 2020, Vol. 40
2020, Vol. 40文章信息
- 薛晨阳, 高英美, 曲波
- XUE Chenyang, GAO Yingmei, QU Bo
- 镉胁迫对瘤突苍耳、苍耳及其杂合体种子萌发与幼苗生长的影响
- Effects of cadmium on seed germination and seedling growth of Xanthium strumarium, X. sibiricum, and their hybrids
- 生态学报. 2020, 40(1): 345-355
- Acta Ecologica Sinica. 2020, 40(1): 345-355
- http://dx.doi.org/10.5846/stxb201810092176
-
文章历史
- 收稿日期: 2018-10-09
- 网络出版日期: 2019-10-25





2. 辽宁省全球变化与生物入侵重点实验室, 沈阳 110161
2. Liaoning Key Laboratory of Biological Invasions and Global Changes, Shenyang 110161, China
污水灌溉、金属开采、冶炼及加工是造成我国土壤重金属污染的最大原因[1]。日益增加的土壤重金属污染不仅严重影响了作物的产量和质量, 而且通过食物链和食物网向上传递, 影响了动物和人类健康[2]。对受重金属污染的土壤进行修复迫在眉睫, 植物修复技术是指利用植物吸收、降解、挥发、过滤、固定等作用来净化土壤、水体中的重金属元素, 其具有成本低, 操作简单, 对土壤扰动低, 无二次污染等优点[3-4]。目前在实际应用中已经取得一定的成效。比如Ji等在沈阳张士污灌区使用龙葵(Solanum nigrum)进行了为期两年的原位植物修复试验, 发现地表 20 cm土层中的重金属镉浓度由最初的2.75 mg/kg降低到1.53 mg/kg, 降幅达到24.9%[5]。
生物量大、生长迅速、适应性广且能够富集或超富集一种或多种重金属的植物是植物修复技术的关键[6]。目前发现的一些重金属富集植物一般都存在一定的缺点, 不利于大规模推广应用。例如杨肖娥等发现东南景天(Sedum alfredii)对土壤中高含量的Zn有很强的忍耐、吸收和积累能力, 当土壤中Zn含量为2269—3858 mg/kg, 植株地上部分Zn含量达到4065—5000 mg/kg, 富集系数为1.25—1.94[7], 但东南景天在寒冷高海拔地区不开花, 生长缓慢。同样, Visoottiviseth等发现蜈蚣草(Pteris vittata)对土壤中高含量的As具有很强的吸收能力, 当土As壤浓度为810—1400 mg/kg, 蜈蚣草羽片As含量达到4240—6030 mg/kg, 富集系数达到4.31—5.23[8], 但蜈蚣草在淮河以北及沈阳地区不能生长。
现有的超富集植物仍难以满足我们的实践需求, 超富集植物种质资源库仍需要持续扩充。杂交是自然界中广泛存在的基因交流现象, 也是植物育种中常用的技术, 其涉及到完整染色体上大量基因的重组, 这很容易引起某些生物性状的改变, 比如更快的生长速度、更大的体积以及生物量等特征, 从而使植物适应复杂多变的环境, 提高后代的适合度[9-10]。Lexer等发现入侵种矛盾向日葵(Helianthus paradoxs)是由向日葵(H. annuus)和原野向日葵(H. petiolaris)经天然杂交产生, 获得了双亲均不具有的耐盐性状, 因此能够成功入侵到盐碱性的湿地生境中[11];Lin等从四川两个不同的气候-生态区采集了龙葵进行杂交, 在盆栽实验中发现杂合体提取效率和生物量均高于亲本, 在田间实验中, 杂合体生物量高于亲本且茎秆的镉提取效率显著增加[12]。
通过野外调查, 我们发现沈阳张士污灌区同时存在瘤突苍耳(Xanthium strumarium)和苍耳(Xanthium sibiricum)两种植物, 且两种植物之间有可能存在杂交现象。Jeelani等[13]和胡小英等[14]研究结果分别显示苍耳和瘤突苍耳对重金属镉的转移系数均小于1, 二者将大量的重金属镉聚集在根部。这种类型植物可以用来稳定土壤中重金属, 防治其迁移[15]。瘤突苍耳原产北美洲, 由于抗逆性强、生长快、生物量大的特点, 在我国东北地区已成为危害较重的入侵植物之一。目前, 对瘤突苍耳和苍耳杂合体的研究主要集中在种子萌发特性[16]、对养分响应[17-18]方面, 关于镉胁迫下二者杂合体种子萌发及幼苗生长的研究尚未见报道。
种子是植物生长周期中特殊而重要的阶段, 其对个体繁殖、种群扩展和抵抗不良环境有着重要的意义, 而种子萌发是将植物种群从幕后转变为现实种群的关键环节[19];幼苗生长阶段是植物生活史中对环境生态因子反应最为敏感的时期之一, 植物在该阶段的表现能初步反映其抵抗外界干扰的能力[20]。因此, 本试验通过比较苍耳、瘤突苍耳及其杂合体种子在不同镉浓度处理下萌发及其幼苗生长情况, 以期初步判断杂合体对重金属镉的耐性是否优于亲本, 为超富集植物的筛选提供一种思路。
