 2019, Vol. 39
2019, Vol. 39文章信息
- 康满春, 朱丽平, 许行, 查同刚, 张志强
- KANG Manchun, ZHU Liping, XU Hang, ZHA Tonggang, ZHANG Zhiqiang
- 基于Biome-BGC模型的北方杨树人工林碳水通量对气候变化的响应研究
- Modelling the responses of carbon and water fluxes with climate change for a poplar plantation in northern China based on the Biome-BGC model
- 生态学报. 2019, 39(7): 2378-2390
- Acta Ecologica Sinica. 2019, 39(7): 2378-2390
- http://dx.doi.org/10.5846/stxb201805291179
-
文章历史
- 收稿日期: 2018-05-29
- 网络出版日期: 2019-01-10





2. 三峡大学生物与制药学院, 宜昌 443002;
3. 北京林业大学水土保持与荒漠化防治教育部重点实验室, 北京 100083
2. College of Biological and Pharmaceutical Sciences, China Three Gorges University, Yichang 443002, China;
3. Key Laboratory of Soil and Water Conservation and Desertification Combating, Ministry of Education, College of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China
根据IPCC(AR5)报告, 气候变暖、CO2浓度上升等全球变化已是不争的事实[1]。在过去几十年中, 人工林由于能够快速地增加森林面积和森林碳储量, 其在应对温室气体浓度增加、气温上升等全球变化方面起着越来越重要的作用。在此背景下, 认识和理解人工林生态系统碳水通量的变化规律及其对气候变化的响应, 对于制定科学的人工林经营管理措施和合理应对气候变化具有十分重要的指导意义。
我国人工林栽植面积居世界首位, 其中杨树作为人工林主要树种之一, 由于生长迅速、适应性强且具有较高的生产力和固碳能力, 被广泛地用于碳汇造林、生物燃料、城市绿化和木材生产等方面, 造林面积截止2007年已达700多万hm2, 是中国北方地区的主要人工林类型[2]。但是杨树人工林在大量增加碳汇的同时也会消耗掉大量的水分和养分, 引起或加剧区域水资源匮乏、土地退化等问题, 尤其在中国北方缺水地区[3-6]。因此, 在未来气候变化条件下, 既发挥杨树人工林增加碳汇、调节气候等生态效益, 同时又不致影响到周边相邻生态系统及生境以至于增大环境退化的风险等[7-8]是森林经营主要挑战之一。因此, 需要更深入地认识杨树人工林生态系统碳水通量对气候变化的响应机理, 并量化气候变化对生态系统碳水通量的影响, 进而为制定科学合理的造林政策和经营管理杨树人工林提供理论依据和技术支持。
近年来, 很多基于生态生理过程的模型被广泛用于研究生态系统生态生理过程及其对气候变化的响应。其中Biome-BGC模型由于对植物生态生理过程的详细描述和具备更加完整的模拟时间尺度, 以及对植被在生态系统尺度上对气候变化响应的模拟, 已被广泛用于不同类型的森林生态系统碳、水通量的模拟研究[9-10], 如应用于亚马逊热带雨林[11]、热带干旱区落叶松林[12]、地中海森林[10]、云杉、松树、榉木、橡树、栎树、落叶松、樟子松等中欧温带森林[13-18]、东亚地区落叶松林[19]和北方森林等生态系统的模拟;国内的学者使用Biome-BGC模型分析研究了如阔叶红松林[20]、长白落叶松[21]、刺槐林[22]、油松林[23-24]、侧柏林、栓皮栎林[25]、华北落叶松林[26]和天山云杉[27]、湿地松[28]和马尾松[29]等森林生态系统生产力、蒸散发等的变化及其对增温、CO2浓度升高、氮沉降等气候变化和管理经营的响应。相比之下, 以往研究更多偏重于杨树人工林生态系统生态生理过程及其对环境因素的响应、固碳、蒸散发、以及碳水耦合等方面, 而对其碳水通量、水分利用效率如何响应气候变化等的研究则相对很少。
因此, 本文基于Biome-BGC模型, 运用生态系统通量观测中获取的气象数据和碳水通量数据驱动和优化模型, 根据当前气候变化的趋势模拟分析了不同气候变化情景下杨树人工林生态系统碳水通量及其水分利用效率, 主要目的在于:(1)分析杨树人工林碳水通量对气候变化的响应规律;(2)探究气候变化对杨树人工林水分利用效率的影响。
1 研究区概况本研究位于北京市大兴区榆垡镇大兴林场(39°31′50″N, 116°15′07″E), 试验样地总面积约为1.0 km2, 75%为2002年人工栽植的欧美107杨树(Populus euramericana cv. “74/76”)人工纯林。研究区属于暖温带亚湿润气候区, 多年均温在11.6℃, 多年内最高和最低温分别为40.6、-27.4℃;年均日照总时数为2772 h, 年均无霜期为209 d, 日均太阳辐射量约6.7 MJ m-2·d-1;风向变化显著, 主风向是东南风(夏季)和西北风(冬季);根据研究区附近气象站观测资料, 该区的降雨主要集中在7、8和9三个月份中, 该时期内的降雨量占全年降雨总量的60%—70%(根据1956—2000年大兴气象站观测数据, 116°15′07″E, 39°31′50″N), 过去20多年的年均降水量约为556 mm, 其中最大年降雨量达1085 mm, 最小年降雨量为262 mm。研究区内土壤为冲积性沙壤土, 平均土层厚度约为200 cm, 土质疏松, 具有通透性强而蓄水保肥能力差等特点;土壤pH值为8.25—8.39, 容重在1.43—1.47 g/cm3之间。
2 研究方法 2.1 Biome-BGC模型Biome-BGC模型是由美国蒙塔那大学陆地动态数值模拟小组NTSG(Numeric Terra Dynamic Simulation Group)所开发的生态过程机理模型, 模型基于生物区系的生物地球化学循环过程和水文过程受其所处气候环境和生物特性驱动的假设, 能够在日尺度上模拟和计算特定陆地生态系统类型中植被、凋落物以及土壤之间能量、碳、水和氮的储量和通量动态变化等[30]。
2.2 模型参数Biome-BGC模型的驱动参数包括站点参数(site parameters)、以日为步长的气象数据(meteorological data)和生态生理参数(eco-physiological parameters)。本研究中, 站点参数及气象数据(2006—2012年)分别来自实际调查测定和通量观测;另外, 采用碳氮分析仪(Vario MAX CN)和纤维素分析仪(Fibertec TM2010)采样测定了模型所需的杨树人工林部分生态生理参数, 如FRC:LC、C:Nleaf、C:Nfr、Llab、Lcel、Llig、FRlab、FRcel、FRlig、DWcel、DWlig;同时, 本研究基于Biome-BGC项目数据库和管理系统(Biome-BGC Project Database & Management System, 简称BBGCDB), 采用Monte Carlo技术和涡度相关观测数据, 对模型中所有的生态生理参数进行了敏感性分析, 并结合GLUE方法对碳水通量影响显著的13个参数进行优化。模型所需的站点及各生态生理参数的具体取值和获取方式见表 1。
