5 Simulated mean growth rate depending on soil moisture and temperature in pointer years of wide rings (black line) and narrow rings (red line) |
在我国干旱半干旱区, VS模型已被广泛用于针叶树种的研究, 研究对象多为祁连圆柏[20, 22, 26]和油松[24-25], 并取得了较好的研究成果。本研究利用VS模型对祁连山中部低海拔地区青海云杉进行了生理机制模拟, 模拟轮宽与实测轮宽拟合较好, 表明VS模型适用于干旱半干旱区青海云杉的研究。
3.1 生长季降水对青海云杉径向生长的影响水分作为至关重要的非生物因素, 能限制植物的生长、生存和繁殖[34-37]。对1953—2003年的模拟结果进行分析, 形成宽轮和窄轮时, 土壤湿度影响的树木生长速率存在不同, 形成宽轮时, 5—8月份生长速率较高。土壤湿度分布特征与降水数据相吻和[38]。通过分析该区域的月降水量发现, 径向生长宽年5—8月降水累积量平均值为115.8mm, 最高年份可达177.5mm, 除了1955年、1980年低于100mm以外, 其他年份均高于100mm。径向生长窄年, 5—8月降水累积量平均值为60.5mm。形成宽轮年份5—8月降水累积量是形成窄轮的年份5—8月降水累积量的1.9倍。故5—8月降水量对祁连山中部低海拔地区青海云杉径向生长起着决定性作用。
张永香等[22]对柴达木东北缘的祁连圆柏进行了模拟研究, 结果表明, 降水(尤其是5、6月份)决定了祁连圆柏的径向生长。Zhang等[26]通过模拟与木材解剖的方法, 研究了祁连山不同地区祁连圆柏的生长, 结果表明, 5、6月份是祁连圆柏径向生长的主要时期, 降水决定了祁连圆柏的径向生长。Shi等[24]利用统计方法和VS模型研究了贺兰山油松的生理机制, 结果显示5—8月份的水热条件对树木生长影响最大。史江峰等[25]通过VS模型研究了贺兰山油松的生长情况, 结果表明, 4—8月的降水量决定了该年的树轮宽度。对比分析表明, 祁连山中部低海拔地区青海云杉的径向生长主要受水分条件限制, 与祁连圆柏、油松径向生长的限制因子较为一致。
生长季的土壤水分条件好, 温度适宜, 会提高树木的光合速率, 促进细胞的分裂和扩大, 树木径向生长量增大[39-45]。生长季的土壤水分条件差, 会影响树木的光合作用[46-47]。水分对植物光合作用的影响主要通过两个途径实现, 气孔抑制和非气孔抑制。气孔抑制是指土壤水分少时, 水分胁迫使气孔导度下降, 二氧化碳进入叶片受阻而使光合下降;非气孔抑制是指由木质部运输到叶片的水分减少, 影响光合器官的活性和植物激素的分泌[48-49]。因此, 水分胁迫会使树木内部的正常生理功能受到干扰, 光合作用降低, 光合作用形成的产物少, 形成层获得的能量少, 不利于形成层细胞的分化、分裂、拉伸、成熟, 因此树木径向生长量小[50-53]。
3.2 降水量对青海云杉径向生长的滞后影响进一步研究发现, 虽然1955年、1980年5—8月降水累积量低于100mm, 但是1954年、1979年9月份降水量明显高于其他年份。表明径向生长除了受当年生长季降水量的影响外, 上年9月份的降水量也会影响当年青海云杉的径向生长。在相关分析中, 树轮宽度指数年表与上年9月份降水量存在显著正相关关系。研究结果表明, 降水对青海云杉径向生长的影响存在滞后效应。
在西北干旱半干旱区, 降水对树木径向生长的影响在众多的气象因子中表现尤为突出。多项研究表明[15, 33, 39, 42-44, 54], 不仅生长季降水量与树轮宽度指数存在显著正相关关系, 上一年9月份降水量与树轮宽度指数也存在显著正相关关系, 表现出明显的滞后效应。阿尔泰山西伯利亚落叶松树轮稳定碳同位素表现出较强的一阶自相关, 当年对翌年有将近40%的气候滞后和同位素滞后影响, 表明当年的生长季末期的降水会对第二年树木生长产生影响[55]。
树木光合作用结束时间晚于形成层活动结束时间, 形成层活动结束后, 光合作用产生的能量一部分用于木材细胞壁加厚, 一部分能量会被储存[39]。在温带低海拔地区, 松树的径向生长早于叶片展开前, 说明树木径向生长开始时所需的能量来自前一年储存的能量[56]。在祁连山中部低海拔地区青海云杉的生长过程中, 当年9月份的降水量可能会影响光合产物的产生、储存以及再分配, 由于当年储存的能量会用于第二年青海云杉的径向生长, 因此, 当年9月份的降水量会对第二年青海云杉的径向生长产生滞后影响。
4 结论VS模型能很好的用于干旱半干旱区青海云杉生理机制的模拟, 可以从生理机制的角度解释树木-气候要素响应关系。模拟结果表明, 降水量对青海云杉径向生长起着决定性作用, 5—8月份的降水(尤其是5、6月份)直接影响青海云杉的径向生长。9月份的降水量也会影响翌年青海云杉的径向生长。温度对青海云杉影响较小, 可能是采样点位于低海拔地区, 温度适宜, 没有成为限制因素。在低海拔地区的青海云杉人工林建设时, 可适时浇灌, 促进树木生长, 加速人工林建设。
致谢: 感谢马里兰大学Evans, M副教授对VS模型的帮助。| [1] |
汪有奎, 郭生祥, 汪杰, 袁虹, 徐柏林, 王多尧. 甘肃祁连山国家级自然保护区森林生态系统服务价值评估. 中国沙漠, 2013, 33(6): 1905-1911. |
| [2] |
Zhang Y, Drobyshev I, Gao L S, Zhao X H, Bergeron Y. Disturbance and regeneration dynamics of a mixed Korean pine dominated forest on Changbai Mountain, North-Eastern China. Dendrochronologia, 2014, 32(1): 21-31. DOI:10.1016/j.dendro.2013.06.003 |