1 材料与方法 1.1 瘤突苍耳、苍耳及其杂合体种子蛋白质电泳实验 1.1.1 实验材料杂合体亲本瘤突苍耳和苍耳种子分别于2012年11月采于辽宁省凌海市右卫满族镇小黄村(121°34′ E, 41°04′ N)大凌河河岸沙地的自然成熟植株。2017年在沈阳农业大学教学科研基地分别进行瘤突苍耳和苍耳自交及杂交试验, 获得瘤突苍耳自交种子1000粒(采自5株植株), 苍耳自交种子1000粒(采自5株植株), 杂合体(X. strumarium♀ × X. sibiricum♂, 简称杂交瘤突)种子591粒(采自5株植株), 杂合体(X. sibiricum♀ × X. strumarium♂, 简称杂交苍耳)种子935粒(采自5株植株)。用于收集自交种子的瘤突苍耳和苍耳在花期时选择健康的花序进行套袋处理, 防止受到干扰作为母本的瘤突苍耳和苍耳在花期时去除雄蕊, 待柱头呈羽毛状分叉并带有光泽时马上分别授以苍耳和瘤突苍耳的花粉, 授粉后的花序套袋隔离。待花期过后去除自交袋和杂交袋, 待果实自然成熟后, 按株采集, 放置于通风避光处晾干, 室温保存备用。
野外疑似杂合体种子于2017年11月采于辽宁省朝阳市建平县沙海镇(119°27′ E, 41°29′ N), 按株采集, 共收集55株。
1.1.2 实验方法参考Osbome[21]对蛋白质的分类, 将种子蛋白分为水溶蛋白(清蛋白)、盐溶蛋白(球蛋白)、剩余蛋白(胶蛋白和谷蛋白)。取瘤突苍耳、苍耳自交及杂合体种子2—4粒(约0.15 g)。放于-20℃预冷的研钵, 加入0.1 g SiO2, 加入9 mL去离子水于冰盒上研磨, 随后转入10 mL离心管, 8000 r/min低温离心4 min, 取上清200 μL, 移入1.5 mL离心管, 按照1:1比例加入事先准备好的蛋白质提取液(0.15% SDS)200 μL, 振荡混匀, 放入25℃光照培养箱中提取30 min后, 配平, 8000 r/min低温离心4 min, 取上清200 μL, 移入1.5 mL离心管中4℃保存, 作为水溶蛋白提取液;弃掉剩余上清液, 加入9 mL去离子水, 振荡混匀, 在25℃光照培养箱中提取30 min, 配平, 8000 r/min低温离心4 min, 去上清, 再重复一次, 去上清后, 加入9 mL 1 mol/L NaCl溶液, 振荡混匀, 放入25℃光照培养箱中提取30 min, 配平, 8000 r/min低温离心4 min, 取上清200μL, 移入1.5 mL离心管中4℃保存, 作为盐溶蛋白提取液;弃掉剩余上清液, 加9 mL 1mol/L NaCl溶液, 振荡混匀, 在25℃光照培养箱中提取30 min, 配平, 8000 r/min低温离心4 min, 去上清, 再重复一次, 加入9 mL蛋白提取液放入25℃光照培养箱中提取30 min, 配平, 8000 r/min低温离心4 min, 取上清200 μL, 移入1.5 mL离心管中4℃保存, 作为剩余蛋白提取液。
采用10.2%醋酸尿素聚丙烯酰胺凝胶溶液制胶, 2%冰乙酸为电极缓冲液(4℃保存), 水溶、盐溶、剩余蛋白点样量分别为12、5、5 μL, 接通电源, 恒定100V下4小时。
水溶蛋白和剩余蛋白胶片染色12 h, 盐溶蛋白胶片染色5 h;水溶蛋白和剩余蛋白胶片加入脱色液(50%甲醇, 7%的冰醋酸水溶液)洗脱1 min倒掉, 重复3次, 随后加入脱色液, 于恒温25℃摇床45 min;盐溶蛋白胶片加入脱色液洗脱1 min, 重复两次, 加入去离子水洗脱1 min, 重复两次, 随后加入去离子水, 于恒温25℃摇床45 min。将胶片置于胶片观察灯上并照相[22]。实验结果进行4次重复。
野外疑似杂合体种子从55株中随机选择11株, 试验方法同上。
1.2 镉胁迫对瘤突苍耳、苍耳及其杂合体种子萌发影响实验 1.2.1 实验材料选择瘤突苍耳、苍耳自交种及杂合体种子各450粒(按株平均随机选择90粒), 由于瘤突苍耳和苍耳种子具有二形性, 相同条件下, 上位种子休眠时间较下位种子长5—7 d[23]。为了消除上下位种子的影响, 本实验均选择下位种子作为实验材料。
1.2.2 实验方法首先用5%的NaClO溶液浸泡种子10 min, 去离子水冲洗5—6次, 然后将消过毒的种子放入铺有2层滤纸的培养皿中(直径9 cm, 使用前均高温高压蒸汽120℃, 30 min灭菌), 每个培养皿放30粒种子。随后将培养皿置于恒温光照培养箱中, 在25℃, 12 h光照/12h黑暗的条件下进行萌发。采用CdCl2 ·2.5H2O(分析纯)设置0(CK)、10、40、80、120 mg/L 5个镉浓度, 每个处理3次重复。每个浓度均采用蒸馏水作为基础液, 对照组施以蒸馏水。为保持溶液浓度恒定, 每隔24 h更换1次处理液和滤纸。
1.2.3 指标测定
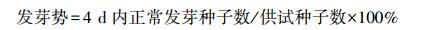
|
(1) |
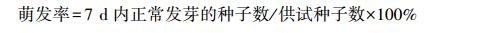