| 参数 Parameters |
符号 Symbols |
取值 Values |
单位 Units |
来源 Data source |
| 站点特性参数Site-specific parameters | ||||
| 有效土壤深度Effective soil depth | — | 2.0 | m | 本研究测定 |
| 海拔Elevation | — | 30 | m | 本研究测定 |
| 纬度Latitude | — | 39.53 | degree | 本研究测定 |
| 经度Longitude | — | 116.25 | degree | 本研究测定 |
| 地表反射率Shorewave albedo | albedo | 0.20 | — | 本研究测定 |
| 土壤机械组成SoilS Texture 砂粒含量Sand | — | 94.96 | % | 本研究测定 |
| 粉粒含量Silt | — | 2.49 | % | 本研究测定 |
| 黏粒含量Clay | — | 2.55 | % | 本研究测定 |
| 大气干湿氮沉降Wet+dry atmospheric deposition of N | — | 0.006 | kg N m-2 a-1 | [31] |
| 生物固氮Symbiotic+asymbiotic fixation of N | — | 0.000337 | kg N m-2 a-1 | |
| 生理生态参数Eco-physiological parameters | ||||
| 木本或非木本植被类型Woody/non-woody flag | — | 1 | Flag | 设定 |
| 常绿或落叶Evergreen/deciduous flag | — | 0 | Flag | 设定 |
| 光合类型C3/C4 | — | 1 | Flag | 设定 |
| 物候模拟或自定义Model phenology/user specified | — | 0 | Flag | 设定 |
| 生长季开始Yearday to start new growth | ONDAY | 101 | Flag | 本研究测定 |
| 生长季结束Yearday to end litterfall | OFFDAY | 302 | Flag | 本研究测定 |
| 转换生长占生长季比例Transfer growth period as fraction of growing season | TFG | 0.204 | Prop. | 参数优化 |
| 落叶时段占生长季比例Litterfall as fraction of growing season | LFG | 0.205 | Prop. | 参数优化 |
| 叶片和细根年周转率Annual leaf and fine root turnover fraction | LFRT | 1 | 1 a-1 | 默认参数 |
| 活立木年周转率Annual live wood turnover fraction | LWT | 0.578 | 1 a-1 | 参数优化 |
| 整株植物死亡率Annual whole-plant mortality fraction | WPM | 0.006 | 1 a-1 | 参数优化 |
| 植物火烧死亡率Annual fire mortality fraction | FM | 0 | 1 a-1 | 本研究测定 |
| 细根与叶片碳分配比New fine root C:new leaf C | FRC:LC | 1.04 | kgC/kgC | 本研究测定 |
| 茎与叶片碳分配比New stem C:new leaf C | SC:LC | 3.1 | kgC/kgC | 参数优化 |
| 活木与木质组织碳分配比New live wood C:new total wood C | LWC:TWC | 0.228 | kgC/kgC | 参数优化 |
| 粗根与茎分配比New croot C:new stem C | CRC:SC | 0.22 | kgC/kgC | 默认参数 |
| 当前生长比例Current growth proportion | CGP | 0.437 | Prop. | 参数优化 |
| 叶片碳氮比C:N of leaves | C:Nleaf | 16.58 | kgC/kgN | 本研究测定 |
| 凋落物碳氮比C:N of leaf litter, after retranslocation | C:Nlit | 49 | kgC/kgN | 默认参数 |
| 细根碳氮比C:N of fine roots | C:Nfr | 43.2 | kgC/kgN | 本研究测定 |
| 活木质组织碳氮比C:N of live wood | C:Nlw | 50 | kgC/kgN | 默认参数 |
| 死木质组织碳氮比C:N of dead wood | C:Ndw | 550 | kgC/kgN | 默认参数 |
| 凋落物中易分解物质比例Leaf litter labile proportion | Llab | 0.20 | % | 本研究测定 |
| 凋落物中纤维素比例Leaf litter cellulose proportion | Lcel | 0.51 | % | 本研究测定 |
| 凋落物中木质素比例Leaf litter lignin proportion | Llig | 0.29 | % | 本研究测定 |
| 细根中易分解物质比例Fine root labile proportion | FRlab | 0.12 | % | 本研究测定 |
| 细根纤维素比例Fine root cellulose proportion | FRcel | 0.54 | % | 本研究测定 |
| 细根中木质素比例Fine root ligin proportion | FRlig | 0.34 | % | 本研究测定 |
| 死木质组织中的纤维素比例Dead wood cellulose proportion | DWcel | 0.78 | % | 本研究测定 |
| 死木质组织中的木质素比例Dead wood lignin proportion | DWcel | 0.22 | % | 本研究测定 |
| 冠层截留系数Canopy water interception coefficient | wint | 0.0355 | 1 LAI-1 d-1 | 参数优化 |
| 冠层消光系数Canopy light extinction coefficient | k | 0.599 | — | 参数优化 |