| [3] |
Allen C D, Macalady A K, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears D D, Hogg E H, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J H, Allard G, Running S W, Semerci A, Cobb N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 2010, 259(4): 660-684. DOI:10.1016/j.foreco.2009.09.001 |
| [4] |
邬建国, 安树青, 冷欣. 现代生态学讲座(Ⅵ):全球气候变化与生态格局和过程. 北京: 高等教育出版社, 2013.
|
| [5] |
张颖, 杨桂红. 森林碳汇与气候变化. 北京: 中国林业出版社, 2015.
|
| [6] |
Qi Z H, Liu H Y, Wu X C, Hao Q. Climate-driven speedup of alpine treeline forest growth in the Tianshan Mountains, Northwestern China. Global Change Biology, 2015, 21(2): 816-826. DOI:10.1111/gcb.12703 |
| [7] |
Wu X C, Liu H Y. Consistent shifts in spring vegetation green-up date across temperate biomes in China, 1982-2006. Global Change Biology, 2013, 19(3): 870-880. DOI:10.1111/gcb.12086 |
| [8] |
Xu C Y, Liu H Y, Williams A P, Yin Y, Wu X C. Trends toward an earlier peak of the growing season in Northern Hemisphere mid-latitudes. Global Change Biology, 2016, 22(8): 2852-2860. DOI:10.1111/gcb.13224 |
| [9] |
Liang E Y, Shao X M, Eckstein D, Liu X H. Spatial variability of tree growth along a latitudinal transect in the Qilian Mountains, northeastern Tibetan Plateau. Canadian Journal of Forest Research, 2010, 40(2): 200-211. DOI:10.1139/X09-186 |
| [10] |
Liu H Y, Yin Y, Hao Q, Liu G. Sensitivity of temperate vegetation to Holocene development of East Asian monsoon. Quaternary Science Reviews, 2014, 98: 126-134. DOI:10.1016/j.quascirev.2014.06.006 |
| [11] |
Chen F H, Yu Z C, Yang M L, Ito E, Wang S M, Madsen D B, Huang X Z, Zhao Y, Sato T, Birks H J B, Boomer I, Chen J H, An C B, Wünnemann B. Holocene moisture evolution in arid central Asia and its out-of-phase relationship with Asian monsoon history. Quaternary Science Reviews, 2008, 27(3/4): 351-364. |
| [12] |
Fang K Y, Gou X H, Chen F H, Li J B, D'Arrigo R, Cook E, Yang T, Davi N. Reconstructed droughts for the southeastern Tibetan Plateau over the past 568 years and its linkages to the Pacific and Atlantic Ocean climate variability. Climate Dynamics, 2010, 35(4): 577-585. DOI:10.1007/s00382-009-0636-2 |
| [13] |
何芳兰, 徐先英, 尉秋实, 刘虎俊, 贺访印, 马俊梅, 金红喜. 祁连山青海云杉人工林与天然林群落结构特征及物种多样性比较研究. 西北林学院学报, 2016, 31(5): 1-7. DOI:10.3969/j.issn.1001-7461.2016.05.01 |
| [14] |