|
(2) |
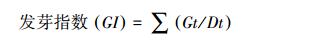
|
(3) |
式中, GI为在t日的发芽种子数;Dt为发芽天数[24]。
1.3 镉胁迫对瘤突苍耳、苍耳及其杂合体幼苗生长影响实验 1.3.1 实验材料待种子萌发实验进行7 d后, 将不同镉处理和对照的种子萌发材料转移至直径15 cm培养皿, 施以相同浓度的镉处理液和蒸馏水, 继续培养3天, 每隔24 h更换1次处理液和滤纸。
1.3.2 实验方法第10天, 用游标卡尺(精度0.01 mm)测量不同镉浓度处理下瘤突苍耳、苍耳及其杂合体幼苗子叶长度和根长, 随后120℃杀青15 min, 60℃烘箱中烘干后称重, 测定地上部分干重、地下部分干重并计算总干重;采用硫代巴比妥酸(TBA)比色法测定植物子叶丙二醛含量[25];采用愈创木酚法测定植物子叶愈创木酚过氧化物酶(GPX)活性[25];采用80%丙酮提取植物子叶叶绿素, 并利用紫外光分光光度计测定470、646、663 nm波长下吸光度, 根据Lichtenthaler和Wellburn[26]公式计算植物叶绿素含量。以上指标均随机选择3株幼苗。
1.4 数据处理采用双因素方差分析(two-way ANOVA)分析杂交、镉浓度及二者交互作用对瘤突苍耳、苍耳种子萌发和幼苗生长的影响;随后, 在双因素方差分析的基础上进行多重比较(LSD)。采用SPSS 22.0进行统计分析, Origin 9.0作图。
2 结果与分析 2.1 瘤突苍耳、苍耳及其杂合体与野外疑似杂合体种子蛋白质电泳瘤突苍耳与苍耳盐溶蛋白条带存在显著差异(图 1绿色箭头), 杂交瘤突和杂交苍耳的水溶蛋白、剩余蛋白分别与瘤突苍耳和苍耳无明显差异(未放入)。杂交瘤突和杂交苍耳的盐溶蛋白同时具有亲本瘤突苍耳和苍耳独有的蛋白条带(图 1红色箭头), 说明杂合体种子确实为瘤突苍耳和苍耳杂交而来。

|
| 图 1 人工杂合体和亲本种子盐溶蛋白聚丙烯酰胺凝胶电泳图 Fig. 1 Salt-protein polyacrylamide gel electrophoresis diagram of artificial hybrids and parental seeds 从左往右依次为瘤突苍耳X. strumarium, 杂交瘤突X. strumarium♀ × X. sibiricum♂, 杂交苍耳X. sibiricum♀ × X. strumarium♂, 苍耳 X. sibiricum |
疑似杂合体种子蛋白质盐溶蛋白电泳结果如图 2所示, 部分疑似杂合体种子的盐溶蛋白条带与人工杂合体情况类似(图 2红色箭头), 其水溶蛋白和剩余蛋白无明显差异(未放入), 说明在自然界中, 瘤突苍耳和苍耳之间存在杂交现象。

|
| 图 2 辽宁省朝阳市建平县沙海镇疑似杂合体种子盐溶蛋白电泳图 Fig. 2 Salt-soluble protein electrophoresis map of suspected hybrids in Shahai Town, Jianping County, Chaoyang City, Liaoning Province 从左往右前2个条带:苍耳; 中间条带:疑似杂交种; 最后2个条带:瘤突苍耳The first two strip from left to right: X. sibiricum; Middle strip: Suspected hybrid; Last two strip: X. strumarium |
杂交对瘤突苍耳种子的萌发率、根长、地下部鲜重、总鲜重、地上部干重、总干重及子叶丙二醛含量有显著影响(P < 0.05, 表 1), 镉浓度则对除发芽指数、子叶长、地上部鲜重及总干重外的其余指标均有显著形影(P < 0.05, 表 1), 二者交互作用对瘤突苍耳的根长、丙二醛、叶绿素含量存在显著影响(P < 0.05, 表 1)。杂交对苍耳的根长、子叶长、丙二醛含量、愈创木酚过氧化物酶活性、叶绿素含量有显著的影响(P < 0.05, 表 1), 镉浓度对根长、子叶长、地上、地下及总鲜重、丙二醛含量、愈创木酚过氧化物酶活性、叶绿素含量均有显著影响(P < 0.05, 表 1), 二者交互作用对苍耳的根长、丙二醛含量、愈创木酚过氧化物酶活性、叶绿素含量存在显著影响(P < 0.05, 表 1)。
| 杂交Hybridization (H) | 镉浓度Cd | 交互作用H × Cd | ||||||
| F | P | F | P | F | P | |||
| 瘤突苍耳X. strumarium | ||||||||
| 萌发率Germination rate | 12.281 | 0.002* | 5.467 | 0.004* | 2.550 | 0.071 | ||
| 发芽势Germination energy | 0.007 | 0.933 | 4.037 | 0.015* | 1.839 | 0.161 | ||
