 2019, Vol. 39
2019, Vol. 39文章信息
- 排孜丽耶·合力力, 吾玛尔·阿布力孜, 阿丽亚·司地克
- Pazliya HELIL, Omar ABLIZ, Aliya SIDIK
- 新疆天山森林公园土壤螨类群落多样性与环境因子的相关性
- Correlation between the diversity of soil mite communities and environmental factors in Tianshan Forest Park in Xinjiang, China
- 生态学报. 2019, 39(5): 1639-1652
- Acta Ecologica Sinica. 2019, 39(5): 1639-1652
- http://dx.doi.org/10.5846/stxb201711302157
-
文章历史
- 收稿日期: 2017-11-30
- 网络出版日期: 2018-12-21

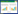




土壤螨类是森林生态系统土壤动物的重要组成部分, 通过自身活动与摄食参与土壤有机质分解和矿化, 改善土壤结构, 调控地上与地下生物间的物质循环和能量流动[1-3];而且对环境变化敏感, 是指示植被演替、环境干扰及气候变化等的重要生物学指标[4-5]。研究表明, 生境变化和地表植被类型通过改变凋落物的组成与数量, 对直接取食凋落物的土壤螨类会产生显著影响[6-8];地上植被组成的改善也会通过土壤理化性质改变而间接影响土壤螨类的分布, 如使土壤螨类类群更丰富、结构更复杂、群落结构异质性和多样性更高[9-10]。另外, 在森林生态系统中放牧和旅游等人类活动、土地利用类型(林型)、土壤微环境及其管理方式等显著影响土壤螨类群落结构[10-14]。到目前为止, 我国从热带、亚热带到温带, 从森林、草地、沙漠到湿地、农田到城市等诸多生态系统都开展了相关研究。从全国范围来看, 已有的研究主要集中在东部, 而西北干旱区, 尤其是新疆土壤螨类群落多样性方面研究报道还很少, 有关土壤螨类群落多样性与环境因素关系的研究更是鲜见报道[15-19]。
新疆天山森林公园位于乌鲁木齐市东南部的天山北坡, 距离乌鲁木齐70 km, 1992年12月确定为自治区级森林公园, 总面积为3918 km2, 主要树种为云杉。森林生长在阴坡, 集中分布在海拔1400—2800 m之间, 森林呈块状或带状与山地草原相间分布, 其建群树种是雪岭云杉(Picea schrenkiana), 在整个林区占绝对优势, 并与其他乔灌木构成有规律的植被垂直景观。天山森林公园风景区具有雪峰、森林、草地、高山、怪石、瀑布等多种自然景观和植被类型。由于以往交通不便等原因, 使其仍然保留着森林生态系统的原始性和自然资源的丰富性。但近年来, 由于过度放牧和旅游活动频繁, 该森林公园生态环境已出现不同程度的退化, 土壤质量状况以及土壤生物多样性等都发生了较大的变化。迄今为止, 针对天山森林公园的研究主要涉及到植被、昆虫以及脊椎动物生态分布等方面。然而, 天山森林公园土壤螨类群落多样性及其与主要环境影响因素关系尚不清楚。因此, 本研究将深入了解土壤螨类群落多样性及其与环境因素间的关系, 为森林生态系统科学管理、土壤环境质量生物评价及生物多样性保护提供科学依据。
1 材料与方法 1.1 研究区概况和样地选择研究区域位于新疆米泉林场(87°48′—88°08′E, 43°42′—43°56′N), 海拔1400—3200 m, 东西宽26 km, 南北长23 km, 林区总面积4万hm2, 林业用地2.87万hm2, 森林覆盖率67.98%。属北温带大陆性气候, 水热条件良好, 冬暖夏凉, 雨量丰富, 年平均气温-11.6℃, 比平原高4℃, 极端最低气温-33.4℃, 7月平均气温15℃, 极端最高气温30.5℃, 年降水量500 mm, 积雪20 cm。土壤为亚高山草甸土、灰褐色森林土、栗钙土及灰钙土。根据研究区的植被生境分布和实际情况设计长达20 km的两条样带, 在每条样带上分别选择了草甸草原、针叶林、林间草地、针阔混交林、阔叶林、苗圃林、灌木林7种典型生境, 用GPS定位系统测量各样地的海拔和经纬度, 使用地温计测量每样地不同土层温度, 各生境具体情况见表 1。
| 生境类型 Habitat types |
地理位置 Location |
海拔/m Elevation |
土壤类型 Soil types |
主要植物类型 Dominant plant species |
|
| Ⅰ | 草甸草原 Meadow steppe |
43°49′06″—43°51′08″ 87°52′24″—87°53′41″ |
2350—3050 | 草甸土 | 羊茅(Festuca ovina L.), 羊草(Leymus chinensis Tzvel.), 嵩草属(Kobresia Willd), 早熟禾(Poa annua L.), 苔藓Bryophytes等 |
| Ⅱ | 针叶林 Coniferous forest |
43°47′22″—43°49′08″ 86°59′36″—87°14′44″ |
1900—2350 | 灰褐森林土 | 雪岭云杉(Picea schrenkiana Fischet Mey.), 冷杉(Abies fabri (Mast.) Craib), 新疆冷杉(Abies sibirica), 老鹳草(Geranium wilfordii Maxim), 地衣Lichens, 苔藓Bryophytes等 |
| Ⅲ | 林中草地 Grass glade |
43°46′09″—43°49′28″ 87°45′24″—87°46′11″ |
1850—2050 | 棕钙土 | 荨麻(Urtica fissa E. Pritz.), 糙苏(Phlomis L.), 唇香草(Ziziphora sp.), 早熟禾(Poa annua L.), 蒲公英(Taraxacum spp.)等 |
| Ⅳ | 针阔混交林 Conifer-broadleaf forest |
43°49′32″—43°51′28″ 87°57′34″—87°58′48″ |