| 叶面积与投影叶面积指数比All-sided to projected leaf arera ratio | LAIall:proj | 2 | LAI/LAI | 默认参数 |
| 冠层比叶面积Canopy average specific leaf area (projected area basis) | SLA | 46.64 | m2/kgC | 参数优化 |
| 阴生叶和阳生叶的比叶面积比例Ratio of shaded SLA:sunlit SLA | SLAshd:sun | 2 | SLA/SLA | 默认参数 |
| 酮糖二磷酸羧化酶中氮含量与叶氮含量Fraction of leaf N in Rubisco | FLNR | 0.157 | — | 参数优化 |
| 最大气孔导度 Maximum stomatal conductance (projected area basis) |
gsmax | 0.0094 | m/s | 参数优化 |
| 表皮层导度Cuticular conductance (projected area basis) | gcut | 0.000094 | m/s | 参数优化 |
| 边界层导度Boundary layer conductance (projected area basis) | gbl | 0.01 | m/s | 默认参数 |
| 气孔开始缩小时的叶片水势 Leaf water potential:start of conductance reduction |
LWPi | -0.34 | Mpa | [9] |
| 气孔完全闭合时的叶片水势 Leaf water potential:complete conductance reduction |
LWPf | -2.2 | Mpa | [9] |
| 气孔开始缩小时的饱和水汽压差 Vapor pressure deficit:start of conductance reduction |
VPDi | 1100 | Pa | [9] |
| 气孔完全闭合时的饱和水汽压差 vapor pressure deficit:complete conductance reduction |
VPDf | 3600 | Pa | [9] |
| 表中默认参数来自[32] | ||||
本研究中, 通过涡度相关系统观测所得的碳水通量用于模型验证, 系统仪器的布设和数据质量控制见于之前的相关研究[7-8]。
2.3.2 模型表现评价参数本研究中, 通过对碳水通量的模拟值与实测值的比较, 选取3个统计变量来评价模型的模拟效果, 分别为决定系数(R2)、Nash-Sutcliffe效率系数(NS)和均方根误差(RMSE), 各自的计算公式如下:
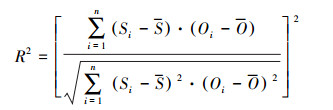
|
(1) |
该值可以通过模型模拟值与观测值之间的线性回归所得, 反映模拟值吻合观测值变异的能力, 越接近1表示模拟结果与实测结果的拟合程度越高。
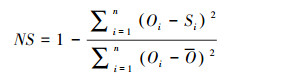
|
(2) |
式中, NS的取值范围从负无穷大(表示模拟效果差)到1(模拟的效果好)。反映模拟值和观测值的吻合程度, 如果模拟与观测之间的方差和观测方差一样大, 则NS=0;如果模拟与观测之间的方差大于观测方差, 则NS < 0;如果模拟与观测之间的方差趋于0, 则NS趋近于1, 表示模型很好地模拟了观测值的变化。

|
(3) |
用于评价模拟值和实测值之间的偏差即模拟结果的准确性, RMSE越小, 表示模拟值与观测值之间的偏差越小, 模型的模拟表现越好。
上述各式中Si和Oi分别代表模拟值和实测值, 而S和O分别为模拟值和观测值的均值, n代表观测的次数。
2.3.3 水分利用效率(WUE)本研究中生态系统水分利用效率(WUE)的计算公式如下:
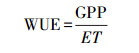
|
(4) |
式中, GPP为生态系统总初级生产力, 单位g C m-2 a-1;ET为生态系统年总蒸散发, 单位mm/a。
2.4 气候变化情景设定IPCC(AR5)根据本世纪末辐射强迫水平的不同, 通过“典型浓度路径(RCP)”描述了4种不同路径下21世纪温室气体排放情景, 分别是RCP2.6、RCP4.5、RCP6.0和RCP8.5, 如其在RCP4.5情景下是580—720 ppm, 而在RCP6.0情景下, 到21世纪中期CO2排放是580—720 ppm, 末期为720—1000 ppm。在4种不同的情景中, 其中RCP4.5情景由于考虑了社会经济和政策对未来气候的影响, 比较符合未来中国的发展情况[33];而RCP6.0相当于AR4中的SRES A1B情景, 所以在关于未来气候变化的模拟研究中被经常采用。
但是, 由于气候具有明显的区域性特征, 所以在研究气候变化对区域环境、生态系统的影响时, 需要在大的全球气候背景下更加关注区域气候及其变化的更为直接的影响[34]。对于中国地区未来气候变化的预估分析一致认为, 未来的气温将升高, 降水区域平均呈增加趋势, 但存在较大的区域性差异[35]。当前, 根据基于区域气候模式在不同排放情景下对中国华北地区未来气候变化的模拟结果, 该地区在21世纪的中期的气温较过去几十年上升1.8—2.2℃、2.3—2.5℃或3—3.9℃, 而全年降水变化为±5%或者增加5%—10%间, 而在21世纪末期, 气温和降水的变化分别是上升3.5—4.0℃或者4.5—5.8℃和增加10%—15%[34-35]。根据以上的分析, 本研究将基于RCP4.5和RCP6.0中的CO2排放情景, 结合华北地区气候变化中的气温和降水的变化情景, 采用的气候变化情景组合如表 2中所示。
| 序号 No. |
气候变化情景 Cliamte change scenarios |
CO2浓度(C)
Atmospheric CO2 concentration |
气温(T)
Air temperature |
降水(P)
Precipitation |
| 1 | C0T0P0 | 不变 | 不变 | 不变 |
| 2 | C1T0P0 | 650 ppm | 不变 | 不变 |
| 3 | C2T0P0 | 860 ppm | 不变 | 不变 |
| 4 | C0T1P0 | 不变 | +2℃ | 不变 |
| 5 | C0T2P0 | 不变 | +4℃ | 不变 |
| 6 | C0T3P0 | 不变 | +6℃ | 不变 |
| 7 | C0T0P1 | 不变 | 不变 | -5% |
| 8 | C0T0P2 | 不变 | 不变 | +5% |
| 9 | C0T0P3 | 不变 | 不变 | +10% |
| 10 | C0T0P4 | 不变 | 不变 | +15% |
| 11 | C1T1P1 | 650 ppm | +2℃ | -5% |
| 12 | C1T1P2 | 650 ppm | +2℃ | +5% |
| 13 | C1T1P3 | 650 ppm | +2℃ | +10% |