| 发芽指数Germination index | 2.071 | 0.166 | 1.532 | 0.231 | 2.619 | 0.066 | ||
| 根长Root length | 29.944 | 0.000* | 601.079 | 0.000* | 28.464 | 0.000* | ||
| 子叶长Cotyledon length | 2.481 | 0.117 | 96.081 | 0.000* | 1.941 | 0.105 | ||
| 地上部鲜重Aboveground fresh weight | 3.621 | 0.065 | 14.489 | 0.000* | 1.582 | 0.201 | ||
| 地下部鲜重Underground fresh weight | 5.817 | 0.021* | 37.530 | 0.000* | 1.259 | 0.304 | ||
| 总鲜重Total fresh weight | 4.685 | 0.037* | 21.750 | 0.000* | 1.170 | 0.341 | ||
| 地上部干重Aboveground dry weight | 12.210 | 0.001* | 4.054 | 0.009* | 1.092 | 0.378 | ||
| 地下部干重Underground dry weight | 0.040 | 0.843 | 22.743 | 0.000* | 0.348 | 0.843 | ||
| 总干重Total dry weight | 10.273 | 0.003* | 1.073 | 0.386 | 0.909 | 0.471 | ||
| 丙二醛MDA, Malondialdehyde | 35.476 | 0.000* | 19.564 | 0.000* | 12.493 | 0.000* | ||
| 愈创木酚过氧化物酶GPX, Guaiacol Peroxidase | 1.301 | 0.267 | 4.087 | 0.014* | 0.416 | 0.795 | ||
| 叶绿素含量Chlorophyll content | 0.026 | 0.874 | 41.854 | 0.000* | 3.029 | 0.042* | ||
| 苍耳 X. sibiricum | ||||||||
| 萌发率Germination rate | 0.079 | 0.782 | 0.062 | 0.992 | 0.667 | 0.622 | ||
| 发芽势Germination energy | 1.472 | 0.239 | 0.305 | 0.871 | 1.144 | 0.365 | ||
| 发芽指数Germination index | 1.239 | 0.279 | 0.041 | 0.997 | 1.246 | 0.324 | ||
| 根长Root length | 11.395 | 0.001* | 73.053 | 0.000* | 14.042 | 0.000* | ||
| 子叶长Cotyledon length | 5.287 | 0.024* | 9.061 | 0.000* | 1.248 | 0.297 | ||
| 地上部鲜重Aboveground fresh weight | 1.760 | 0.196 | 4.993 | 0.004* | 0.909 | 0.473 | ||
| 地下部鲜重Underground fresh weight | 0.609 | 0.442 | 9.856 | 0.000* | 0.965 | 0.443 | ||
| 总鲜重Total fresh weight | 0.565 | 0.459 | 6.653 | 0.001* | 0.965 | 0.443 | ||
| 地上部干重Aboveground dry weight | 2.263 | 0.145 | 1.234 | 0.322 | 1.220 | 0.327 | ||
| 地下部干重Underground dry weight | 4.099 | 0.054 | 1.210 | 0.331 | 1.036 | 0.408 | ||
| 总干重Total dry weight | 2.788 | 0.107 | 1.420 | 0.256 | 1.436 | 0.251 | ||
| 丙二醛MDA, Malondialdehyde | 11.709 | 0.003* | 5.175 | 0.005* | 6.118 | 0.002* | ||
| 愈创木酚过氧化物酶GPX, Guaiacol Peroxidase | 47.807 | 0.000* | 7.920 | 0.001* | 10.895 | 0.000* | ||
| 叶绿素含量Chlorophyll content | 76.117 | 0.000* | 234.660 | 0.000* | 2 9.720 | 0.000* | ||
| *: P < 0.05 | ||||||||