1850—1950 | 灰褐森林土 | 雪岭云杉(Picea schrenkiana Fischet Mey.), 天山花楸(Sorbus tianschanica Rupr.), 天山桦(Betula tianschanica), 山杨(Populus davidiana), 早熟禾(Poa annua L.), 地衣Lichens, 苔藓Bryophytes |
| Ⅴ | 阔叶林 Broad-leaved forest |
43°47′12″—43°48′08″ 87°56′24″—87°56′41″ |
1720—1850 | 栗钙土 | 密叶杨(Populus talassica Kom.), 山杨(Populus davidiana), 柳树(Salix babylonica L.), 裂叶榆(Ulmus laciniata), 酸模(Rumex acetosa L.), 车前(Plantago asiatica L.), 蒲公英(Taraxacum sp.), 早熟禾(Poa annua L.), 地衣Lichens, 苔藓Bryophytes等 |
| Ⅵ | 苗圃林 Nursery garden |
43°46′02″—43°59′18″ 87°46′24″—87°46′41″ |
1710—1750 | 灰钙土 | 新疆冷杉(Abies sibirica), 西伯利亚落叶松(Larix sibirica Ledeb), 天山桦(Betula tianschanica), 野青茅(Deyeuxia arundinacea L.), 老鹳草(Geranium wilfordii Maxim), 地衣Lichens, 苔藓Bryophytes等 |
| Ⅶ | 灌木林 Shrubbery |
43°48′38″—43°49′45″ 87°59′24″—87°59′49″ |
1710—1950 | 灰钙土 | 侧柏(Platycladus orientalis L.), 小檗(Berbens sp.), 蔷薇(Rosa sp.), 忍冬(Lonicera japonica Thunb.), 刺芹(Eryngium foetidum), 早熟禾(Poa annua L.), 地衣Lichens, 苔藓Bryophytes等 |
分别在2014年4、7、9月及11月中旬对天山森林公园两条样带上选择了7种典型生境进行定点采样。每个生境内设置3个20 m×20 m的样方, 每个样方内随机选取5个样点, 每个样点间隔5 m左右, 在每样点内按对角线法采5个环刀样品, 环刀样品容积为100 cm3采样器, 分4个土层(0—5、5—10、10—15、15—20 cm)进行采样, 共采集560个样。同时在各样点采集一份混合土样待测土壤理化性质。将所有土样装入保鲜袋带回实验, 采用改良的Tullgren干漏斗法将中小型土壤动物从土壤样品中分离出来, 并制作装片。螨类标本依据尹文英、青木淳一、江原昭三、Balogh J和Balogh P以及Krantz GW和Walter DE等的分类检索方法进行分类鉴定, 一般鉴定到属级分类单元[20-24]。
1.3 主要环境因子的测定在调查土壤螨类的同时, 测定各采样点的主要环境因子(土壤容重、温度、湿度、pH、有机质、全氮、全磷、全钾和总盐含量)。土壤容重采用土壤环刀法;土壤温度和土壤酸碱分别使用地温计和土壤酸度计测定;土壤水分含量采用烘干法测定(GB 7172—1987), 按照以下公式计算含水率:含水率(%)=(G0-G2)(湿土重-干土重)/G2×100%。土壤有机质含量采用重铬酸钾-硫酸溶液氧化法(GB 9834—1988)测定。土壤全钾含量参照GB 9836—1988法测定;土壤全磷含量参照GB 9837—1988法测定;土壤全氮含量参照GB 7173—1987法测定;总盐量采用水溶性盐总量(质量法)测定(NY/T 1121.16—2006)[14, 30]。
1.4 数据处理对7种生境间土壤螨类群落组成和数量进行多样性分析[18]:
(1) Shannon-Wiener多样性指数(H):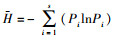
(2) Margalef丰富度指数(M):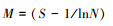
(3) Pielou均匀度指数(E):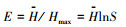
(4) Simpson优势度指数(C):
(5) Jacccard群落相似性系数(q):
式中, N为群落的总个体数, S代表土壤螨类的总类群数, Pi代表第i类土壤螨类的个体数与土壤螨类总个体数的比, ni为第i类群个体数。a为群落A含有的全部种类, b为群落B含有的全部种类, c为群落A与B的共有种类。采用Canoco 4.5软件分析土壤螨类群落物种重要值与环境变量的关系。在进行排序分析之前, 首先对物种矩阵做除趋势对应分析(Detrended Correspondence Analysis, DCA), 结果显示, 所有排序的最长轴长度小于3, 因此适合选用冗余分析(Redundancy Analysis, RDA)排序方法。考虑到环境因子的多样性可能导致排序结果的不可靠, 在进行RDA分析前, 计算了环境因子的膨胀系数(inflation factor, IFs)。结果表明, 所有环境因子的膨胀系数均小于10, 说明环境因子间的共线性十分微弱, 可以不予考虑。同时为了最小化稀有物种对结果的影响, 分析前删除了出现在样方数少于3个的物种。所有排序的显著性均由Monte Carlo随机置换(499次, P < 0.05)检验进行检验。为了最小化稀有物种对排序结果的影响, 对所有排序都执行了稀有物种的降权处理, 最后用Canoco Draw绘制RDA排序图。采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异, 单因素方差分析使用SPSS 19.0软件辅以Excel 2010实现。
2 结果与分析 2.1 土壤螨类群落组成和数量变化天山森林公园7种不同生境共捕获土壤螨类24399只成体螨类标本, 属于4目56科108属(包括9个中国新记录属, 见表 2), 其中甲螨亚目21814只, 40科81属, 占总数的89.4%;中气门亚目2131只, 10科16属, 占总数的8.73%;前气门亚目416只5科9属, 占总数的1.71%;无气门亚目38只, 1科2属, 占总数的0.16%, 见表 2。
