 2019, Vol. 39
2019, Vol. 39文章信息
- 夏珊珊, 胡大明, 邓玥, 钟雪, 白文科, 张晋东, 王彬, 周材权
- XIA Shanshan, HU Daming, DENG Yue, ZHONG Xue, BAI Wenke, ZHANG Jindong, WANG Bin, ZHOU Caiquan
- 同域分布红腹锦鸡和红腹角雉在不同空间尺度下的生境分化
- Habitat partitioning between sympatric Golden Pheasant and Temminck's Tragopan at different spatial scales
- 生态学报. 2019, 39(5): 1627-1638
- Acta Ecologica Sinica. 2019, 39(5): 1627-1638
- http://dx.doi.org/10.5846/stxb201809051894
-
文章历史
- 收稿日期: 2018-09-05
- 网络出版日期: 2019-12-29





2. 西华师范大学生态研究院, 南充 637002;
3. 四川白水河国家级自然保护区, 彭州 611930;
4. 西华师范大学学报编辑部, 南充 637009
2. Institute of Ecology, China West Normal University, Nanchong 637002, China;
3. Baishuihe National Nature Reserve, Pengzhou 611930, China;
4. Editorial Department of Journal, China West Normal University, Nanchong 637009, China
长期以来, 同域动物的共存机制都是群落生态学领域的重要研究内容[1]。根据竞争排斥原理, 完全的竞争者不能共存, 同域物种的长期稳定共存建立在种间的生态位分化上[2-3]。在众多的生态维度中, 生境分化是同域动物最为常见和重要的生态位分化形式[4]。当群落中的不同物种需求相似的限制性资源, 对生境的差异性选择和利用能够降低种间竞争的强度, 促进其稳定共存[5]。动物的生境选择是一个具有层级结构的生态过程, 在不同的空间尺度下会有不同的因子和驱动力来塑造物种的地理分布和栖息地格局[6]。因此, 同域动物的生境分化可能同样具有空间尺度效应, 在不同的空间尺度下受到不同因素的影响, 体现出不同的分化模式和程度[7]。在研究动物同域共存问题时, 忽略空间尺度效应会使我们无法全面了解种间关系和生态位分化, 可能导致片面解读物种的共存模式和机制[8-9]。
鸡形目(Galliformes)鸟类是典型的地栖性鸟类, 较弱的飞行能力使其绝大部分行为都局限于地面生境, 难以在垂直空间上扩展。因此, 同域鸡形目物种可能面临更为激烈的空间竞争, 是检验生境分化的理想类群。目前, 已有一些研究在不同地区探讨了同域鸡形目物种的生境分化, 但已有研究大多只集中在微生境[10-11]或景观[12-13]等单一空间尺度, 缺乏对不同尺度下生境分化差异的关注[14], 限制了对同域鸡形目鸟类生境分化和共存机制的全面认识。
红腹锦鸡(Chrysolophus pictus)和红腹角雉(Tragopan temmminckii)同属鸡形目雉科(Phasianidae), 都是典型的山地森林雉类。红腹锦鸡为我国特有种, 仅分布于我国的中部山地海拔800—1600 m区域, 而红腹角雉主要分布在海拔1000—3500 m的中国西南部和印度等地[15], 并且二者均为以植物性食物为主的杂食性鸟类[16-17]。高度重叠的地理范围和海拔分布, 以及相似的食性, 暗示着红腹锦鸡和红腹角雉具有相似的生境需求和偏好, 同域分布时可能会成为互相的潜在竞争者。同时, 红腹锦鸡和红腹角雉均为我国二级重点保护野生动物, 红腹锦鸡属我国特有物种, 因此二者都具有较高的保护价值。然而, 目前关于二者生境的研究大多只关注单一物种的生境选择和利用[18-19], 尚无研究探讨二者在同域分布情况下的生境分化模式。基于此, 本研究针对同域共存的红腹锦鸡和红腹角雉, 从宏生境(种群共同占据的大块生境)和微生境(个体家域范围内具体活动点周围的环境)[20]两个空间尺度研究二者的生境分化, 旨在(1)分析不同空间尺度下生境选择的种间差异, 确定其生境分化的主要方式和关键尺度;(2)了解二者生境需求的异同, 探讨种间关系, 为区域雉类整合保护提供理论支持。
1 研究地概况研究区域位于四川龙门山脉东南部, 覆盖了四川白水河国家级自然保护区及其周边区域(31°10′—31°28′N, 103°38′—103°57′E), 总面积约991 km2。研究区域温暖湿润, 四季分明, 属于亚热带湿润气候类型, 年均温15.6℃, 年均降水量932.5 mm, 年日照仅711 h, 无霜期长达276 d。研究区域地处四川盆地向青藏高原东缘川西高山峡谷过渡地带, 地形地貌复杂, 地势东南低西北高, 海拔跨度超过4000 m(693—4814 m), 植被垂直带谱明显, 生物多样性丰富。红腹锦鸡和红腹角雉的地理分布在该区域重叠[21], 前期访问调查发现两个物种已长期稳定共存并且均有较大的种群数量。