苍耳、瘤突苍耳及杂交瘤突苍耳种子萌发率随着Cd浓度的增加而增加。在5个Cd浓度下, 瘤突苍耳和杂交瘤突苍耳种子萌发率均显著高于苍耳和杂交苍耳(P < 0.05), 苍耳和杂交苍耳种子萌发率无显著差异(P > 0.05);在80 mg/L Cd处理下, 杂交瘤突苍耳种子萌发率显著高于瘤突苍耳, 其余处理下则无显著性差异(P < 0.05)。在5个Cd浓度下, 瘤突苍耳和杂交瘤突苍耳种子发芽指数均显著高于苍耳和杂交苍耳(P < 0.05);在80 mg/L Cd处理下, 杂交瘤突苍耳种子发芽指数显著高于瘤突苍耳, 其余处理下则无显著性差异(P > 0.05);在120 mg/L Cd处理下, 杂交苍耳种子发芽势显著低于苍耳(P < 0.05), 其余处理下则无显著性差异(P > 0.05)。苍耳、瘤突苍耳及其杂合体种子发芽势表现出与发芽指数一致的规律性。
苍耳、瘤突苍耳及其杂合体幼苗根长和子叶长度随着Cd浓度的增加而降低(图 4)。杂交瘤突苍耳和杂交苍耳子叶长在Cd浓度40 mg/L处理下分别显著低于瘤突苍耳和苍耳(P < 0.05);杂交瘤突苍耳根长在CK和10 mg/L处理下显著高于瘤突苍耳(P < 0.05), 在40—120 mg/L处理下显著低于瘤突苍耳(P < 0.05), 在80和120 mg/L下显著高于苍耳(P < 0.05);杂交苍耳根长在40 mg/L处理下显著低于苍耳(P < 0.05)。

|
| 图 3 不同镉浓度下瘤突苍耳、苍耳及其杂合体种子萌发特征 Fig. 3 Germination feature of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE) 在同一镉浓度处理下, 不同小写字母代表苍耳、瘤突苍耳及其杂合体存在显著差异(P < 0.05)。CK:蒸馏水; 10:镉浓度10 mg/L; 40:镉浓度40 mg/L; 80:镉浓度80 mg/L; 120:镉浓度120 mg/L |

|
| 图 4 不同镉浓度下瘤突苍耳、苍耳及其杂合体根长和子叶长 Fig. 4 Root length and cotyledon length of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE) |
瘤突苍耳、苍耳及其杂合体的地上鲜重、地下鲜重、总鲜重及地下干重随着Cd浓度的增加而降低, 其地上干重和总干重则随着Cd浓度的增加而增加。杂交苍耳的地上鲜重和总鲜重仅在镉浓度为120 mg/L时显著低于苍耳(P < 0.05), 地上、地下及总鲜重在CK下与苍耳无显著差异(P > 0.05), 但亲本瘤突苍耳显著高于苍耳(P < 0.05), 说明在CK处理下, 杂交使得苍耳鲜重增加;杂交苍耳的地上干重和总干重在CK和80 mg/L时与苍耳无显著差异(P > 0.05), 其余浓度处理均显著低于苍耳(P < 0.05);杂交瘤突苍耳幼苗地上鲜重和总鲜重在镉浓度为10 mg/L时显著低于瘤突苍耳(P < 0.05), 地上干重和总干重在镉浓度为40和120 mg/L时显著低于瘤突苍耳(P < 0.05), 地下干重在120 mg/L时显著低于瘤突苍耳(P < 0.05)。
苍耳、瘤突苍耳及其杂合体幼苗子叶丙二醛含量和愈创木酚过氧化物酶活性随着Cd浓度的增加而增加, 叶绿素含量则相反(图 6)。对照组中, 杂交瘤突苍耳和杂交苍耳的叶绿素总含量分别显著高于瘤突苍耳和苍耳(P < 0.05), 在80 mg/L处理下则相反(P < 0.05);在10、40、80 mg/L处理下, 瘤突苍耳的丙二醛含量显著高于杂交瘤突(P < 0.05), 仅在80 mg/L处理下, 杂交苍耳的丙二醛含量显著高于苍耳(P < 0.05);杂交瘤突苍耳和杂交苍耳的愈创木酚过氧化物酶活性在对照处理下分别显著高于瘤突苍耳和苍耳(P < 0.05);在10、80、120 mg/L处理下, 杂交苍耳的愈创木酚过氧化物酶活性显著低于苍耳(P < 0.05)。

|
| 图 5 不同镉浓度下瘤突苍耳、苍耳及其杂合体生物量特征 Fig. 5 Biomass characteristics of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE) |

|
| 图 6 不同镉浓度下瘤突苍耳、苍耳及其杂合体叶绿素含量、丙二醛及愈创木酚过氧化物酶活性 Fig. 6 Chlorophyll content, malondialdehyde (MDA) and Guaiacol peroxidase activity (POD) of X. strumarium, X. sibiricum and their hybrids under different Cd concentration (means±SE) |