| 属名 Genera |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | 总数 Total |
密度 Density |
百分比/% Percentage |
丰富度 Abundance |
| 古甲螨属Palaeacarus | 0 | 12 | 0 | 4 | 0 | 6 | 0 | 22 | 99.5±22 | 0.09 | + |
| 短甲螨属Brachychthonius | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 8 | 36.2±8 | 0.03 | + |
| 滑缝甲螨属Liochthonius | 0 | 16 | 0 | 0 | 32 | 6 | 0 | 54 | 244±54 | 0.22 | + |
| 长毛缝甲螨属Trichthonius | 30 | 20 | 0 | 0 | 10 | 18 | 6 | 84 | 379.8±84 | 0.34 | + |
| 三皱甲螨属Rhysotritia | 556 | 424 | 0 | 0 | 44 | 82 | 48 | 1154 | 5217±1154 | 4.73 | ++ |
| 霍卷甲螨属Hoplophthiracarus | 22 | 38 | 0 | 17 | 46 | 10 | 24 | 157 | 710±157 | 0.64 | + |
| 卷甲螨属Phthiracarus | 0 | 20 | 0 | 0 | 6 | 0 | 0 | 26 | 117.5±26 | 0.11 | + |
| 直卷甲螨属Archoplophora | 0 | 138 | 6 | 99 | 0 | 40 | 8 | 291 | 1316±291 | 1.19 | ++ |
| 上罗甲螨属Epilohmannia | 0 | 90 | 0 | 180 | 38 | 78 | 102 | 488 | 2206±488 | 2.00 | ++ |
| 盲甲螨属Malaconothrus | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 5 | 22.6±5 | 0.02 | + |
| 三盲甲螨属Trimalaconothrus | 0 | 0 | 0 | 4 | 0 | 0 | 13 | 17 | 76.9±17 | 0.07 | + |
| 懒甲螨属Nothrus | 82 | 100 | 14 | 51 | 108 | 66 | 20 | 441 | 1994±441 | 1.81 | ++ |
| 礼服甲螨属Trhypochthonius | 64 | 234 | 52 | 114 | 200 | 134 | 104 | 902 | 4078±902 | 3.70 | ++ |
| 异懒甲螨属Allonothrus | 0 | 4 | 0 | 45 | 0 | 0 | 0 | 49 | 221.5±49 | 0.20 | + |
| 洼甲螨属Camisia | 24 | 256 | 8 | 210 | 50 | 126 | 98 | 772 | 3490±772 | 3.16 | ++ |
| 平懒甲螨属Platynothrus | 30 | 0 | 0 | 0 | 0 | 14 | 0 | 44 | 198.9±44 | 0.18 | + |
| 沙甲螨属Eremulus | 0 | 116 | 0 | 21 | 0 | 24 | 0 | 161 | 728±161 | 0.66 | + |
| 沙足甲螨属Eremobelba | 0 | 8 | 0 | 0 | 8 | 12 | 0 | 28 | 126.6±28 | 0.11 | + |
| 下珠足甲螨属Hypodamaeus | 110 | 32 | 6 | 22 | 82 | 26 | 58 | 336 | 1519±336 | 1.38 | ++ |
| 珠足甲螨属Belba | 0 | 30 | 0 | 0 | 0 | 0 | 0 | 30 | 135.6±30 | 0.12 | + |
| 表珠甲螨属Epidamaeus | 44 | 166 | 8 | 66 | 80 | 18 | 38 | 420 | 1899±420 | 1.72 | ++ |
| 扇珠甲螨属Licnodamaeus | 0 | 12 | 0 | 0 | 56 | 12 | 0 | 80 | 361.7±80 | 0.33 | + |
| 龙骨足甲螨属Eremaeus | 0 | 200 | 0 | 0 | 0 | 28 | 0 | 228 | 1031±228 | 0.93 | + |
| Pleodamaeus Paschoal, 1983 | 54 | 22 | 0 | 0 | 18 | 96 | 0 | 190 | 859±190 | 0.78 | + |
| Plesiodamaeus Grandjean, l954 | 6 | 0 | 0 | 11 | 0 | 8 | 0 | 25 | 113±25 | 0.10 | + |
| Adrodamaeus Paschoal, 1983 | 0 | 4 | 0 | 0 | 0 | 0 | 4 | 8 | 36.2±8 | 0.03 | + |
| Odontodamaeus Paschoal, 1982 | 0 | 32 | 0 | 0 | 14 | 8 | 0 | 54 | 244.1±54 | 0.22 | + |
| Joshuella Wallwork, 1972 | 0 | 14 | 0 | 19 | 0 | 0 | 0 | 33 | 149.2±33 | 0.14 | + |
| Crotonia Thorell, 1876 | 0 | 46 | 0 | 23 | 0 | 0 | 0 | 69 | 311.9±69 | 0.28 | + |
| 平壳甲螨属Platyliodes | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 8 | 36.2±8 | 0.03 | + |