2 研究方法 2.1 野外调查使用样线法调查研究区域内红腹锦鸡和红腹角雉的分布和生境利用情况。根据研究区域内植被类型和地形条件, 系统布设了15条调查样线, 样线总长度约55 km, 海拔跨度1200—3200 m, 尽可能穿越多种植被类型, 从而尽量覆盖两种雉类的潜在分布范围。2018年1月、3月、5月和8月分别对样线进行重复调查, 当调查中发现红腹锦鸡或红腹角雉实体或羽毛, 记录发现点坐标并建立10 m×10 m样方, 测量并记录反映微生境食物丰富度、安全性和热量维持三方面特征的16个环境变量[22]。首先在样方中心测定海拔、坡向、坡度、坡位和最近水源距离等地形数据;然后在10 m×10 m样方内估测乔木层的盖度、均高和平均胸径;接着将样方等分为4个5 m×5 m的灌木样方, 测量每一样方内灌木的盖度和高度, 以其平均值代表微生境样方的灌木层特征;最后在每一灌木样方的中心设置1 m×1 m的草本样方, 测量4个草本样方内的草本盖度和均高、苔藓盖度、裸地盖度、岩石盖度和落叶盖度, 平均后代表微生境样方的草本层和地表层特征。
当样线调查中听到两个物种的鸣叫时, 记录鸣叫发生地点与调查者之间的距离和方向, 以及调查者所在位置的坐标, 后期在ArcGIS 10.2软件中测算得到鸣叫发生的实际位点。
2.2 宏生境分析采用最大熵模型(MaxEnt)对两种雉类的宏生境进行建模分析。
2.2.1 物种分布点在4次野外调查中, 一共获得了46个红腹锦鸡记录点和35个红腹角雉出现点。为进一步增加出现点样本量从而提高模型准确度, 将白水河保护区本底调查(红腹锦鸡19个, 红腹角雉27个)和红外相机监测(红腹锦鸡2个, 红腹角雉64个)中记录到的物种分布点纳入分析。为避免MaxEnt模型的过度拟合, 按最小活动区原则对分布点进行筛选[23]。已有研究表明, 红腹锦鸡和红腹角雉的最小活动区面积分别为(11.5±3.8) hm2[24]和(8.9±0.8) hm2[25], 分别近似于半径200 m和170 m的圆。在ArcGIS中以这两个距离作为阈值分别对两种雉类的所有出现点进行buffer分析, 当多个出现点间的距离小于各自阈值时, 随机保留其中一个, 其余删除。经过筛选, 最终保留了47个红腹锦鸡出现点和70个红腹角雉出现点进入MaxEnt模型。
2.2.2 环境变量根据以往的雉类物种生境研究经验, 选择了地形、植被、气候和人为干扰等4类共31个潜在影响红腹锦鸡和红腹角雉宏生境选择的环境变量作为MaxEnt模型的备选变量。
地形变量:下载分辨率30 m×30 m的ASTER GDEM V2图层, 并提取海拔、坡度和坡向数据。由于坡向数据属圆周变量, 对其进行实际坡向减180°后取绝对值的处理, 以此来反映各栅格对正南阳坡的靠近程度[26]。在ArcGIS中计算河流图层的欧氏距离(Euclidean distance)以反映各栅格距离其最近河流的距离。
植被变量:下载研究区域分辨率30 m×30 m LANDSAT 8遥感影像数据, 通过ERDAS IMAGINE 9.2对研究区域的遥感影像进行非监督分类, 选取50%的实地调查点(n=127)作为参考点对分类结果进行定义, 根据《中国植被》分类系统将区域内的植被划分为寒温性针叶林、温性针阔混交林、温性针叶林、暖性针叶林、落叶阔叶林、常绿阔叶林、灌丛及灌草丛、高山流石滩、草甸9种植被类型, 其余地表覆盖划分为滑坡体、农耕地、水体3种覆盖类型。以剩余的50%实地调查点(n=127)作为评价点, 提取其分类值与实际植被或覆盖类型比较, 结果显示79.5%的评价点的分类结果与实地调查一致, 总体分类精度良好[27]。下载研究区域分辨率250 m×250 m的MOD13Q1植被指数16天合成产品2017年全年数据, 提取增强型植被指数(Enhanced Vegetation Index, EVI)。在ArcGIS中对全年23个EVI图层进行计算, 分别得到年最大EVI、年最小EVI、年均EVI、年EVI变幅和年EVI标准差。EVI衍生变量与植被初级生产力及其季节性变化密切相关[28], 有助于提高生态位模型的准确度[29]。
气候变量:气候对物种的分布、生存和繁殖等都有直接影响[30], 被广泛用于生态位模型分析。从WorldClim数据库下载得到WorldClim 2.0的19个生物气候变量[31], 作为备选环境变量。
人为干扰变量:人类的活动会对动物栖息地产生影响, 而居住和交通是最为直接和强烈的两种方式。因此, 利用ArcGIS软件计算公路和居民点图层的欧氏距离以分别反映各栅格距离其最近公路和居民的距离。
将31个环境变量统一重采样为30 m×30 m的栅格图层, 其中分类变量(植被类型)以最邻近分配法重采样, 其余30个连续型变量以双线性插值法重采样, 进入后续分析。
2.2.3 模型过程为避免环境变量的空间共线性对模型准确性的影响[32], 对上述31个备选环境变量进行相关性分析, 当多个变量间相关系数大于±0.8, 仅随机保留其中一个。最终保留了14个环境变量进入MaxEnt模型:海拔、坡向、坡度、距河流距离、植被类型、年均EVI、年EVI变幅、等温性(bio3)、温度季节性(bio4)、年温差(bio7)、降水季节性(bio15)、最干季降水(bio17)、距居民点距离和距公路距离。
使用MaxEnt 3.3.3k建立模型[33], 随机选取75%的出现点作为训练数据集, 剩余25%作为检验数据集。对每个物种分别进行10次自举法(bootstrap)重复以保证模型结果的稳定性。以受试者工作特征曲线(ROC)下的面积(area under curve, AUC)对模型结果进行评价, 其评判标准为:AUC在0.5—0.6为失败;0.6—0.7为较差;0.7—0.8为一般;0.8—0.9为好;0.9—1为极好[34]。