种子蛋白质电泳实验结果显示, 苍耳和瘤突苍耳的杂交后代具有亲本独有的某些盐溶蛋白, 这说明杂交使得二者基因之间发生交换和重组, 随后双因素方差分析表明杂交确实影响了瘤突苍耳和苍耳的某些性状, 但这些变化性状是否有增强了植物对重金属镉的耐受性, 我们仍需要继续进行验证。由于杂交亲本筛选比较困难、杂交后代形状不稳定、杂种优势受重组、杂合性下降等因素随世代的增加而消失以及较长的研究周期, 目前很少有研究通过入侵植物与本地种杂交来进行超富集植物的筛选, 但植物对重金属的积累受控于体内特有基因表达。比如Liu等[27]对镉锌超积累植物伴矿景天Cd耐性基因进行研究, 发现伴矿景天体内的ATP酶3基因(SpHMA3)具有Cd专一运输活性, 在非超积累型景天中超量表达可增加其对镉的耐性和积累性。因此利用杂交改变植物的遗传特征, 这值得我们进行尝试。
萌发率反映了种子的生命力, 发芽势反映种子出苗快慢, 两者均为鉴定种子质量最直接和稳定的指标, 发芽指数则反映了整个发芽期种子活力的综合特征[28]。本试验结果显示, 瘤突苍耳的发芽率、发芽势及发芽指数随着重金属镉浓度的增加而增加, 说明镉胁迫并未抑制其种子活力, 相反还提高了其发芽速度, 这与胡小英等[29]研究结果一致。杂交瘤突(X. strumarium♀ × X. sibiricum♂)种子的萌发情况与瘤突苍耳类似, 且当镉浓度为80 mg/L时, 杂交瘤突(X. strumarium♀ × X. sibiricum♂)种子的萌发率显著高于瘤突苍耳并达到最大(83.33%), 这说明了以瘤突苍耳为母本, 苍耳为父本的杂交后代种子在萌发情况上得到提高, 并未受到抑制。相反, 杂交苍耳(X. sibiricum♀ ×X. strumarium ♂)不仅在种子萌发率上低于苍耳, 而且在高浓度(120 mg/L)时种子发芽势和发芽指数均显著低于苍耳, 说明以苍耳为母本, 瘤突苍耳为父本的杂交后代种子在高浓度镉胁迫下的表现弱于苍耳。
幼苗的生长状况是植物对逆境胁迫适应的综合体现, 胁迫环境对植物生长的抑制程度直观地反映了植物体的受伤程度[30]。本研究结果表明, 苍耳、瘤突苍耳及杂合体幼苗的根长、子叶长、鲜重、叶绿素含量随着镉浓度的增加而降低, 说明重金属镉对幼苗的生长具有明显的抑制作用。这与张大鹏等[31]、韩宝贺等[32]的研究结果一致。随着镉浓度增加, 苍耳、瘤突苍耳及其杂合体根长降幅远高于子叶, 这是因为在种子萌发过程中, 胚根是主要吸收水分和养分的器官, 长时间与镉接触, 所以根长受到的抑制程度高于子叶。
重金属镉会导致植物体内自由基和活性氧含量大幅度上升, 进而氧化蛋白质、DNA、脂类和碳水化合物, 从而扰乱许多生理生化过程[33-34]。蔺永和等研究了野生垂穗披碱草(Elymus nutans)对铝胁迫的响应, 发现随着铝浓度的增加, 丙二醛含量不断增加[35]。张虎等研究了紫花苜蓿种子萌发(Medicago sativa)对钴胁迫的生理生化响应, 发现MDA含量随着钴浓度的增加而增加[24]。本研究结果与这些研究基本一致。苍耳和杂交苍耳的GPX活性持续增加, 但在10、40、80、120 mg/L下, 苍耳的GPX却高于杂交苍耳(仅在40 mg/L处理下不显著), 说明苍耳的抗氧化能力高于杂交苍耳。
本试验基础液均采用蒸馏水, 排除了不同离子对幼苗生长的影响, 植物生长发育取决于植物本身。结果显示, 在对照和低浓度(10 mg/L)胁迫下, 杂交瘤突幼苗根长和子叶叶绿素含量均高于瘤突苍耳;在高浓度镉(80、120 mg/L)胁迫下, 杂交瘤突幼苗根长和子叶叶绿素含量低于瘤突苍耳。Mobin和Khan研究了Varuna和RH-30两种芥菜(Brassica juncea)在镉胁迫下的光合活性、色素组成及抗氧化反应, 结果显示当土壤镉浓度为25、50、100 mg/kg时, RH-30根和叶中的镉浓度均显著高于Varuna, 但Varuna中叶绿素总含量分别降低20.0%, 35.0%和50.0%, RH-30则分别降低31.0%, 43.0%和57.0%[36]。相似的结果也在Shi等[37]、He等[38]、Zhang等[39]、Fang等[40]研究中发现。前人研究结果已经证明镉可以通过抑制叶绿素合成酶和CO2固定酶的活性[36, 41]或光系统色素蛋白复合物的聚集[42]来扰乱叶绿体功能。结合我们的实验结果, 我们推测杂交有可能提高了瘤突苍耳在高浓度镉胁迫时对重金属镉的转运能力, 幼苗根系吸收大量的镉并向上运输给子叶, 这导致根系生长受到抑制、子叶叶绿素含量下降。同时我们发现当镉浓度为10、40、80 mg/L时, 杂交瘤突幼苗MDA含量显著低于瘤突苍耳, 其GPX活性高于瘤突苍耳, 但并不显著, 因此杂交瘤突抗氧化酶系统是否在这一过程中发挥了作用仍需要我们继续研究。杂交是否提高了瘤突苍耳对重金属镉的吸收与转运, 我们仍然需要后续盆栽实验给出确切的证据, 同时深入探究杂交在这一过程中发挥的作用。
| [1] |
环境保护部国土资源部发布全国土壤污染状况调查公报.中华人民共和国国土资源部. (2014-04-17)[2018-10-09].http://www.mlr.gov.cn/xwdt/jrxw/201404/t20140417_1312998.htm.