| 奥甲螨属Oppia | 0 | 64 | 0 | 21 | 16 | 0 | 0 | 101 | 456.6±101 | 0.41 | + |
| 多奥甲螨属Multioppia | 0 | 69 | 0 | 12 | 19 | 0 | 16 | 134 | 605.8±134 | 0.48 | + |
| 单奥甲螨属Phauloppia | 0 | 15 | 0 | 0 | 3 | 0 | 0 | 18 | 81.4±18 | 0.07 | + |
| 小奥甲螨属Oppiella | 0 | 1006 | 24 | 453 | 94 | 90 | 36 | 1703 | 7699±1703 | 6.98 | ++ |
| 四奥甲螨属Quadroppia | 0 | 90 | 8 | 240 | 86 | 10 | 22 | 456 | 2061.6±456 | 1.87 | ++ |
| 微奥甲螨属Microppia | 0 | 0 | 0 | 0 | 14 | 0 | 0 | 14 | 63.3±14 | 0.06 | + |
| Stachyoppia Balogh, 1961 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 9±2 | 0.01 | + |
| 大奥甲螨属Lasiobelba | 0 | 0 | 0 | 6 | 4 | 0 | 0 | 10 | 45.2±10 | 0.04 | + |
| 丽甲螨属Liacarus | 208 | 128 | 18 | 45 | 92 | 62 | 16 | 569 | 2572±569 | 2.33 | + |
| 步甲螨属Carabodes | 46 | 32 | 0 | 9 | 30 | 16 | 16 | 149 | 673.6±149 | 0.61 | + |
| 盖头甲螨属Tectocepheus | 32 | 300 | 16 | 99 | 330 | 48 | 76 | 901 | 4073±901 | 3.69 | ++ |
| 翼盖头甲螨属Tegeozetes | 4 | 4 | 0 | 0 | 0 | 0 | 0 | 8 | 36.2±8 | 0.03 | + |
| 细若甲螨属Incabates | 6 | 6 | 0 | 16 | 0 | 4 | 4 | 36 | 162.8±36 | 0.15 | + |
| 若甲螨属Oribatula | 0 | 106 | 0 | 0 | 22 | 40 | 106 | 274 | 1239±274 | 1.12 | ++ |
| 合若甲螨属Zygoribatula | 4 | 76 | 0 | 42 | 40 | 4 | 14 | 180 | 814±180 | 0.74 | + |
| 贪颈尖棱甲螨属Jugatala | 370 | 406 | 30 | 0 | 186 | 160 | 46 | 1198 | 5416±1198 | 4.91 | ++ |
| 菌板鳃甲螨属Mycobates | 18 | 454 | 34 | 29 | 0 | 240 | 100 | 875 | 3956±875 | 3.59 | ++ |
| 点肋甲螨属Punctoribates | 0 | 30 | 0 | 0 | 74 | 44 | 46 | 194 | 877±194 | 0.80 | + |
| 缰板鳃甲螨属Chamobates | 36 | 42 | 6 | 0 | 64 | 32 | 16 | 196 | 886±196 | 0.80 | + |
| 菌甲螨属Scheloribates | 0 | 201 | 0 | 99 | 256 | 18 | 81 | 655 | 2961±655 | 2.68 | ++ |
| 全菌甲螨属Perscheloribates | 0 | 49 | 0 | 12 | 0 | 0 | 19 | 80 | 361.7±80 | 0.33 | + |
| 刀肋甲螨属Cultroribula | 0 | 86 | 0 | 279 | 26 | 6 | 4 | 401 | 1813±401 | 1.64 | ++ |
| 足肋甲螨属Podoribates | 108 | 0 | 0 | 0 | 12 | 0 | 0 | 120 | 543±120 | 0.49 | + |
| 爪甲螨属Unguizetes | 100 | 155 | 0 | 0 | 68 | 0 | 30 | 353 | 1596±353 | 1.45 | ++ |
| 杆棱甲螨属Mochlozetes | 0 | 12 | 0 | 0 | 5 | 0 | 0 | 17 | 76.9±17 | 0.07 | + |
| 毛甲螨属Trichoribates | 0 | 22 | 0 | 0 | 0 | 0 | 0 | 22 | 99.5±22 | 0.09 | + |
| 裂头甲螨属Fissicepheus | 0 | 5 | 0 | 0 | 2 | 0 | 0 | 7 | 31.6±7 | 0.03 | + |
| 残领甲螨属Defectamerus | 0 | 4 | 0 | 0 | 1 | 0 | 0 | 5 | 22.6±5 | 0.02 | + |
| 佐渡藓甲螨属Sadocepheus | 10 | 0 | 0 | 0 | 8 | 0 | 4 | 22 | 99.5±22 | 0.09 | + |
| 小尖棱甲螨属Ceratozetella | 10 | 40 | 0 | 0 | 0 | 0 | 0 | 50 | 226.1±50 | 0.20 | + |
| 尖棱甲螨属Ceratozetes | 36 | 492 | 10 | 102 | 0 | 284 | 170 | 1092 | 4936±1092 | 4.48 | ++ |
| 肩翅尖棱甲螨属Diapterobates | 0 | 0 | 0 | 0 | 0 | 16 | 0 | 16 | 72.3±16 | 0.07 | + |
| 鳞顶甲螨属Lepidozetes | 0 | 312 | 54 | 60 | 320 | 84 | 30 | 860 | 3888±860 | 3.52 | ++ |