| 14 | C1T2P1 | 650 ppm | +4℃ | -5% |
| 15 | C1T2P2 | 650 ppm | +4℃ | +5% |
| 16 | C1T2P3 | 650 ppm | +4℃ | +10% |
| 17 | C1T2P4 | 650 ppm | +4℃ | +15% |
| 18 | C2T2P1 | 860 ppm | +4℃ | -5% |
| 19 | C2T2P2 | 860 ppm | +4℃ | +5% |
| 20 | C2T2P3 | 860 ppm | +4℃ | +10% |
| 21 | C2T2P4 | 860 ppm | +4℃ | +15% |
| 22 | C2T3P3 | 860 ppm | +6℃ | +10% |
| 23 | C2T3P4 | 860 ppm | +6℃ | +15% |
基于观测所得杨树人工林生态系统2006—2007年总初级生产力(GPP)和生态系统蒸散发(ET), 对比分析了Biome-BGC模型在默认参数和优化参数后两种情形下对杨树人工林GPP、ET的模拟效果, 如表 3所示。从表 3中可以看出, Biome-BGC模型对杨树人工林GPP、ET的模拟表现为, GPP在默认参数和优化参数情形下实测值和模拟值的拟合斜率分别为1.24和0.88, 说明在两种情形下各自存在高估和低估现象;GPP的模拟在两种情形下的模拟拟合程度均较好(决定系数R2约为0.75)。相比之下, 参数优化后Biome-BGC对杨树人工林GPP模拟表现为RMSE从2.64 g C m-2·d-1减小为1.94 g C m-2·d-1, 降低幅度为26.5%, NS从0.42上升至0.69(更趋近于1), 其增幅达64.3%, 表明校准后的Biome-BGC模型对GPP变化模拟的吻合程度及其准确性均有显著提升。对于ET的模拟, 两种情形下都存在高估现象, 模型模拟值能够解释实测值70%—80%的变化, 模型模拟值的拟合程度相比于GPP的其表现较好;同时, 参数优化后模型对ET变化模拟的吻合程度有所提升, 表现为RMSE从1.18 mm/d降低为0.88 mm/d, 降低幅度达25.4%, 而对其模拟准确性上则有显著提升, NS从0.35提升至0.63(更趋近于1), 其上升幅度为80%。
| 参数组合情形 Parameter combination |
碳水通量 Carbon and water flux |
回归方程 Regression equation |
决定系数(R2)
Determination coefficient |
效率系数 (NS Nash-Sutcliffe coefficient |
均方根 误差(RMSE) Root mean square error |
显著性水平 (P) Significance level |
| 默认参数 | GPP | y = 1.2397x - 0.4462 | 0.75 | 0.42 | 2.64 g C m-2·d-1 | < 0.001 |
| Default parameters | ET | y = 1.1931x - 0.1893 | 0.70 | 0.35 | 1.18 mm/d | < 0.001 |
| 优化参数 | GPP | y = 0.8769x - 0.2155 | 0.75 | 0.69 | 1.94 g C m-2·d-1 | < 0.001 |
| Modified parameters | ET | y=1.1619x - 0.2036 | 0.80 | 0.63 | 0.88 mm/d | < 0.001 |
| GPP:总初级生产力, gross primary productivity;ET:蒸散发, evapotranspiration;y:模型模拟值, simulated value;x:站点实测值, observed value | ||||||
为了进一步检验模型的校准效果是否可信, 利用2008—2009年GPP、ET对校准后Biome-BGC模型进行验证, 结果如图 1所示, 整体上, 参数优化后的Biome-BGC对GPP季节变化的模拟与实测值具有较高的一致性, 相比于校准期的决定系数R2、NS和RMSE, 验证期模拟结果的这些参数分别为0.87、0.85和1.60 g C m-2 d-1, 而对ET的模拟则有所高估, 其在验证期的R2、NS和RMSE分别为0.79、0.71和0.87 mm/d, 其较低的R2可能与Biome-BGC模型对土壤水平衡过程、冠层水分截留等过程的简化有关。总体上, 验证期R2、NS和RMSE基本与校准期一致, 表明校准后的Biome-BGC模型对杨树人工林GPP、ET的模拟效果较好, 且具有一致性和可信性, 因而可用于杨树人工林生态系统的模拟。

|
| 图 1 验证期(2008—2009年)杨树人工林总初级生产力(GPP)、蒸散发(ET)日尺度实测值与模拟值对比图 Fig. 1 Linear fitting between simulated and observed daily gross primary productivity (GPP), evapotranspiration (ET) during validation period (2008—2009) in poplar plantation |
在不同气候变化情景下, 杨树人工林生态系统总初级生产力(GPP)对气温上升响应变化如图 2所示。在当前大气CO2浓度及降水不变的情况(对应C0P0)下, 单独气温的上升会降低对杨树人工林生态系统GPP, 且气温升高幅度越大, GPP降低幅度越大, 其中在气温升高6℃时GPP降低幅度为5%;在其他气候变化情景中, GPP相比于C0T0P0情景下的均有所提升, 但气温上升仍会导致GPP上升幅度减弱, 如C1T1P1与C1T2P1, C1T1P2和C2T2P2等情景中。这表明无论在当前气候条件或者在未来气候变化情景下, 气温的上升会抑制杨树人工林的GPP增加幅度。

|
| 图 2 不同气候变化情景下的总初级生产力(GPP)及其各自相对于C0T0P0情景下GPP的变化幅度 Fig. 2 Gross primary productivity (GPP) under climate change scenarios and their relative variation to GPP of C0T0P0 图中横坐标C0T0P0—C2T3P4代表由C:CO2浓度Atmospheric CO2 concentration;T:气温Air temperature;P:降水Precipitation; 三个因子组合形成的23种气候变化情景, 其中数字0—4分别代表各因子的不同变化水平, 不同情景各因子变化水平详见表 1 |
本研究对未来CO2浓度上升共设置了650 ppm和860 ppm两种情景, 当CO2浓度达到650 ppm和860 ppm时杨树人工林的GPP分别较C0T0P0情景下的上升了28%和44%;其中在C2T2P4情景下, GPP提高的程度最大, 达到53%;在其他气温上升、降水变化情景相同的条件下, 如在T2P1、T2P2、T2P3等背景下, CO2浓度上升会显著地增加生态系统GPP, 且随着降水的增加, 其促进GPP增长的作用也越大, 如在T2P3情景中, CO2浓度达到650 ppm和860 ppm时所对应的GPP的分别提高了34%和50%, 高于T2P2和T2P1情景下CO2对应浓度时其对GPP的促进作用。