|
| [2] |
Zhao F J, Ma Y, Zhu Y G, Tang Z, McGrath S P. Soil contamination in China:current status and mitigation strategies. Environmental Science & Technology, 2014, 49(2): 750-759. |
| [3] |
安婧, 宫晓双, 魏树和. 重金属污染土壤超积累植物修复关键技术的发展. 生态学杂志, 2015, 34(11): 3261-3270. |
| [4] |
王庆海, 却晓娥. 治理环境污染的绿色植物修复技术. 中国生态农业学报, 2013, 21(2): 261-266. |
| [5] |
Ji P H, Song Y F, Jiang Y L, Tang X W, Tong Y A, Gao P C, Han W S. A two-year field study of phytoremediation using Solanum nigrum L. in China. International Journal of Phytoremediatio, 2016, 18(9): 924-928. DOI:10.1080/15226514.2015.1058325 |
| [6] |
陈文麟. 土壤重金属污染及植物修复技术综述. 科技创新与应用, 2015(25): 184-184. |
| [7] |
杨肖娥, 龙新宪, 倪吾钟, 傅承新. 东南景天(Sedum alfredii H)——一种新的锌超积累植物. 科学通报, 2002, 47(13): 1003-1006. DOI:10.3321/j.issn:0023-074X.2002.13.010 |
| [8] |
Visoottiviseth P, Francesconi K, Sridokchan W. The potential of Thai indigenous plant species for the phytoremediation of arsenic contaminated land. Environmental Pollution, 2002, 118(3): 453-461. DOI:10.1016/S0269-7491(01)00293-7 |
| [9] |
李宏, 许惠. 外来物种入侵科学导论. 北京: 科学出版社, 2016: 10-16.
|
| [10] |
万方浩, 侯有明, 蒋明星. 入侵生物学. 北京: 科学出版社, 2015: 69-73.
|
| [11] |
Lexer C, Welch M E, Durphy J L, Rieseberg L H. Natural selection for salt tolerance quantitative trait loci (QTLs) in wild sunflower hybrids:Implications for the origin of Helianthus paradoxus, a diploid hybrid species. Molecular Ecology, 2003, 12(5): 1225-1235. DOI:10.1046/j.1365-294X.2003.01803.x |
| [12] |
Lin L J, He J, Wang X., Wang J, Lv X L, Liao M A, Wang Z H, Tang Y, Liang D, Xia H, Lai Y S. Cadmium accumulation characteristics of F1 hybrids by reciprocal hybridizing of Solanum nigrum in two climate-ecology regions. Environmental Science and Pollution Research, 2016, 23(18): 18842-18849. DOI:10.1007/s11356-016-7089-z |
| [13] |
Jeelani N, Yang W, Qiao Y J, Li J J, An S Q, Leng X. Individual and combined effects of cadmium and polycyclic aromatic hydrocarbons on the phytoremediation potential of Xanthium sibiricum in co-contaminated soil. International Journal of Phytoremediation, 2018, 20(8): 773-779. DOI:10.1080/15226514.2018.1425666 |
| [14] |
胡小英, 齐淑艳, 李瑞莉. 镉胁迫对意大利苍耳(Xanthium italicum)生长、叶绿素及光合特性的影响. 沈阳大学学报(自然科学版), 2018, 30(1): 18-24. |
| [15] |
Pignattelli S, Colzi I, Buccianti A, Cecchi L, Arnetoli M, Monnanni R, Gabbrielli R, Gonnelli C. Exploring element accumulation patterns of a metal excluder plant naturally colonizing a highly contaminated soil. Journal of Hazardous Materials, 2012, 227-228: 362-369. DOI:10.1016/j.jhazmat.2012.05.075 |
| [16] |
曲波, 荀挚峰, 许玉凤. 瘤突苍耳入侵的种子生态学机制. 草业科学, 2015, 32(11): 1801-1807. DOI:10.11829/j.issn.1001-0629.2015-0354 |
| [17] |
荀挚峰, 白龙, 曲波, 许玉凤, 李光海, 詹忠浪, 石九曜. 不同氮添加对入侵植物瘤突苍耳和本地近缘植物苍耳及两者杂交种的生长影响. 草业学报, 2017, 26(5): 51-61. |
| [18] |
薛晨阳, 许玉凤, 曲波. 不同氮水平下瘤突苍耳、苍耳及其杂交种形态、光合及生长特征比较. 生物多样性, 2018, 26(6): 554-563. |
| [19] |
叶平扬, 董姗姗, 卢宝荣, 王锋, 苏军, 宋志平. 转基因水稻与野生稻(Oryza nivara Sharma et Shastry)杂交后代种子萌发率. 复旦学报:自然科学版, 2008, 47(3): 329-335. |
| [20] |
徐承远, 张文驹, 卢宝荣, 陈家宽. 生物入侵机制研究进展. 生物多样性, 2001, 9(4): 430-438. DOI:10.3321/j.issn:1005-0094.2001.04.016 |
| [21] |
Osborne T B. The Proteins of the Wheat Kernel. Washington, DC: Publication of the Carnegie Institution, 1907.
|
| [22] |
郑建鹏.向日葵种子纯度室内快速检测方法的研究[D].泰安: 山东农业大学, 2015.