| 盾顶甲螨属Scutozetes | 0 | 37 | 82 | 13 | 58 | 30 | 48 | 268 | 1212±268 | 1.10 | ++ |
| 隐肋甲螨属Costeremus | 0 | 5 | 0 | 3 | 12 | 0 | 0 | 20 | 90.4±20 | 0.08 | + |
| 梁甲螨属Lamellobates | 0 | 20 | 0 | 0 | 0 | 0 | 0 | 20 | 90.4±20 | 0.08 | + |
| 木单翼甲螨属Xylobates | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 3 | 13.6±3 | 0.01 | + |
| 全单翼甲螨属Perxylobates | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 9±2 | 0.01 | + |
| 小甲螨属Oribatella | 238 | 450 | 224 | 939 | 284 | 120 | 200 | 2455 | 11099±2455 | 10.06 | +++ |
| 蛇轮甲螨属Ophidiotrichus | 0 | 59 | 0 | 0 | 0 | 0 | 24 | 83 | 375.2±83 | 0.34 | + |
| 足肋甲螨属Podoribates | 0 | 0 | 0 | 147 | 0 | 0 | 10 | 157 | 709±157 | 0.64 | + |
| 角翼甲螨属Achipteria | 0 | 34 | 0 | 0 | 47 | 0 | 0 | 86 | 388±86 | 0.33 | + |
| 圆单翼甲螨属Peloribates | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 5 | 22.6±5 | 0.02 | + |
| 瘤前翼甲螨属Peloptulus | 0 | 60 | 34 | 414 | 0 | 72 | 0 | 580 | 2622±580 | 2.38 | ++ |
| 角甲螨属Ceratoppia | 0 | 20 | 12 | 63 | 6 | 14 | 224 | 339 | 1532±339 | 1.39 | ++ |
| 真前翼甲螨属Eupelops | 24 | 186 | 16 | 0 | 178 | 46 | 14 | 464 | 2097±464 | 1.90 | ++ |
| 无前翼甲螨属Anachipteria | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1±4 | 0.02 | + |
| 原大翼甲螨属Protokalumma | 14 | 6 | 20 | 0 | 10 | 40 | 0 | 90 | 406.9±406 | 0.37 | + |
| 副大翼甲螨属Parakalumma | 0 | 18 | 0 | 3 | 0 | 0 | 2 | 23 | 104±23 | 0.09 | + |
| 大翼甲螨属Galumna | 0 | 86 | 0 | 90 | 8 | 16 | 16 | 216 | 976.5±216 | 0.89 | + |
| 全大翼甲螨属Pergalumna | 0 | 52 | 0 | 54 | 10 | 2 | 22 | 140 | 632.9±140 | 0.57 | + |
| 革赛螨属Gamasellus | 190 | 80 | 4 | 0 | 64 | 8 | 16 | 362 | 1636±362 | 1.48 | + |
| 上厉螨属Hyperlaelaps | 0 | 36 | 0 | 60 | 52 | 38 | 8 | 194 | 877.1±194 | 0.80 | + |
| 虫穴螨属Zercon | 0 | 84 | 338 | 48 | 0 | 38 | 12 | 520 | 2350±520 | 2.13 | ++ |
| 囊螨属Asca | 12 | 22 | 0 | 0 | 0 | 4 | 0 | 38 | 171.8±38 | 0.16 | + |
| 滑绥螨属Leioseius | 0 | 0 | 0 | 0 | 8 | 6 | 4 | 18 | 81.4±18 | 0.07 | + |
| 毛绥螨属Lasioseius | 0 | 4 | 34 | 0 | 0 | 0 | 0 | 38 | 171.8±38 | 0.16 | + |
| 美绥螨属Ameroseius | 0 | 74 | 2 | 25 | 14 | 58 | 8 | 181 | 818.3±181 | 0.74 | + |
| 新曲厉螨属Neocypholaelaps | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 4 | 18.1±4 | 0.02 | + |
| 钝绥螨属Amblyseius | 0 | 30 | 0 | 4 | 0 | 0 | 4 | 38 | 171.8±38 | 0.16 | + |
| 植绥螨属Phytoseius | 0 | 20 | 10 | 12 | 4 | 16 | 36 | 98 | 443±98 | 0.40 | + |
| 盲走螨属Typhlodromus | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 6 | 27.1±6 | 0.02 | + |
| Mixozercon Halašková, 1963 | 20 | 14 | 0 | 0 | 22 | 84 | 4 | 144 | 651±144 | 0.59 | + |
| 小全盾螨属Holostaspella | 0 | 84 | 42 | 168 | 6 | 96 | 38 | 434 | 1962±434 | 1.78 | ++ |
| 尾绥螨属Uroseius | 0 | 8 | 2 | 0 | 6 | 4 | 6 | 26 | 117.5±26 | 0.11 | + |
| 双革螨属Digamasellus | 0 | 0 | 0 | 4 | 0 | 6 | 0 | 10 | 45.2±10 | 0.04 | + |