2.2.4 种间宏生境比较分析以10次重复后MaxEnt输出的平均宏生境适宜性指数作为最终的模型结果。根据最大约登指数将研究区域内的所有栅格二分类为适宜/不适宜宏生境。最大约登指数, 即敏感性和特异性取最大值时的阈值, 能将模型结果正观察和负观察错误率的平均值最小化, 在仅有物种出现点的模型方法中具有较高的准确性[35]。以软件内建的变量贡献分析和响应曲线来分析各环境变量在模型中的相对重要性及其对物种宏生境适宜性的影响, 并比较两种雉类在影响因子重要性和选择偏好上的异同。
为量化研究区域内两种雉类宏生境的相似程度, 使用软件ENMTools基于两种雉类MaxEnt输出的生境适宜性指数计算两种雉类宏生境生态位重叠指数D和I[36-37]。重叠指数D和I的取值都从0(两者生态位没有重叠)到1(两者生态位完全一致)。
2.3 微生境分析 2.3.1 变量分级为了尽可能减小调查人员主观差异导致的测量偏差, 将微生境样方调查中人为估算的连续型变量进行分级处理, 具体划分见表 1。
| 变量 Variable |
等级划分Ordinal division | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 坡位Slope position | 沟谷 | 下 | 中 | 上 | 山脊 | |
| 坡度Slope | 0—5 | 6—20 | 21—30 | 31—40 | >40 | |
| 距水源距离Distance to water resources/m | 0—50 | 51—100 | >100 | |||
| 乔木均高Mean tree height/m | 无 | 5—9 | 10—19 | 20—29 | >30 | |
| 乔木盖度Tree coverage/% | 无 | 0—24 | 25—49 | 50—74 | 75—100 | |
| 乔木胸径Tree diameter at breast height/cm | 无 | 0—10 | 11—20 | 21—30 | >31 | |
| 灌木均高Mean shrub height/m | 无 | 0—1 | 1—2 | 2—3 | 3—4 | 4—5 |
| 灌木盖度Shrub coverage/% | 无 | 0—24 | 25—49 | 50—74 | 75—100 | |
| 草本均高Mean herb height/m | 无 | 0—5 | 6—10 | 11—15 | 16—20 | >20 |
| 草本盖度Herb coverage/% | 无 | 0—24 | 25—49 | 50—74 | 75—100 | |
| 苔藓盖度Moss coverage/% | 无 | 0—10 | 11—20 | 21—30 | 31—40 | >41 |
| 裸地盖度Bare land coverage/% | 无 | 0—10 | 11—20 | 21—30 | 31—40 | >41 |
| 岩石盖度Rock coverage/% | 无 | 0—10 | 11—20 | 21—30 | 31—40 | >41 |
| 落叶盖度Litter coverage/% | 无 | 0—24 | 25—49 | 50—74 | 75—100 | |
为整合众多微生境变量的共同变化模式, 并且克服多重共线性问题, 对所有连续型变量进行主成分分析(Principal Components Analysis, PCA)。提取特征根(Eigenvalue)>1的成分, 保留最大方差法标准旋转后的因子得分, 作为降维后的微生境变量进入后续分析。用单因素多元方差分析(One-Way Multivariate Analysis of Variance, MANOVA)比较两种雉类微生境特征的整体差异。若MANOVA发现两种雉类的微生境存在显著的整体种间差异, 进一步用单因素方差分析(One-Way Analysis of Variance, ANOVA)比较每一微生境特征成分的种间差异。为量化两种雉类微生境特征的相似程度, 用Maurer[38]重叠指数公式计算二者在各成分上的重叠指数:
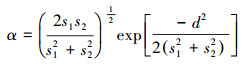
|
(1) |
式中, α即种间微生境特征重叠指数, s1和s2分别是物种1和2在给定成分上的标准差, d是两物种这一成分平均值差值的绝对值。采用积模型计算整体微生境特征的种间重叠指数, 即计算为所有成分上的重叠指数的乘积[38]。重叠指数的范围从0(微生境完全分离)到1(微生境完全重叠)[39]。
统计分析中除特别说明外, 结果以平均值±标准误显示, 视P < 0.05为统计显著。
3 结果 3.1 宏生境在10次重复后得到两种雉类的模型ROC曲线验证结果:红腹锦鸡训练AUC=0.972, 验证AUC=0.950, 红腹角雉训练AUC=0.955, 验证AUC=0.927, 表明两种雉类的MaxEnt模型结果均达到极好水平, 能够较准确预测两种雉类的宏生境适宜性。
3.1.1 宏生境适宜性影响因子变量贡献性分析显示, 等温性、距居民点距离、最干季降水、植被类型和距河流距离是影响红腹锦鸡宏生境适宜性的主要环境因子, 累计贡献率达到75.7%;而红腹角雉则有所不同, 其最主要的影响因子是等温性和最干季降水, 降水季节性次之, 累计贡献率达76.4%(表 2)。