Gou X, Chen F, Yang M, Li J, Peng J, Jin L. Climatic response of thick leaf spruce (Picea crassifolia) tree-ring width at different elevations over Qilian Mountains, northwestern China. Journal of Arid Environments, 2005, 61(4): 513-524. DOI:10.1016/j.jaridenv.2004.09.011 |
| [15] |
彭剑峰, 勾晓华, 陈发虎, 刘普幸, 张永, 方克艳. 阿尼玛卿山地不同海拔青海云杉(Picea crassifolia)树轮生长特性及其对气候的响应. 生态学报, 2007, 27(8): 3268-3276. DOI:10.3321/j.issn:1000-0933.2007.08.021 |
| [16] |
徐金梅, 鲍甫成, 吕建雄, 黄荣风, 赵有科, Robea E. 祁连山青海云杉径向生长对气候的响应. 北京林业大学学报, 2012, 34(2): 1-6. DOI:10.3969/j.issn.1671-6116.2012.02.001 |
| [17] |
Anchukaitis K J, Evans M N, Kaplan A, Vaganov E A, Hughes M K, Grissino-Mayer H D, Cane M A. Forward modeling of regional scale tree-ring patterns in the southeastern United States and the recent influence of summer drought. Geophysical Research Letters, 2006, 33(4): L04705. |
| [18] |
Vaganov E A, Anchukaitis K J, Evans M N. How well understood are the processes that create dendroclimatic records? A mechanistic model of the climatic control on conifer tree-ring growth dynamics//Hughes M K, Swetnam T W, Diaz H F, eds. Dendroclimatology: Progress and Prospects. Dordrecht: Springer, 2011.
|
| [19] |
Vaganov E A, Hughes M K, Kirdyanov A V, Schweingruber F H, Silkin P P. Influence of snowfall and melt timing on tree growth in subarctic Eurasia. Nature, 1999, 400(6740): 149-151. DOI:10.1038/22087 |
| [20] |
Yang B, He M H, Shishov V, Tychkov I, Vaganov E, Rossi S, Ljungqvist F C, Bräuning A, Grießinger J. New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(27): 6966-6971. DOI:10.1073/pnas.1616608114 |
| [21] |
贺敏慧, 杨保. 树轮生理模型研究进展——以V-S, TREE-RING和MAIDENiso模型为例. 第四纪研究, 2015, 35(5): 1261-1270. |
| [22] |
张永香, 邵雪梅, 徐岩, Wilmking M. 利用生理模型模拟的柴达木东北缘祁连圆柏对气候要素的响应过程. 科学通报, 2011, 56(12): 971-978. |
| [23] |
Allen S T, Keim R F. Wetland-tree growth responses to hydrologic variability derived from development and optimization of a non-linear radial growth model. Ecological Modelling, 2017, 354: 49-61. DOI:10.1016/j.ecolmodel.2017.03.016 |
| [24] |
Shi J F, Liu Y, Vaganov E A, Li J B, Cai Q F. Statistical and process-based modeling analyses of tree growth response to climate in semi-arid area of north central China:a case study of Pinus tabulaeformis. Journal of Geophysical Research:Biogeosciences, 2008, 113(G1): G01026. |
| [25] |
史江峰, 刘禹, Vaganov E, 蔡秋芳, Shishov V. 贺兰山油松生长的气候响应机制初步探讨. 第四纪研究, 2005, 25(2): 245-251. DOI:10.3321/j.issn:1001-7410.2005.02.016 |
| [26] |