| 新似巨须螨属Neocunaxoides | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1±4 | 0.02 | + |
| 巨须螨属Cunaxa | 0 | 48 | 10 | 180 | 14 | 0 | 0 | 252 | 1139±252 | 1.03 | ++ |
| 拟邦佐螨属 Pseudobonzia | 0 | 12 | 4 | 0 | 0 | 50 | 0 | 66 | 298.4±66 | 0.27 | + |
| Neomolgus Oudemans, 1937 | 0 | 8 | 0 | 5 | 0 | 0 | 0 | 13 | 58.8±13 | 0.05 | + |
| 管吸螨属Cyta | 2 | 4 | 0 | 0 | 0 | 10 | 0 | 16 | 72.3±16 | 0.07 | + |
| 土皮须螨属Ledermuelleria | 0 | 6 | 0 | 7 | 0 | 0 | 0 | 13 | 58.8±13 | 0.05 | + |
| 长须螨属Stigmaeus | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 10 | 45.2±10 | 0.04 | + |
| 扁螨属Pentamerismus | 3 | 0 | 0 | 0 | 0 | 10 | 0 | 13 | 58.8±13 | 0.05 | + |
| 隐爪螨属Nanorchestes | 0 | 0 | 0 | 21 | 8 | 0 | 0 | 29 | 131.1±29 | 0.12 | + |
| 苔螨属Bryobia | 0 | 3 | 0 | 0 | 0 | 2 | 0 | 5 | 22.6±5 | 0.02 | + |
| 嗜粉螨属Aleuroglyphus | 0 | 9 | 0 | 0 | 0 | 14 | 0 | 23 | 104±23 | 0.09 | + |
| 脂螨属Lardoglyphus | 0 | 6 | 0 | 0 | 4 | 0 | 0 | 10 | 45.2±10 | 0.04 | + |
| 个体数(N) Individuals | 2517 | 7846 | 1128 | 4670 | 3404 | 2756 | 2078 | 24399 | 110307±243 | 100.00 | |
| 类群数(S) Group number | 34 | 87 | 30 | 51 | 62 | 61 | 54 | 108 | |||
| +++:优势类群Dominant groups(>10%);++:常见类群Common groups(1%—10%);+:稀有类群Rare group(<1%) | |||||||||||
对整个研究区域来说, 小甲螨属Oribatella为优势类群, 占总个体数的10.09%;三皱甲螨属Rhysotritia、直卷甲螨属Archoplophora、上罗甲螨属Epilohmannia、懒甲螨属Nothrus和礼服甲螨属Trhypochthonius等28属为常见类群, 占总个体数的71.69%。古甲螨属Palaeacarus、短甲螨属Brachychthonius、滑缝甲螨属Liochthonius、长毛缝甲螨属Trichthonius及卷甲螨属Phthiracarus等79属为稀有类群, 占总数的18.22%。
七种不同生境土壤螨类个体数和类群数时空分布均有显著差异(P < 0.05)。其中个体数水平分布表现为针叶林>针阔混交林>阔叶林>苗圃林>草甸草原>灌木林>林中草地。类群数依次为针叶林>苗圃林>阔叶林>灌木林>针阔混交林>草甸草原>林中草地。
根据七种不同生境捕获土壤螨类个体数和类群数, 分别进行群落多样性、相似性及相关性比较分析。Shannon-Wiener多样性指数(H)针叶林最高, 而林中草地最低, 其顺序依次为针叶林>苗圃林>阔叶林>灌木林>针阔混交林>草甸草原>林中草地;Simpson优势度指数(C)林中草地最高, 苗圃林最低, 其顺序依次为林中草地>草甸草原>针阔混交林>灌木林>阔叶林>针叶林>苗圃林。相关性分析表明, 在七种不同生境之间土壤螨类群落多样性指标均有显著差异(P < 0.05), 但针叶林环境个别指标的差异不太明显, 见表 3。
| 生境类型 Habitat types |
个体数(N) Individual number |
类群数(S) Group number |
多样性指数(H) Shannon-Wiener |
丰富度指数(M) Margalef index |
优势度指数(C) Simpson index |
均匀度指数(E) Pielou index |
| Ⅰ | 2531.67±166.49d | 33.33±16.01b | 2.73±0.14c | 4.21±0.69d | 0.1±0.06ab | 0.83±0.07ab |
| Ⅱ | 7831.67±196.89a | 75.45±18.33a | 3.58±0.18a | 8.66±0.76a | 0.03±0.03b | 0.83±0.06ab |
| Ⅲ | 1132.22±165.04f | 29.33±16.01b | 2.52±0.13c | 4.1±0.64d | 0.14±0.07a | 0.79±0.06b |
| Ⅳ | 4666.67±214.02b | 48.33±23.01ab | 3.09±0.16b | 5.65±0.76c | 0.07±0.01b | 0.81±0.06ab |
| Ⅴ | 3519.12±101.12c | 57.33±18.5ab | 3.39±0.17a | 6.85±0.65bc | 0.05±0.0b | 0.85±0.06ab |
| Ⅵ | 2716.32±173.49d | 61.33±18.5ab | 3.56±0.18a | 7.54±0.75ab | 0.04±0.0b | 0.88±0.07a |
| Ⅶ | 2077.67±179.5e | 50.33±20.5ab | 3.35±0.17ab | 6.44±0.86bc | 0.05±0.01b | 0.75±0.07ab |