3.2.3 GPP对降水变化的响应对未来降水变化情景共设置了4个梯度, 分别为比当前年总降水量减少5%, 增加5%、10%和15%的变化情景, 由模拟结果(图 2)可知, 在当前大气CO2浓度和气温条件下, 降水减少5%, 杨树人工林生态系统的GPP下降了4%, 随着降水增加5%、10%和15%, GPP则分别增加了3%、7%和10%;在未来CO2浓度上升且气温升高的气候情景下(C1T1、C1T2、C2T2以及C3T3), 随着降水的增加, GPP的增加幅度也逐渐变大。
3.3 杨树人工林水通量对气候变化的响应模拟 3.3.1 Es对气候变化的响应土壤蒸发(Es)在不同气候条件下的模拟结果及其相对C0T0P0情景的变化如图 3中所示, 相对于当前气候条件下的Es, 不同气候变化情景中杨树人工林生态系统的Es对单独气温上升、大气CO2浓度升高和降水增加的响应分别表现为下降、上升和上升, 且气候变化的幅度越大, 造成的Es变化幅度也越大;其中气温上升和降水减少将导致Es的下降, 气温上升会显著的抑制Es, 且气温上升的幅度越大, Es下降的越大, 在气温上升6℃时导致Es下降幅度为11%;大气CO2浓度上升和降水增加使Es增大, 增加的最大幅度分别为18%和10%。在组合的气候变化情景中, 气温上升对Es的降低在一定程度上抵消了CO2浓度和降水增加对Es的增强作用;整体上, 在C2T0P0和C2T2P4气候变化情景中Es上升幅度最大, 而在高温和少雨的C0T3P0情景中Es降低幅度最大(11%)。

|
| 图 3 不同气候变化情景下的蒸散发(ET)、蒸腾(Tr)和土壤蒸发(Es)量及各自相对于C0T0P0情景下模拟值的变化幅度 Fig. 3 Evapotranspiration (ET), tranpiration (Tr) and soil evaporation (Es) under climate change scenarios and their relative variation to that of C0T0P0 ET:蒸散发, Evapotranspiration;Tr:蒸腾, transpiration;Es:土壤蒸发, soil evaporation |
根据图 3所示, 相比于当前气候条件(C0T0P0), 不同气候变化情景中气温上升和降水增加均使杨树人工林的Tr增加, 而CO2浓度上升和降水减少则会抑制Tr;其中单独气温上升引起的Tr的增加幅度在1%—4%, 降水增加促使Tr增加的幅度在5%—16%, 且Tr升高比例与降水增加的比例一致, 而CO2浓度上升和降水减少导致Tr的降低幅度分别为4%—7%和5%;各气候变化情景中气温上升、降水变化以及CO2浓度上升的作用通过相互叠加从而影响Tr的变化, 其中以C0T0P4情景中Tr较当前气候条件下的上升幅度最大(16%), 而C1T1P1情景中的Tr则较当前气候条件下的下降幅度最大(8%)。
3.3.3 ET对气候变化的响应Biome-BGC模型所模拟的生态系统蒸散发(ET)是土壤蒸发(Es)和植物蒸腾(Tr)之和, Tr约占ET的72%, ET对未来气候情景的响应分别是Es和Tr对各气候变化情景响应结果的总和。根据图 3, 各气候变化情景中, 在大气CO2浓度、降水条件相同时, 气温上升对ET没有显著影响;而在相同的气温和降水条件下, ET并不随CO2浓度的上升而增加;这是由于Es对气温升高和大气CO2浓度上升的响应分别为减弱和上升, 而Tr对两种因素的响应则分别为上升和降低, 且Es的响应幅度要大于Tr的, 所以两者之和ET对气温升高和CO2浓度增大的响应不显著。
ET对降水变化的响应则显著不同于对气温和CO2浓度变化的响应, ET在气温和大气CO2浓度一致的气候变化情景中, 如在C0T0、C1T1、C1T2、C2T2和C2T3等情景, 由于Es和Tr都表现出随着降水的增加而上升的趋势, 所以杨树人工林生态系统的ET表现出随着降水的增加呈逐渐上升的趋势;由于ET只对降水的变化有显著响应, 因而在不同气候变化情景中只要降水多, 该气候情景下的ET也相应地上升, 如在降水增加15%的C0T0P4、C1T2P4、C2T2P4和C2T3P4这些气候变化情景中, 对应的ET比当前气候条件下的增加了14%。
3.4 气候变化下生态系统水分利用效率(WUE)的响应基于Biome-BGC模型对GPP和ET模拟结果, 分别估算了不同气候变化情景下杨树人工林WUE及其相对于当前的变化, 结果如图 4所示。杨树人工林WUE在当前气候条件下(C0T0P0)为2.8 g C /kg H2O, 在所模拟的气候变化情景中, WUE最大值和最小值分别出现在C2T2P1(4.0g C /kg H2O)和C0T3P0(2.6 g C /kg H2O)情景, 相对于当前气候下WUE分别上升了45.3%和下降了5.8%;WUE对气候变化各因素的响应总体上表现为:WUE随着气温的上升和降水的增加呈小幅下降, 随降水的减少呈小幅上升, 随着大气CO2浓度的上升而大幅提高;其中, 在各气候变化情景中, 当大气CO2浓度达到650 ppm(C1)时, 其对应的WUE高于C0T0P0情景下的20%以上, 而当大气CO2浓度达到860 ppm(C2)时, 各情景下WUE均超过C0T0P0的33%以上。

|
| 图 4 不同气候变化情景下的水分利用效率(WUE)及其各自相对于C0T0P0情景下WUE的变化幅度(%) Fig. 4 Water use efficiency (WUE) under climate change scenarios and their relative variation (%) to WUE of C0T0P0 |
气温上升对生态系统生产力的影响是双面的, 一方面可以延长植物的生长季, 继而提高光合作用效率和植物的生产力[36], 如研究表明气温升高提高了中国北方地区森林的生态系统生产力[37];另一方面则增加水分消耗而引起干旱胁迫, 反而降低生态系统的生产力[36]。在干旱或半干旱地区, 当土壤水分成为胁迫因子时, 是影响土壤蒸发的主要因素。另外, 由于气温上升会导致叶子内部水汽压的急剧增大, 其与稳定大气水汽压之间的饱和水汽压差会大大提高, 从而显著地提高植物的蒸腾作用[38]。本研究中杨树人工林GPP随气温上升反而降低, 这可能是由于其所在地沙土的保水能力差, 导致杨树人工林无论在干旱还是湿润年份均处于水分短缺情况[39], 增温则会进一步加剧干旱胁迫, 从而降低其GPP。相应地, 增温加剧干旱胁迫导致Es的较大幅度下降;Tr随气温的上升而增加, 但处于干旱胁迫状态下的杨树人工林为了减少失水会降低气孔导度甚至关闭气孔, 所以其Tr随着气温上升只有小幅度的增加。因此, 导致杨树人工林ET(Es和Tr之和)由于增温对Es和Tr相反的影响作用而无显著变化。