|
| [23] |
吴冬, 黄姝博, 李宏庆. 意大利苍耳二形性种子萌发、植株生长差异及生态适应性. 生态学报, 2009, 29(10): 5258-5264. DOI:10.3321/j.issn:1000-0933.2009.10.011 |
| [24] |
张虎, 寇江涛, 师尚礼. 紫花苜蓿种子萌发对钴胁迫的生理生化响应. 草业学报, 2015, 24(9): 146-153. |
| [25] |
樊金娟, 阮燕晔. 植物生理学实验教程(第二版). 北京: 中国农业大学出版社, 2015.
|
| [26] |
Linchenthaler H R, Wellburn A R. Determination of total carotenoides and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions, 1983, 11(5): 591-592. DOI:10.1042/bst0110591 |
| [27] |
Liu H, Zhao H X, Wu L H, Liu A N, Zhao F J, Xu W Z. Heavy metal ATPase 3(HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytologist, 2017, 215(2): 687-698. DOI:10.1111/nph.14622 |
| [28] |
潘高, 张合平, 刘鹏, 刘文胜. 锰胁迫对苍耳种子萌发及幼苗生理生化特性的影响. 草业学报, 2017, 26(11): 157-166. DOI:10.11686/cyxb2017033 |
| [29] |
胡小英, 齐淑艳, 秦子晴. 镉胁迫对意大利苍耳种子萌发及幼根生理特性的影响. 沈阳大学学报:自然科学版, 2018, 30(3): 200-206. |
| [30] |
Zimmer D, Kruse J, Baum C, Borca C, Laue M, Hause G, Meissner R, Leinweber P. Spatial distribution of arsenic and heavy metals in willow roots from a contaminated floodplain soil measured by X-ray fluorescence spectroscopy. Science of the Total Environment, 2011, 409(19): 4094-4100. |
| [31] |
张大鹏, 蔡春菊, 范少辉, 苏文会. 重金属Pb2+和Cd2+对毛竹种子萌发及幼苗早期生长的影响. 林业科学研究, 2012, 25(4): 500-504. DOI:10.3969/j.issn.1001-1498.2012.04.014 |
| [32] |
韩宝贺, 朱宏, 张艺馨. 镉对三种草坪草种子萌发与生长的影响. 中国草地学报, 2014, 36(3): 98-103. |
| [33] |
Belkhadi A, Hediji H, Abbes Z, Nouairi I, Barhoumi Z, Zarrouk M, Chaïbi W, Djebali W. Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content in Linum usitatissimum L. Ecotoxicology and Environmental Safety, 2010, 73(5): 1004-1011. DOI:10.1016/j.ecoenv.2010.03.009 |
| [34] |
Fernández R, Bertrand A, Reis R, Mourato M P, Martins L L, González A. Growth and physiological responses to cadmium stress of two populations of Dittrichia viscosa (L.) Greuter. Journal of Hazardous Materials, 2013, 244-245: 555-562. DOI:10.1016/j.jhazmat.2012.10.044 |
| [35] |
蔺永和, 吴景, 方江平, 张卫红, 苗彦军, 李勇胜. 铝胁迫对西藏野生垂穗披碱草种子萌发及幼苗生长的影响. 草业学报, 2018, 27(7): 155-165. |
| [36] |
Mobin M, Khan N A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. Journal of Plant Physiology, 2007, 164(5): 601-610. DOI:10.1016/j.jplph.2006.03.003 |
| [37] |
Shi G R, Liu C F, Cai Q S, Liu Q Q, Hou C P. Cadmium accumulation and tolerance of two safflower cultivars in relation to photosynthesis and antioxidantive enzymes. Bulletin of Environmental Contamination and Toxicology, 2010, 85(3): 256-263. DOI:10.1007/s00128-010-0067-0 |
| [38] |
He J L, Ma C F, Ma Y L, Li H, Kang J Q, Liu T X, Polle A, Peng C H, Luo Z B. Cadmium tolerance in six poplar species. Environmental Science and Pollution Research, 2013, 20(1): 163-174. DOI:10.1007/s11356-012-1008-8 |
| [39] |
Zhang X F, Gao B, Xia H P. Effect of cadmium on growth, photosynthesis, mineral nutrition and metal accumulation of bana grass and vetiver grass. Ecotoxicology and Environmental Safety, 2014, 106: 102-108. DOI:10.1016/j.ecoenv.2014.04.025 |
| [40] |
Fang Z G, Lou L Q, Tai Z L, Wang Y F, Yang L, Hu Z B, Cai Q S. Comparative study of Cd uptake and tolerance of two Italian ryegrass (Lolium multiflorum) cultivars. PeerJ, 2017, 5(3): e3621. |
| [41] |
Siedlecka A, Krupa Z, Samuelsson G, Öquist G, Gardeström P. Primary carbon metabolism in Phaseolus vulgaris plants under Cd(II)/Fe interaction. Plant Physiology and Biochemistry, 1997, 35(12): 951-957. |
| [42] |
Horváth G, Droppa M, Oravecz Á, Raskin V I, Marder J B. Formation of the photosynthetic apparatus during greening of cadmium-poisoned barley leaves. Planta, 1996, 199(2): 238-243. |