| 环境变量 environment variable | 红腹锦鸡 Golden Pheasant 贡献性 Percent contribution/% | 红腹角雉 Temminck′s Tragopan 贡献性 Percent contribution/% |
| 等温性Isothermality | 36.8 | 35.6 |
| 距居民点距离Distance to residential location | 13.2 | 1.5 |
| 最干季降水Precipitation of the driest quarter | 11.1 | 35.4 |
| 植被类型Vegetation type | 9.6 | 4.1 |
| 距河流距离Distance to river | 5.0 | 1.7 |
| 增强型植被指数年变幅Annual range of enhanced vegetation index | 4.4 | 2.8 |
| 坡向Aspect | 4.0 | 2.8 |
| 年温差Temperature annual range | 3.8 | 2.8 |
| 温度季节性Temperature seasonality | 3.1 | 2.4 |
| 降水季节性Precipitation seasonality | 3.0 | 5.4 |
| 距公路距离Distance to road | 2.2 | 2.1 |
| 坡度Slope | 2.0 | 1.0 |
| 年均增强型植被指数Annual mean enhanced vegetation index | 1.7 | 1.7 |
| 海拔Altitude | 0.2 | 0.6 |
等温性和最干季降水是两种雉类共同的重要影响因子, 并且环境变量响应曲线显示它们对两种雉类宏生境适宜性有相似的影响:两种雉类都偏好等温性较高的区域, 在等温性约39—40时达到适宜性峰值;两种雉类都偏好最干季降水量大的区域, 并且适宜性都在39 mm左右达到峰值。而对于其他的重要环境因子, 两种雉类表现出了一定的种间差异:红腹角雉比红腹锦鸡更加倾向于选择远离居民点的生境, 两者分别在距居民点1800 m和1000 m时达到最大适宜性;红腹锦鸡主要选择落叶阔叶林、暖性针叶林、温性针阔混交林和农耕地, 而红腹角雉则偏好更多类型的植被, 包括温性针叶林、落叶阔叶林、灌丛及灌草丛、温性针阔混交林和常绿阔叶林;虽然二者都倾向于选择靠近河流的生境, 但红腹锦鸡受距河流距离的影响更为显著, 其宏生境适宜性随着远离河流快速下降;红腹角雉偏好降水季节性较低的生境, 其宏生境适宜性在74左右最高, 红腹锦鸡的适宜性峰值则出现于81左右(图 1)。

|
| 图 1 红腹锦鸡和红腹角雉宏生境适宜性对重要环境因子的响应曲线 Fig. 1 Response curves of macrohabitat suitability of Golden Pheasant and Temminck′s Tragopan to important environmental factors |
红腹锦鸡和红腹角雉MaxEnt模型的最大约登指数分别为0.245和0.247, 据此将两种雉类的模型结果二分类为适宜/非适宜宏生境(图 2)。在研究区域内, 红腹锦鸡的适宜宏生境面积为75.92 km2, 占研究区域总面积的7.7%, 其中39.35 km2位于白水河保护区内, 占保护区面积的11.8%;红腹角雉适宜宏生境面积100.60 km2, 占研究区域总面积的10.2%, 其中70.50 km2位于白水河保护区内, 占保护区面积的23.4%。从空间分布上来看, 两种雉类的适宜宏生境都依龙门山脉山势, 呈条带状沿东北-西南走向分布于白水河保护区的东南边界两侧, 并且存在较大面积的相互重叠, 重叠面积达44.59 km2, 分别占红腹锦鸡和红腹角雉适宜宏生境面积的58.73%和44.3%。虽然如此, 红腹角雉的适宜宏生境分布更靠西北, 即沿山势海拔更高的区域, 而红腹锦鸡的宏生境则主要分布在海拔稍低的东南部, 两者之间存在一定的平移。同时, 两者之间宏生境重叠指数也较高:D= 0.578, I = 0.835, 表明两种雉类的适宜宏生境在空间分布上具有较高的相似性。

|
| 图 2 红腹锦鸡和红腹角雉适宜宏生境分布图 Fig. 2 Suitable macrohabitat distribution of Golden Pheasant and Temminck′s Tragopan |
野外调查中共发现并记录红腹锦鸡微生境样方32个, 红腹角雉微生境样方34个。
3.2.1 主成分分析对野外获得的所有微生境利用样方进行主成分分析, 结果显示, 前5个成分的特征根>1, 累计贡献率达到62%, 基本能够解释两种雉类微生境特征的变化模式。其中, 成分1与海拔、坡位、最近水源距离正相关, 与乔木层盖度负相关;成分2表示了更大的灌木层均高和盖度, 而与乔木平均胸径和落叶盖度负相关;成分3反映了更发达的草本层和更高的裸地盖度;成分4反映了更靠阴坡的坡向, 更大的坡度和更大的苔藓盖度;成分5与乔木层均高和平均胸径正相关, 与岩石盖度负相关(表 3)。
| 微生境变量 Microhabitat variable |
成分Component | ||||
| 1 | 2 | 3 | 4 | 5 | |
| 海拔Altitude | 0.76 | 0.02 | -0.27 | -0.25 | -0.29 |