4.2 杨树人工林碳水通量对降水变化的响应在干旱及半干旱地区, 降水以及由此引起的土壤水分动态变化是植被演变的主要限制性因子[23]。水分胁迫会导致植物叶片的气孔阻力增大甚至关闭气孔, 从而使植物的蒸腾和光合作用都显著降低[40]。未来气候变化中, 降水的增加会改善中国北方半干旱地区的水分短缺状况, 提高土壤中的可利用水分, 减缓该地区杨树人工林所受的干旱胁迫, 在一定程度上能够促使增温发挥其增加GPP的作用, 从而使杨树人工林的GPP比当前气候下的有所增加;同时, Es和Tr由于降水增加改善了土壤水分条件而上升, 使得杨树人工林生态系统ET也随着降水增加呈上升趋势。
4.3 杨树人工林碳水通量对CO2浓度上升的响应一般认为, CO2浓度上升对植物起着“施肥”作用, 能够在短期内促进森林生产力和生物量的增加。目前, 几乎所有探究关于CO2浓度上升如何影响植物的实验都是在人工气室中进行的, 对CO2的“施肥效应”是如何影响生长在野外的树木以及生态系统还缺乏直接证据。大部分的人工控制环境下的模拟实验结果表明:CO2浓度升高能够在一定程度上促进光合速率, 从而促进植物的生长[23], 尤其对C3类植物的促进作用可能更大。但是CO2浓度上升的影响因植物品种、光合作用类型、生长发育期等的不同而有所不同[41], 如有很多物种对CO2浓度上升没有反应, 而且长期的熏蒸也往往导致光合驯化的发生[42];本研究中, 无论在CO2浓度上升单独作用下, 还是在其与气温上升和降雨增加两个因素的协同影响下, 杨树人工林GPP都表现为上升, 这可能是因为CO2既可以作为植物光合作用的原料, 也可以通过对温度、植物水分需求及植物营养物质需求等方面的作用来影响植物生长[40];也有研究表明, CO2浓度升高能够提高土壤氮的可获得性, 从而提高了植物的光合效率[23]。
另外, CO2浓度升高将引起植物叶片及冠层温度的上升以及气孔阻力的增大, 导致蒸腾速率的下降[43], 这可能是导致杨树人工林Tr随CO2浓度上升而下降的原因。同时, CO2浓度升高所引起的气温的上升会增加土壤水分的蒸发, 导致土壤水分含量下降等, 引起植物的“生理干旱”, 进而限制植物的光合作用和生长速度[44]。
4.4 杨树人工林水分利用效率(WUE)对气候变化的响应WUE是量化生态系统碳水循环耦合关系的重要参数[45]。研究中国北方缺水地区杨树人工林WUE对气候变化的响应, 有助于制定科学的经营管理措施以应对区域气候变化。本研究中, 不同气候变化情景下杨树人工林WUE的变化范围为2.6—4.0 g C /kg H2O, 其中当前气候条件下WUE的估算结果为2.8 g C /kg H2O, 与在干旱条件下基于实测值的计算结果一致[46]。WUE随气温上升而降低, 已有研究认为主要原因是气温上升导致干旱胁迫进而造成植物生长受阻且同时使ET增加[47], 但本研究中ET并没有随气温上升而增加。WUE随降水增加而降低, 随降水减少而升高, 主要是因为ET对降水变化的响应大于GPP, 这可能与本研究中杨树人工林即使在设定的最大降水增加幅度下仍处于干旱胁迫状态有关[39];WUE随大气CO2浓度上升而提高, 且其对WUE的影响远大于气温和降水变化的, 这与Pan[47]、Keenan[48]等人的研究一致, 其可能原因被认为是CO2浓度上升通过“施肥效应”促进了植物光合, 又通过降低叶片气孔导度进而减弱ET[48], 在本研究中则主要是由于CO2浓度上升导致GPP的增加幅度远高于ET的。
5 结论基于参数优化和通量数据校准的Biome-BGC模型有效提升了其在模拟杨树人工林GPP和ET的吻合程度和准确性, 能够有效应用于杨树人工林对气候变化响应的研究。在设定的23种气候变化情景中, 杨树人工林碳水通量对气候变化的响应主要受气温上升、降水变化和大气CO2浓度上升三者复合作用的影响, GPP、Es、Tr和ET对不同因素的响应表现为:对气温上升的响应分别为降低、减少、升高和不变, 随降水的增加和减少均呈上升和下降趋势, 而随着大气CO2浓度上升则分别表现为升高、上升、下降和不变;其中大气CO2浓度上升对GPP和Es的影响远高于气温上升和降水变化的, 降水变化则是影响Tr和ET的主要因素。基于GPP和ET对气候变化的响应, WUE在未来气候变化情景中的变化范围在2.6—4.0 g C /kg H2O之间, 其对气温上升、降水增加和大气CO2浓度升高的响应则分别表现为降低、降低和升高, 且未来气候变化中, 其对大气CO2浓度升高的响应程度远高于对气温上升和降水变化两者的, 表明未来气候变化中大气CO2浓度上升是影响杨树生长的主要因素。
| [1] |
沈永平, 王国亚. IPCC第一工作组第五次评估报告对全球气候变化认知的最新科学要点. 冰川冻土, 2013, 35(5): 1068-1076. |
| [2] |
方升佐. 中国杨树人工林培育技术研究进展. 应用生态学报, 2008, 19(10): 2308-2316. |
| [3] |
Migliavacca M, Meroni M, Manca G, Matteucci G, Montagnani L, Grassi G, Zenone T, Teobaldelli M, Goded I, Colombo R, Seufert G. Seasonal and interannual patterns of carbon and water fluxes of a poplar plantation under peculiar eco-climatic conditions. Agricultural and Forest Meteorology, 2009, 149(9): 1460-1476. DOI:10.1016/j.agrformet.2009.04.003 |
| [4] |
Kim H S, Oren R, Hinckley T M. Actual and potential transpiration and carbon assimilation in an irrigated poplar plantation. Tree Physiology, 2008, 28(4): 559-577. DOI:10.1093/treephys/28.4.559 |
| [5] |
Li Y Z, Qin H Y, Xie Y H, Wang W, Chen X S, Zhang C M. Physiological mechanism for the reduction in soil water in poplar (Populus deltoides) plantations in Dongting Lake wetlands. Wetlands Ecology and Management, 2014, 22(1): 25-33. |
| [6] |
Stanturf J A, van Oosten C. Operational poplar and willow culture//Isebrands J G, Richardson J, eds. Poplars and Willows: Trees for Society and the Environment. Oxfordshire, England: CABI, 2012: 200-257.