Zhang J Z, Gou X H, Zhang Y X, Lu M, Xu X Y, Zhang F, Liu W H, Gao L L. Forward modeling analyses of Qilian Juniper (Sabina przewalskii) growth in response to climate factors in different regions of the Qilian Mountains, northwestern China. Trees, 2016, 30(1): 175-188. DOI:10.1007/s00468-015-1286-0 |
| [27] |
曾立雄, 雷蕾, 王晓荣, 朱建华, 肖文发, 刘贤德, 敬文茂. 海拔梯度对祁连山青海云杉林乔木层和土壤层碳密度的影响. 生态学报, 2018, 38(20): 7168-7177. |
| [28] |
张雷.祁连山青海云杉林结构和树木生长随海拔的变化[D].北京: 中国林业科学研究院, 2015. http://cdmd.cnki.com.cn/Article/CDMD-82201-1015622663.htm
|
| [29] |
Sidor C G, Popa I, Vlad R, Cherubini P. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees, 2015, 29(4): 985-997. DOI:10.1007/s00468-015-1178-3 |
| [30] |
苏凯, 白红英, 张扬, 黄晓月, 秦进. 基于树轮-气候资料的160多年来秦岭太白山降水变化特征重建. 生态学杂志, 2018, 37(5): 1467-1475. |
| [31] |
霍玉侠.树轮宽度记录的新疆北疆地区气候变化研究[D].兰州: 兰州大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10730-1017714224.htm
|
| [32] |
张瑞波.基于树轮的中亚西天山干湿变化研究[D].兰州: 兰州大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10730-1018803985.htm
|
| [33] |
李颖俊.祁连山地区树轮记录的气候变化研究[D].兰州: 兰州大学, 2011. http://cdmd.cnki.com.cn/article/cdmd-10730-1011140239.htm
|
| [34] |
Allen C D, Breshears D D, McDowell N G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere, 2015, 6(8): 1-55. |
| [35] |
Bandurska H, Cieślak M. The interactive effect of water deficit and UV-B radiation on salicylic acid accumulation in barley roots and leaves. Environmental and Experimental Botany, 2013, 94: 9-18. DOI:10.1016/j.envexpbot.2012.03.001 |
| [36] |
Ordoñez J C, van Bodegom P M, Witte J P M, Wright I J, Reich P B, Aerts R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecology and Biogeography, 2009, 18(2): 137-149. DOI:10.1111/j.1466-8238.2008.00441.x |
| [37] |
Ramirez-Valiente J A, Cavender-Bares J. Evolutionary trade-offs between drought resistance mechanisms across a precipitation gradient in a seasonally dry tropical oak (Quercus oleoides). Tree Physiology, 2017, 37(7): 889-901. DOI:10.1093/treephys/tpx040 |
| [38] |
白志远, 邢立新, 潘军, 李鹏飞, 许振北. 土壤湿度信息遥感研究. 测绘与空间地理信息, 2011, 34(3): 50-55. DOI:10.3969/j.issn.1672-5867.2011.03.016 |
| [39] |
Babst F, Alexander M R, Szejner P, Bouriaud O, Klesse S, Roden J, Ciais P, Poulter B, Frank D, Moore D J P, Trouet V. A tree-ring perspective on the terrestrial carbon cycle. Oecologia, 2014, 176(2): 307-322. DOI:10.1007/s00442-014-3031-6 |
| [40] |
Vaganov E A, Hughes M K, Shashkin A V. Growth Dynamics of Conifer Tree Rings. Berlin: Springer, 2006: 183.