| 坡向Aspect | -0.02 | 0.22 | -0.26 | 0.62 | 0.07 |
| 坡位Slope position | 0.82 | 0.21 | 0.08 | -0.10 | 0.14 |
| 坡度Slope | 0.02 | -0.12 | 0.35 | 0.70 | -0.23 |
| 最近水源距离Nearest water source distance | 0.73 | 0.02 | -0.05 | 0.21 | 0.08 |
| 乔木层均高Mean tree height | -0.23 | 0.05 | -0.11 | 0.35 | 0.71 |
| 乔木层盖度Tree coverage | -0.63 | -0.36 | -0.01 | 0.09 | 0.36 |
| 乔木层平均胸径Mean tree diameter at breast height | 0.02 | -0.64 | 0.04 | 0.04 | 0.55 |
| 灌木层均高Mean shrub height | 0.32 | 0.76 | -0.15 | 0.06 | -0.05 |
| 灌木层盖度Shrub coverage | 0.33 | 0.57 | 0.02 | 0.02 | -0.05 |
| 草本层均高Mean herb height | -0.05 | 0.20 | 0.77 | 0.05 | 0.14 |
| 草本层盖度Herb coverage | -0.05 | -0.02 | 0.79 | -0.19 | 0.04 |
| 苔藓盖度Moss coverage | 0.06 | 0.23 | 0.23 | -0.70 | -0.16 |
| 裸地盖度Bare land coverage | -0.02 | -0.29 | 0.41 | -0.32 | -0.26 |
| 岩石盖度Rock coverage | -0.07 | 0.06 | -0.16 | 0.15 | -0.68 |
| 落叶盖度Litter coverage | 0.08 | -0.68 | -0.28 | 0.12 | -0.10 |
| 特征根Eigenvalue | 3.39 | 2.24 | 1.73 | 1.48 | 1.17 |
| 解释方差百分比Percent of variance | 21.18 | 13.99 | 10.78 | 9.23 | 7.33 |
| 累计解释方差Cumulative percent | 21.18 | 35.18 | 45.96 | 55.19 | 62.53 |
MANOVA显示, 红腹锦鸡和红腹角雉的微生境特征存在着显著的种间差异(Wilks′ λ=0.641, F=6.717, P < 0.001)。进一步的ANOVA显示, 红腹锦鸡的成分1显著小于红腹角雉, 其余成分没有显著的种间差异(表 4)。根据主成分分析的因子载荷, 红腹锦鸡微生境的海拔和坡位低于红腹角雉, 更加靠近水源并且有更高的乔木盖度(表 3)。Maurer重叠指数的计算显示, 两种雉类的整体重叠指数较低, 仅为0.523, 其中成分1上的重叠最小, 而在其他成分上两种都有极高的相似性(表 4)。
| 成分 Component |
均值 Mean value |
单因素方差分析 ANOVA |
重叠指数 Overlap index |
||||
| 红腹锦鸡 Golden Pheasant |
红腹角雉 Temminck′s Tragopan |
F | P | α | |||
| 成分1 | -0.556±0.155 | 0.523±0.140 | 26.794 | <0.001 | 0.666 | ||
| 成分2 | 0.005±0.163 | -0.005±0.186 | 0.002 | 0.966 | 0.994 | ||
| 成分3 | 0.117±0.153 | -0.110±0.191 | 0.841 | 0.363 | 0.972 | ||
| 成分4 | 0.131±0.121 | -0.123±0.210 | 1.069 | 0.305 | 0.908 | ||
| 成分5 | 0.190±0.121 | -0.178±0.207 | 2.277 | 0.136 | 0.895 | ||
生境分化是同域分布近缘物种间生态位分化最为普遍和重要的形式, 它能够降低物种对空间和其他环境资源的种间竞争, 从而促进近缘物种的相互共存[40]。本研究从宏生境和微生境两个空间尺度, 对同域分布的红腹锦鸡和红腹角雉的生境分化进行了研究, 结果表明其生境分化在不同的空间尺度下表现出不同的分化程度和方式, 但又兼具一定的一致性。
宏生境尺度上, 红腹锦鸡和红腹角雉适宜宏生境的空间分布存在大面积的相互重叠(图 2), 并且有非常高的宏生境重叠指数[41-42], 表明二者在宏生境尺度上没有发生明显的种间分化。高度重叠的宏生境主要是由于等温性和最干季降水是影响两种雉类生境适宜性的共同重要因子, 并且两种雉类对其具有相似的选择性(图 2)。这与刘小斌[43]的研究结果相似, 其基于红外相机的监测结果显示这两种雉类的活动温度范围具有相同的趋势。然而, 两种雉类对其他重要影响因子则有不同的响应模式, 并且这些差异在一定程度上反映了两个物种对不同海拔带的适应性:红腹锦鸡对来自居民点的人为干扰耐受性更高, 更加依赖河流, 并且对农耕地生境有一定的利用, 使得红腹锦鸡更加适应于居民点、农耕地和河流都更密集的低海拔带;而红腹角雉更加回避居民点, 偏好更低的降水季节性, 并且能够选择分布于高海拔的灌丛及灌草丛生境, 所以这一物种更适应于人为干扰较少, 年降水量较为稳定的亚高山带。这些差异使得红腹锦鸡与红腹角雉的适宜宏生境在空间分布上发生了一定的分离, 即红腹角雉的宏生境沿山势分布于比红腹锦鸡海拔更高的区域(图 2), 这与两种雉类自身的分布海拔差异相一致[15]。这种同域分布物种间不同的资源响应与选择策略, 一定程度上减少了物种空间分布上的重叠, 从而减轻种间竞争强度[44]。