|
| [7] |
康满春, 蔡永茂, 王小平, 查同刚, 朱丽平, 牛勇, 周洁, 张志强.表层阻力和环境因素对杨树(Populus sp.)人工林蒸散发的控制.生态学报, 2016, 36(17): 5508-5518.
|
| [8] |
Zhou J, Zhang Z Q, Sun G, Fang X R, Zha T G, McNulty S, Chen J Q, Jin Y, Noormets A. Response of ecosystem carbon fluxes to drought events in a poplar plantation in Northern China. Forest Ecology and Management, 2013, 300: 33-42. DOI:10.1016/j.foreco.2013.01.007 |
| [9] |
Migliavacca M, Meroni, M, Busetto L, Colombo R, Zenone T, Matteucci G, Manca G, Seufert G. Modeling gross primary production of agro-forestry ecosystems by assimilation of satellite-derived information in a process-based model. Sensors, 2009, 9(2): 922-942. DOI:10.3390/s90200922 |
| [10] |
Chiesi M, Chirici G, Corona P, Duce P, Salvati R, Spano D, Vaccari F P, Maselli F. Use of BIOME-BGC to simulate water and carbon fluxes within Mediterranean macchia. iForest-Biogeosciences and Forestry, 2012, 5(5): 38-43. |
| [11] |
Hashimoto H, Melton F, Ichii K, Milesi C, Wang W L, Nemani R R. Evaluating the impacts of climate and elevated carbon dioxide on tropical rainforests of the western Amazon basin using ecosystem models and satellite data. Global Change Biology, 2010, 16(1): 255-271. DOI:10.1111/(ISSN)1365-2486 |
| [12] |
Dai Z, Johnson K D, Birdsey R A, Hernandez-Stefanoni J L, Dupuy J M. Assessing the effect of climate change on carbon sequestration in a Mexican dry forest in the Yucatan Peninsula. Ecological Complexity, 2015, 24: 46-56. DOI:10.1016/j.ecocom.2015.09.004 |
| [13] |
Churkina G, Tenhunen J, Thornton P, Falge E M, Elbers J A, Erhard M, Grünwald T, Kowalski A S, Rannik V, Sprinz D. Analyzing the ecosystem carbon dynamics of four european coniferous forests using a biogeochemistry model. Ecosystems, 2003, 6(2): 168-184. DOI:10.1007/s10021-002-0197-2 |
| [14] |
Eastaugh C S, Pötzelsberger E, Hasenauer H. Assessing the impacts of climate change and nitrogen deposition on Norway spruce (Picea abies L. Karst) growth in Austria with BIOME-BGC. Tree Physiology, 2011, 31(3): 262-274.
|
| [15] |
Hlásny T, Barcza Z, Fabrika M, Balázs B, Churkina G, Pajtík J, Sedmák R, Tuřáni M. Climate change impacts on growth and carbon balance of forests in Central Europe. Climate Research, 2011, 47(3): 219-236. DOI:10.3354/cr01024 |
| [16] |
Jochheim H, Puhlmann M, Beese F, Berthold D, Einert P, Kallweit R, Konopatzky A, Meesenburg H, Meiwes K J, Raspe S, Schulte-Bisping H, Schulz C. Modelling the carbon budget of intensive forest monitoring sites in Germany using the simulation model BIOME-BGC. iForest-Biogeosciences and Forestry, 2009, 2: 7-10. DOI:10.3832/ifor0475-002 |
| [17] |
Merganicová K, Merganic J, Hasenauer H. Assessing the carbon flux dynamics within virgin forests:the case study 'Babia hora' in Slovakia. Austrian Journal of Forest Science, 2012, 129(1): 1-21. |
| [18] |
Tatarinov F A, Cienciala E, Vopenka P, Avilov V. Effect of climate change and nitrogen deposition on central-European forests:Regional-scale simulation for South Bohemia. Forest Ecology and Management, 2011, 262(10): 1919-1927. DOI:10.1016/j.foreco.2011.02.020 |
| [19] |
Ueyama M, Ichii K, Hirata R, Takagi K, Asanuma J, Machimura T, Nakai Y, Ohta T, Saigusa N, Takahashi Y, Hirano T. Simulating carbon and water cycles of larch forests in East Asia by the BIOME-BGC model with AsiaFlux data. Biogeosciences, 2010, 7(3): 959-977. DOI:10.5194/bg-7-959-2010 |
| [20] |
吴玉莲, 王襄平, 李巧燕, 孙阎. 长白山阔叶红松林净初级生产力对气候变化的响应:基于BIOME-BGC模型的分析. 北京大学学报:自然科学版, 2014, 50(3): 577-586. |
| [21] |
何丽鸿, 王海燕, 王璐, 王岳. 长白落叶松林生态系统净初级生产力对气候变化的响应. 北京林业大学学报, 2015, 37(9): 28-36. |
| [22] |
张艺, 余新晓, 范敏锐, 常存, 陆晓宇. 北京山区刺槐林净初级生产力对气候变化的响应. 水土保持研究, 2012, 19(3): 151-155. |
| [23] |
彭俊杰, 何兴元, 陈振举, 崔明星, 张先亮, 周长虹. 华北地区油松林生态系统对气候变化和CO2浓度升高的响应——基于BIOME-BGC模型和树木年轮的模拟. 应用生态学报, 2012, 23(7): 1733-1742. |
| [24] |
张文海, 吕锡芝, 余新晓, 范敏锐. 气候和CO2变化对北京山区油松林NPP的影响. 广东农业科学, 2012(6): 4-7. DOI:10.3969/j.issn.1004-874X.2012.06.002 |
| [25] |
范敏锐, 余新晓, 张振明, 史宇, 吕锡芝, 周彬. CO2倍增和气候变化对北京山区栓皮栎林NPP影响研究. 生态环境学报, 2010, 19(6): 1278-1283. DOI:10.3969/j.issn.1674-5906.2010.06.003 |
| [26] |
苏薇, 余新晓, 吕锡芝, 范敏锐, 张艺. 气候变化对北京山区华北落叶松林NPP影响研究. 广东农业科学, 2012(7): 69-72. DOI:10.3969/j.issn.1004-874X.2012.07.026 |
| [27] |
苏宏新.全球气候变化条件下新疆天山云杉林生长的分析与模拟[D].北京: 中国科学院研究生院(植物研究所), 2005.