| 数据以平均值标准误形式表示, 不同小写字母代表不同生境间具有统计学显著差异, P < 0.05 | ||||||
同一个生境不同季节土壤螨类群落多样性特征存在显著差异(P < 0.05)。从不同指标看, 秋季多样性指数和丰富度指数在各生境中均最高, 而优势度指数最低, 显示出多样性指数和优势度指数呈负相关关系, 见图 1。

|
| 图 1 新疆天山森林公园不同生境土壤螨类多样性特征指数季节动态 Fig. 1 Diversity indices of soil mites in different habitats of Tianshan forest park of Xinjiang |
根据各生境种类组成的相似程度, 计算出不同生境间的群落相似性系数, 结果见表 4。
| 生境类型 Habitat types |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ |
| Ⅰ | 1.0000 | ||||||
| Ⅱ | 0.3294 | 1.0000 | |||||
| Ⅲ | 0.3061 | 0.3457 | 1.0000 | ||||
| Ⅳ | 0.2206 | 0.5059 | 0.3860 | 1.0000 | |||
| Ⅴ | 0.3382 | 0.5281 | 0.3594 | 0.3766 | 1.0000 | ||
| Ⅵ | 0.4179 | 0.6279 | 0.4219 | 0.4667 | 0.5526 | 1.0000 | |
| Ⅶ | 0.3770 | 0.4659 | 0.4545 | 0.5231 | 0.5507 | 0.5857 | 1.0000 |
| 当0 < q < 0.25时, 极不相似significantly dissimilar;当0.25≤q < 0.50时, 中等不相似moderately dissimilar;当0.50≤q < 0.75时, 中等相似moderately similar;当0.75≤q < 1.00时, 极相似significantly similar | |||||||
由表 4可见, 7种不同生境土壤螨类群落间的Jaccard相似性系数具有较大的差异, 针叶林和苗圃林之间相似性系数最大(0.6279), 草甸草原和针阔混交林之间相似性系数最低(0.2206)。由于针叶林和苗圃林两种生境的植被均为高大乔木, 具有较厚的凋落物层, 并土壤条件类似, 而草甸草原和针阔混交林这两种生境的植被、土壤环境及人为干扰等差异较大。所有系数中有14个系数在0.25和0.50之间表现为中等不相似, 而7个系数在0.50和0.75之间表现为中等相似。
2.2 土壤螨类群落特征与环境因子的关系环境因子相关性分析表明, 在七种不同生境土壤环境因子中除了土壤温度和土壤pH以外其他环境因素之间均有显著差异(P < 0.05), 结果见表 5。
| 生境类型 Habitat types |
海拔 Altitude/m | 土壤温度 Soil temperature/ ℃ | 土壤含水量 Soil water content/% | 土壤容重 Bulk density/ (g/m3) | 土壤pH Soil pH | 土壤有机质 Soil organic matter/ (g/kg) | 土壤全氮 Total nitrogen/ (g/kg) | 土壤全磷 Total phosphorus/ (g/kg) | 土壤全钾 Total potassium/ (g/kg) | 土壤总盐量 Total salt content/ (g/kg) |
| Ⅰ | 1712.50±12.58g | 18.08±10.91a | 18.60±1.97e | 1.15±1.13a | 7.59±0.34a | 17.45±2.07cd | 1.74±0.15d | 1.10±0.17b | 17.37±0.58ab | 1.35±0.33b |
| Ⅱ | 2317.50±28.72a | 9.00±9.61a | 36.95±3.04a | 0.88±0.04b | 7.59±0.07a | 28.25±3.16a | 3.58±0.30a | 1.52±0.10a | 21.27±0.13a | 2.18±0.10a |
| Ⅲ | 1747.00±2.45f | 16.25±10.78a | 26.33±1.30d | 1.10±0.09a | 7.51±0.16a | 20.37±2.79cd | 2.29±0.43cd | 1.09±0.08b | 20.04±1.29ab | 1.23±0.10b |
| Ⅳ | 2151.25±2.50b | 12.48±9.54a | 33.80±2.34ab | 1.08±0.05a | 7.45±0.08a | 25.42±2.32ab | 2.85±0.42b | 1.39±0.06a | 21.24±2.97a | 2.03±0.18a |
| Ⅴ | 2022.5±20.21c | 13.30±10.17a | 30.23±4.43bcd | 1.12±0.06a | 7.31±0.11a | 21.38±3.14bc | 2.53±0.48bc | 1.08±0.22b | 19.11±3.20ab | 1.45±0.14b |
| Ⅵ | 1958.75±6.29d | 13.10±9.39a | 32.25±2.91bc | 1.13±0.01a | 7.44±0.22a | 19.45±3.28cd | 2.08±0.28cd | 0.97±0.17b | 16.79±1.61b | 1.38±0.05b |
| Ⅶ | 1861.25±7.50e | 15.20±10.36a | 28.60±2.64cd | 1.05±0.09a | 7.43±0.23a | 16.81±2.42d | 1.97±0.25d | 1.02±0.05b | 18.70±5.03ab | 1.32±0.10b |
为探讨不同生境土壤环境因素对土壤螨类群落多样性特征的影响, 采用One-way ANOVA和Pearson相关性分析方法进行分析, 相关分析结果见表 6。
| 环境因素 Environmental factors |
个体数(N) Individual number |