在微生境尺度上, 红腹锦鸡和红腹角雉在海拔、坡位、最近水源距离和乔木层盖度等微生境特征上表现出了显著的种间差异, 表明了两者微生境的显著分化。海拔变化是影响野生动物垂直分布格局的重要因素[45], 红腹角雉利用微生境的海拔高度显著高于红腹锦鸡, 与二者在宏生境尺度上的空间分离相一致, 同样表明了两个物种对不同海拔带的适应性。红腹锦鸡的利用微生境比红腹角雉更加靠近水源, 这也与宏生境尺度的分析一致, 进一步表明了水源在红腹锦鸡生境选择中的重要性。相比之下, 而红腹角雉能够在更远离水源的微生境生活, 可能是由于白水河保护区内降雨丰富, 红腹角雉能够从雨水或植物中获取足够的水分。红腹锦鸡微生境的坡位低于红腹角雉, 与李宏群[46]和丛培昊[47]分别对红腹锦鸡(偏好下坡位)和红腹角雉(偏好上坡位)的研究结果一致, 这与二者对海拔的不同选择和对水源不同程度的依赖有关。此外, 红腹锦鸡比红腹角雉偏好乔木层盖度更高的微生境, 这主要是因为红腹锦鸡只活动于乔木层发达的森林之中, 而红腹角雉在利用森林生境的同时, 还会利用乔木层稀疏的灌丛及灌草丛生境, 导致了两者微生境乔木层盖度的差异。对这些微生境变量的差异性利用, 塑造了红腹锦鸡和红腹角雉间分化显著的微生境利用模式, 进一步缓解了两者对相同资源的种间竞争, 是其生境分化的关键尺度, 使两个物种即使在高度重叠的宏生境之内也能实现稳定共存。
雉类在维持生态系统稳定和生物多样性保育等方面发挥着重要的作用, 其生存状况在一定程度上可以作为反映当地生态系统健康状况的有效指标[48]。红腹锦鸡和红腹角雉均为我国二级重点保护野生动物, 红腹锦鸡属我国特有物种, 因此二者都具有较高的保护价值。本研究发现, 两种雉类虽然在宏生境上重叠程度较大, 但其微生境的利用存在显著的种间分化, 使其能够分享共同栖息地内的有限资源而无强烈的种间竞争关系。因此, 对于其中某一物种的保护管理不会对另一物种造成负面影响[11, 49]。从宏生境的空间分布来看, 研究区域内两种雉类的大部分适宜宏生境已处于由白水河及相邻的龙溪-虹口和九顶山保护区形成的保护网络当中, 得到了良好的保护。但仍有小部分的适宜宏生境坐落于保护区外, 建议未来对这些适宜宏生境斑块增加关注和监测, 使生存其中的雉类种群得到有效保护。本研究结果显示, 两种雉类对宏生境和微生境的需求存在一些异同, 能够为未来的生境管理提供一定的理论支持。首先, 虽然响应程度不同, 但两种雉类的宏生境选择都倾向于远离居民点, 暗示了人为干扰对于雉类而言是一种消极影响。而在调查中发现, 白水河保护区内及周边旅游区域多且客流量大, 同时当地社区百姓在夏季有进山采药的传统, 这些活动都对雉类种群及其生境带来了直接干扰。因此, 建议一方面通过严格控制进山人数, 合理设计旅游区域等手段, 尽可能减少人为干扰的强度;另一方面, 加强对保护区周边社区居民及进山旅游人群的宣传教育, 普及法律法规, 尽量降低盗猎、拾卵和垃圾污染等的可能性。其次, 虽然两种雉类都偏好针阔混交林、常绿阔叶林和落叶阔叶林, 但是对于其他植被类型的选择则存在差异, 并且二者对微生境乔木层盖度的偏好也有不同。因此, 对两种雉类生境的管理中, 除了应重点保护其共同需求的自然植被外, 还应注意维持植被类型的多样性和镶嵌格局, 在恢复栖息地时需避免单一树种的人工造林。此外, 白水河保护区的南部, 有零星的适宜宏生境与龙溪-虹口保护区的灯草坪区域相连接, 而在北部, 也有一定的适宜宏生境延伸至九顶山保护区内。这些相互连通的生境可能为两种雉类在不同保护区之间的扩散和迁移提供了廊道, 在未来的管理中应重点保护, 避免生境破碎化的发生。
本研究探讨了红腹锦鸡和红腹角雉两种典型山地森林雉类在宏生境和微生境尺度的分化程度和方式, 初步揭示了其同域共存的模式。然而, 值得注意的是, 由于数据获取测定和获取的难度性, 本研究依然存在一定的局限性。首先, 宏生境建模所使用的WorldClim数据虽然是目前全球广泛使用的气象数据图层, 但在龙门山脉这种山峰沟谷交错, 海拔落差较大的山区, 其1×1 km的空间分辨率难以准确反映微气候的快速变化, 存在一定局限性。另一方面, 本研究虽然揭示了两种雉类全年的整体生境分化模式, 但雉类物种的生境选择和利用通常存在季节性变化[50], 其生境分化可能也因此存在季节性, 然而囿于有限的数据量, 本研究未分季节进行生境分化的探讨, 也是一点遗憾。因此, 建议在未来通过开展更多调查, 积累充足的物种生境利用数据, 对两种雉类不同季节的生境分化进行比较分析, 从而进一步揭示其同域共存模式的潜在季节性变异。
| [1] |
Begon M, Harper J L, Townsend C R. Ecology:Individuals, Populations and Communities4th ed. Oxford: Blackwell Science, 2006: 407-408.