|
| [41] |
Wu X C, Liu H Y, Wang Y F, Deng M H. Prolonged limitation of tree growth due to warmer spring in semi-arid mountain forests of Tianshan, northwest China. Environmental Research Letters, 2013, 8(2): 024016. DOI:10.1088/1748-9326/8/2/024016 |
| [42] |
陈峰, 袁玉江, 魏文寿, 喻树龙, 范子昂, 张瑞波, 尚华明, 张同文. 树轮记录的甘肃山丹近224年降水变化. 地理与地理信息科学, 2010, 26(5): 82-86. |
| [43] |
刘晶晶. 树轮记录的青藏高原南部过去526年降水变化特征. 兰州大学学报:自然科学版, 2014, 50(3): 293-298. |
| [44] |
尚华明, 魏文寿, 袁玉江, 喻树龙, 陈霞, 张同文, 刘新华. 树轮记录的中天山150年降水变化特征. 干旱区研究, 2010, 27(3): 443-449. |
| [45] |
张芬, 勾晓华, 苏军德, 高琳琳, 刘文火, 满自红. 祁连山东部不同树龄油松径向生长对气候的响应. 冰川冻土, 2011, 33(3): 634-639. |
| [46] |
Choat B, Brodribb T J, Brodersen C R, Duursma R A, López R, Medlyn B E. Triggers of tree mortality under drought. Nature, 2018, 558(7711): 531-539. DOI:10.1038/s41586-018-0240-x |
| [47] |
McDowell N G, Beerling D J, Breshears D D, Fisher R A, Raffa K F, Stitt M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends in Ecology & Evolution, 2011, 26(10): 523-532. |
| [48] |
关义新, 戴俊英, 林艳. 水分胁迫下植物叶片光合的气孔和非气孔限制. 植物生理学通讯, 1995, 31(4): 293-297. |
| [49] |
张津林, 张志强, 查同刚, 陈吉泉, 孙阁, 刘晨峰, 崔令军, 陈军, 申李华. 沙地杨树人工林生理生态特性. 生态学报, 2006, 26(5): 1523-1532. DOI:10.3321/j.issn:1000-0933.2006.05.030 |
| [50] |
Billings S A, Boone A S, Stephen F M. Tree-ring δ13C and δ18O, leaf δ13C and wood and leaf N status demonstrate tree growth strategies and predict susceptibility to disturbance. Tree Physiology, 2016, 36(5): 576-588. DOI:10.1093/treephys/tpw010 |
| [51] |
Dufour B, Morin H. Tracheid production phenology of Picea mariana and its relationship with climatic fluctuations and bud development using multivariate analysis. Tree Physiology, 2010, 30(7): 853-865. DOI:10.1093/treephys/tpq046 |
| [52] |
Gričar J, Zavadlav S, Jyske T, Lavrič M, Laakso T, Hafner P, Eler K, Vodnik D. Effect of soil water availability on intra-annual xylem and phloem formation and non-structural carbohydrate pools in stem of Quercus pubescens. Tree Physiology, 2018, 39(2): 222-233. |
| [53] |
Simard S, Giovannelli A, Treydte K, Traversi M L, King G M, Frank D, Fonti P. Intra-annual dynamics of non-structural carbohydrates in the cambium of mature conifer trees reflects radial growth demands. Tree Physiology, 2013, 33(9): 913-923. DOI:10.1093/treephys/tpt075 |
| [54] |
田沁花, 周秀骥, 勾晓华, 赵平, 范泽鑫.祁连山中部近500年来降水重建序列分析//第三届区域气候变化监测与检测学术研讨会论文集.呼和浩特: 中国气象学会, 中国气象局气候研究开放实验室, 内蒙古自治区气象局, 2014.
|
| [55] |
张瑞波, 袁玉江, 魏文寿, 尚华明, 喻树龙, 张同文, 陈峰, 范子昂, 秦莉. 西伯利亚落叶松树轮稳定碳同位素对气候的响应. 干旱区研究, 2012, 29(2): 328-334. |
| [56] |
Michelot A, Simard S, Rathgeber C, Dufrêne E, Damesin C. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiology, 2012, 32(8): 1033-1045. DOI:10.1093/treephys/tps052 |