|
| [28] |
Ma Z Q, Liu Q J, Wang H M, Li X R, Zeng H Q, Xu W J. Observation and modeling of NPP for Pinus elliottii plantation in subtropical China. Science in China Series D:Earth Sciences, 2008, 51(7): 955-965. DOI:10.1007/s11430-008-0075-7 |
| [29] |
Zhang T L, Sun R, Peng C H, Zhou G Y, Wang C L, Zhu Q A, Yang Y Z. Integrating a model with remote sensing observations by a data assimilation approach to improve the model simulation accuracy of carbon flux and evapotranspiration at two flux sites. Science China Earth Sciences, 2016, 59(2): 337-348. DOI:10.1007/s11430-015-5160-0 |
| [30] |
曾慧卿, 刘琪璟, 冯宗炜, 王效科, 马泽清. 基于BIOME-BGC模型的红壤丘陵区湿地松(Pinus elliottii)人工林GPP和NPP. 生态学报, 2008, 28(11): 5314-5321. DOI:10.3321/j.issn:1000-0933.2008.11.013 |
| [31] |
韩玉国, 李叙勇, 南哲, 李波. 北京地区2003-2007年人类活动氮累积状况研究. 环境科学, 2011, 32(6): 1537-1545. |
| [32] |
White M A, Thornton P E, Running S W, Nemani R R. Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model:net primary production controls. Earth Interactions, 2000, 4(3): 1-85. |
| [33] |
Zhen S, Jia S F, Lv A F, Jesper S, Gao Y C. Impacts of climate change on growth period and planting boundaries of spring wheat in China under RCP4.5 scenario. Journal of Resources and Ecology, 2016, 7(1): 1-11.
|
| [34] |
翟颖佳, 李耀辉, 徐影. RCPs情景下中国北方地区干旱气候变化特征. 高原气象, 2016, 35(1): 94-106. |
| [35] |
张冬峰, 石英. 区域气候模式RegCM3对华北地区未来气候变化的数值模拟. 地球物理学报, 2012, 55(9): 2854-2866. |
| [36] |
韩其飞, 罗格平, 李超凡, 叶辉, 冯异星. 基于Biome-BGC模型的天山北坡森林生态系统碳动态模拟. 干旱区研究, 2014, 31(3): 375-382. |
| [37] |
赵敏, 周广胜. 中国北方林生产力变化趋势及其影响因子分析. 西北植物学报, 2005, 25(3): 466-471. DOI:10.3321/j.issn:1000-4025.2005.03.008 |
| [38] |
叶兵.北京延庆小叶杨与刺槐林的蒸腾耗水特性与水量平衡研究[D].北京: 中国林业科学研究院, 2007.
|
| [39] |
Kang M C, Zhang Z Q, Noormets A, Fang X R, Zha T G, Zhou J, Sun G, McNulty S G, Chen J Q. Energy partitioning and surface resistance of a poplar plantation in northern China. Biogeosciences, 2015, 12(14): 4245-4259. DOI:10.5194/bg-12-4245-2015 |
| [40] |
赵俊芳, 延晓冬, 贾根锁. 东北森林净第一性生产力与碳收支对气候变化的响应. 生态学报, 2008, 28(1): 92-102. DOI:10.3321/j.issn:1000-0933.2008.01.011 |
| [41] |
张臻.时空异质条件下的大气CO2施肥效应对全球碳水循环影响的模拟研究[D].南京: 南京大学, 2013.
|
| [42] |
徐胜, 陈玮, 何兴元, 黄彦青, 高江艳, 赵诣, 李波. 高浓度CO2对树木生理生态的影响研究进展. 生态学报, 2015, 35(8): 2452-2460. |
| [43] |
Huang M T, Piao S L, Sun Y, Ciais P, Cheng L, Mao J F, Poulter B, Shi X Y, Zeng Z Z, Wang Y P. Change in terrestrial ecosystem water-use efficiency over the last three decades. Global Change Biology, 2015, 21(6): 2366-2378. DOI:10.1111/gcb.12873 |
| [44] |
刘国华, 傅伯杰. 全球气候变化对森林生态系统的影响. 自然资源学报, 2001, 16(1): 71-78. DOI:10.3321/j.issn:1000-3037.2001.01.013 |
| [45] |
Niu S L, Xing X R, Zhang Z, Xia J Y, Zhou X H, Song B, Li L H, Wan S Q. Water-use efficiency in response to climate change:from leaf to ecosystem in a temperate steppe. Global Change Biology, 2011, 17(2): 1073-1082. DOI:10.1111/gcb.2010.17.issue-2 |
| [46] |
周洁, 张志强, 孙阁, 方显瑞, 查同刚, 张燕, 王小平, 陈俊崎, 陈吉泉. 不同土壤水分条件下杨树人工林水分利用效率对环境因子的响应. 生态学报, 2013, 33(5): 1465-1474. |
| [47] |
Pan S F, Chen G S, Ren W, Dangal S R S, Banger K, Yang J, Tao B, Tian H Q. Responses of global terrestrial water use efficiency to climate change and rising atmospheric CO2 concentration in the twenty-first century. International Journal of Digital Earth, 2018, 11(6): 558-582. DOI:10.1080/17538947.2017.1337818 |
| [48] |
Keenan T F, Hollinger D Y, Bohrer G, Dragoni D, Munger J W, Schmid H P, Richardson A D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature, 2013, 499(7458): 324-327. DOI:10.1038/nature12291 |