|
| [2] |
Gause G F. The Struggle for Existence. New York: Hafner, 1934.
|
| [3] |
Hardin G. The competitive exclusion principle. Science, 1960, 131(3409): 1292-1297. DOI:10.1126/science.131.3409.1292 |
| [4] |
Schoener T W. Resource partitioning in ecological communities. Science, 1974, 185(4145): 27-39. DOI:10.1126/science.185.4145.27 |
| [5] |
Chesson P. Mechanisms of maintenance of species diversity. Annual review of Ecology and Systematics, 2000, 31: 343-366. DOI:10.1146/annurev.ecolsys.31.1.343 |
| [6] |
George T L, Zack S. Spatial and temporal considerations in restoring habitat for wildlife. Restoration Ecology, 2001, 9(3): 272-279. DOI:10.1046/j.1526-100x.2001.009003272.x |
| [7] |
Davies K F, Chesson P, Harrison S, Inouye B D, Melbourne B A, Rice K J. Spatial heterogeneity explains the scale dependence of the native-exotic diversity relationship. Ecology, 2005, 86(6): 1602-1610. DOI:10.1890/04-1196 |
| [8] |
Boeye J, Kubisch A, Bonte D. Habitat structure mediates spatial segregation and therefore coexistence. Landscape Ecology, 2014, 29(4): 593-604. DOI:10.1007/s10980-014-0010-6 |
| [9] |
Pita R, Lambin X, Mira A, Beja P. Hierarchical spatial segregation of two Mediterranean vole species:the role of patch-network structure and matrix composition. Oecologia, 2016, 182(1): 253-263. DOI:10.1007/s00442-016-3653-y |
| [10] |
刘鹏, 黄晓凤, 顾署生, 鲁长虎. 江西官山自然保护区四种雉类的生境选择差异. 动物学研究, 2012, 33(2): 170-176. |
| [11] |
Sachot S, Perrin N, Neet C. Winter habitat selection by two sympatric forest grouse in western Switzerland:implications for conservation. Biological Conservation, 2003, 112(3): 373-382. DOI:10.1016/S0006-3207(02)00334-8 |
| [12] |
Hagen C A, Pitman J C, Robel R J, Loughin T M, Applegate R D. Niche partitioning by lesser prairie-chicken Tympanuchus pallidicinctus and ring-necked pheasant Phasianus colchicus in southwestern Kansas. Wildlife Biology, 2007, 13(S1): 34-41. |
| [13] |
Jia F, Wang N, Zheng G M. Winter habitat requirements of white eared-pheasant Crossoptilon crossoptilon and blood pheasant Ithaginis cruentus in south-west China. Bird Conservation International, 2005, 15(3): 303-312. DOI:10.1017/S095927090500047X |
| [14] |
崔鹏, 康明江, 邓文洪. 繁殖季节同域分布的红腹角雉和血雉的觅食生境选择. 生物多样性, 2008, 16(2): 143-149. DOI:10.3321/j.issn:1005-0094.2008.02.006 |
| [15] |
郑光美. 中国雉类. 北京: 高等教育出版社, 2015.
|
| [16] |
何少文, 巩会生. 红腹锦鸡的越冬生态观察. 动物学杂志, 1994, 29(6): 47-48. |
| [17] |
史海涛, 郑光美. 红腹角雉取食栖息地选择的研究. 动物学研究, 1999, 20(2): 131-136. DOI:10.3321/j.issn:0254-5853.1999.02.009 |
| [18] |
梁伟.红腹锦鸡(Chrysolophus pictus)的活动性及栖息地选择研究[D].北京: 北京师范大学, 1997.
|
| [19] |
邵晨. 红腹锦鸡的冬季栖息地. 动物学杂志, 1998, 33(2): 38-41. DOI:10.3969/j.issn.0250-3263.1998.02.013 |
| [20] |
Mayor S J, Schneider D C, Schaefer J A, Mahoney S P. Habitat selection at multiple scales. Écoscience, 2009, 16(2): 238-247. DOI:10.2980/16-2-3238 |
| [21] |
郑光美. 中国鸟类分类与分布名录 (第三版). 北京: 科学出版社, 2017.
|
| [22] |
Sutherland W J, Newton I, Green R. Bird Ecology and Conservation:A Handbook of Techniques. Oxford, New York: Oxford University Press, 2004.
|
| [23] |
Wang B, Xu Y, Ran J. Predicting suitable habitat of the Chinese monal (Lophophorus lhuysii) using ecological niche modeling in the Qionglai Mountains, China. PeerJ, 2017, 5: e3477. DOI:10.7717/peerj.3477 |
| [24] |
梁伟.红腹锦鸡(Chrysolophus pictus)的生态适应与保护[D].北京: 北京师范大学, 2000.
|
| [25] |
史海涛.红腹角雉(Tragopan temminckii)的活动区、食性及栖息地的研究[D].北京: 北京师范大学, 1995.
|
| [26] |
Kalkhan M A. Spatial Statistics:GeoSpatial Information Modeling and Thematic Mapping. Boca Raton: CRC Press, 2011.
|
| [27] |
邓书斌. ENVI遥感图像处理方法. 北京: 科学出版社, 2010.
|
| [28] |
Alcaraz-Segura D, Paruelo J M, Epstein H E, Cabello J. Environmental and human controls of ecosystem functional diversity in temperate South America. Remote Sensing, 2013, 5(1): 127-154. DOI:10.3390/rs5010127 |
| [29] |
Requena-Mullo J M, López E, Castro A J, Cabello J, Virgós E, González-Miras E, Castro H. Modeling spatial distribution of European badger in arid landscapes:an ecosystem functioning approach. Landscape Ecology, 2014, 29(5): 843-855. DOI:10.1007/s10980-014-0020-4 |
| [30] |
Guisan A, Zimmermann N E. Predictive habitat distribution models in ecology. Ecological Modelling, 2000, 135(2/3): 147-186. |
| [31] |
Hijmans R J, Cameron S E, Parra J L, Jones P G, Jarvis A. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 2005, 25(15): 1965-1978. DOI:10.1002/(ISSN)1097-0088 |
| [32] |
Swanepoel L H. Extent and fragmentation of suitable leopard habitat in South Africa. Animal Conservation, 2013, 16(1): 41-50. DOI:10.1111/acv.2013.16.issue-1 |
| [33] |
Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 2006, 190(3/4): 231-259. |
| [34] |
Swets J A. Measuring the accuracy of diagnostic systems. Science, 1988, 240(4857): 1285-1293. DOI:10.1126/science.3287615 |
| [35] |
Jiménez-Valverde A, Lobo J M. Threshold criteria for conversion of probability of species presence to either-or presence-absence. Acta Oecologica, 2007, 31(3): 361-369. DOI:10.1016/j.actao.2007.02.001 |
| [36] |
Warren D L, Glor R E, Turelli M. ENMTools:a toolbox for comparative studies of environmental niche models. Ecography, 2010, 33(3): 607-611. |
| [37] |
Warren D L, Glor R E, Turelli M. Environmental niche equivalency versus conservatism:quantitative approaches to niche evolution. Evolution, 2008, 62(11): 2868-2883. DOI:10.1111/evo.2008.62.issue-11 |
| [38] |
Maurer B A. Statistical inference for MacArthruy-Levins niche overlap. Ecology, 1982, 63(6): 1712-1719. DOI:10.2307/1940113 |
| [39] |
May R M. Some notes on estimating the competition matrix. Ecology, 1975, 56(3): 737-741. DOI:10.2307/1935511 |
| [40] |
Parra G J. Resource partitioning in sympatric delphinids:space use and habitat preferences of australian snubfin and indo-pacific humpback dolphins. Journal of Animal Ecology, 2006, 75(4): 862-874. DOI:10.1111/j.1365-2656.2006.01104.x |
| [41] |
Hu J H, Jiang Z G, Chen J, Qiao H J. Niche divergence accelerates evolution in Asian endemic Procapra gazelles. Scientific Reports, 2015, 5: 10069. DOI:10.1038/srep10069 |
| [42] |
Hosseinian Yousefkhani S S, Mirshamsi O, Ilgaz Ç, Kumlutas Y, Avci A. Ecological niche divergence between Trapelus ruderatus (Olivier, 1807) and T. persicus (Blanford, 1881) (Sauria:Agamidae) in the Middle East. Asian Herpetological Research, 2016, 7(2): 96-102. |
| [43] |
刘小斌, 韦伟, 郑筱光, 赵凯辉, 何少文, 周文良. 红腹锦鸡和红腹角雉活动节律——基于红外相机监测数据. 动物学杂志, 2017, 52(2): 194-202. |
| [44] |
Tokeshi M. Species Coexistence:Ecological and Evolutionary Perspectives. Oxford: Blackwell Science, 1999.
|
| [45] |
徐言朋, 郑家文, 丁平, 蒋萍萍, 蔡路昀, 黄晓风, 姚小华, 徐向荣, 余泽平. 官山白颈长尾雉活动区域海拔高度的季节变化及其影响因素. 生物多样性, 2007, 15(4): 337-343. DOI:10.3321/j.issn:1005-0094.2007.04.002 |
| [46] |
李宏群, 韩宗先, 吴少斌, 曹长雷, 岳碧松. 大木山自然保护区红腹锦鸡对冬季生境的选择性. 东北林业大学学报, 2011, 39(11): 76-78. DOI:10.3969/j.issn.1000-5382.2011.11.021 |
| [47] |
丛培昊, 郑光美. 四川老君山地区红腹角雉的夜栖行为和夜栖地选择. 生物多样性, 2008, 16(4): 332-338. DOI:10.3321/j.issn:1005-0094.2008.04.003 |
| [48] |
张正旺, 丁长青, 丁平, 郑光美. 中国鸡形目鸟类的现状与保护对策. 生物多样性, 2003, 11(5): 414-421. DOI:10.3321/j.issn:1005-0094.2003.05.007 |
| [49] |
Zhang Z J, Wei F W, Li M, Hu J C. Winter microhabitat separation between Giant and Red Pandas in Bashania faberi bamboo forest in Fengtongzhai Nature Reserve. Journal of Wildlife Management, 2006, 70(1): 231-235. DOI:10.2193/0022-541X(2006)70[231:WMSBGA]2.0.CO;2 |
| [50] |
刘鹏, 张微微. 官山自然保护区白颈长尾雉季节性生境选择. 生态学报, 2017, 37(18): 6005-6013. |