类群数(S) Group number | 多样性指数(H) Shannon- Wiener index | 丰富度指数(M) Margalef index |
优势度指数(C) Simpson index |
均匀度指数(E) Pielou index |
| 海拔Altitude/m | 0.893** | 0.714* | 0.693* | 0.790* | -0.607 | 0.321 |
| 土壤温度Soil temperature/℃ | -0.857** | -0.750* | -0.752* | -0.870** | 0.679* | -0.393* |
| 土壤含水量Soil Water Content/% | 0.857** | 0.750* | 0.723* | 0.808* | -0.679* | 0.393* |
| 土壤容重Bulk density/(g/m3) | -0.321 | -0.321 | -0.457* | -0.638 | 0.143 | -0.071 |
| 土壤pH Soil pH | 0.179 | -0.107 | -0.315 | -0.153 | 0.250 | -0.607 |
| 土壤有机质Soil organic matter/(g/kg) | 0.786** | 0.357* | 0.321** | -0.504* | -0.214 | -0.214 |
| 土壤全氮Total nitrogen/(g/kg) | 0.750* | 0.429 | 0.413* | 0.602 | -0.286 | -0.071 |
| 土壤全磷Total phosphorus/(g/kg) | 0.536 | -0.071 | 0.168 | 0.337 | 0.286 | -0.643 |
| 土壤全钾Total potassium/(g/kg) | 0.107 | 0.071 | 0.187 | 0.062 | -0.055 | -0.036 |
| 土壤总盐量Total salt content/(g/kg) | 0.999** | 0.679* | 0.413 | 0.53 | -0.582 | 0.214 |
| *P < 0.05, **P < 0.01 | ||||||
由表 6可知, 螨类类群数与海拔、土壤温度、湿度和土壤有机质存在显著的相关关系(P < 0.05);多样性指数(H)与海拔高度、土壤温度、湿度、容重、有机质和全氮含量存在显著的相关关系(P < 0.05), 其中多样性指数(H)与有机质含量呈极显著的相关关系(P < 0.01);丰富度指数(M)与海拔高度、土壤温度、湿度、容重和有机质含量存在显著的相关关系(P < 0.05);优势度指数(C)与土壤温度和湿度存在显著的相关关系(P < 0.05), 均匀性指数与土壤温度和湿度存在显著的负相关关系(P < 0.05)。
为进一步探讨不同生境土壤理化性质对土壤螨类群落的影响, 采用冗余分析(RDA)方法进行分析, 分析结果见图 2和3及表 5和6。

|
| 图 2 新疆天山森林公园不同生境样地与环境因子的RDA二维排序图 Fig. 2 Biplot between different habitats and environmental factors in Tianshan forest park of Xinjiang Altitude:海拔Altitude;ST:土壤温度Soil temperature;SWC:土壤含水量Soil Water Content;BD:容重Bulk density;pH:土壤pH;SOM:土壤有机质Soil organic matter;TN:全氮Total nitrogen;TP:全磷Total phosphorus;TK:土壤全钾Total potassium;TSC:总盐量Total salt content |

|
| 图 3 新疆天山森林公园土壤螨类群落和环境因子的RDA排序图 Fig. 3 Biplot between soil mite communities and environmental factors in Tianshan forest park of Xinjiang Orib:小甲螨属Oribatella;Oppi:小奥甲螨属Oppiella;Quad:四奥甲螨属Quadroppia;Rhys:三皱甲螨属Rhysotritia;Cera:尖棱甲螨属Ceratozetes;Trhy:礼服甲螨属Trhypochthonius;Myco:菌板鳃甲螨属Mycobates;Sche:菌甲螨属Scheloribates;Tect:盖头甲螨属Tectocepheus;Cami:洼甲螨属Camisia;Lepi:鳞顶甲螨属Lepidozetes;Liac:丽甲螨属Liacarus;Pelo:瘤前翼甲螨属Peloptulus;Zerc:虫穴螨属Zercon;Noth:懒甲螨属Nothrus;Hypo:下珠足甲螨属Hypodamaeus;Epid:表珠甲螨属Epidamaeus;Oriba:若甲螨属Oribatula;Cult:刀肋甲螨属Cultroribula:Ungu:爪甲螨属Unguizetes;Arch:直卷甲螨属Archoplophora;Juga:贪颈尖棱甲螨属Jugatala;Cerat:角甲螨属Ceratoppia;Eupe:真前翼甲螨属Eupelops;Epil:上罗甲螨属Epilohmannia;Gama:革赛螨属Gamasellus;Holo:全盾螨属Holostaspella;Cuna:巨须螨属Cunaxa;Erem:龙骨足甲螨属Eremaeus;Punc:点肋甲螨属Punctoribates |
从排序图 2可知, 28个样地土壤螨类群落的相对位置, 其中17个样地位于第2排序轴的左侧, 这些样地与环境因子土壤有机质、含水量、温度和全氮含量相对应, 表征了这些样地土壤容重、全磷、全钾、pH值、海拔和全盐量较低的特征;另外11个样地位于第二排序轴的右侧, 与土壤环境因子土壤容重、全磷、全钾、pH值、海拔和全盐量相对应, 表征了这些样地土壤有机质、含水量、温度和全氮含量较低的特征。
由图 3和表 7可知, 所有环境因子共解释了土壤螨类群落物种组成变异的82.1%。排序轴Axis 1解释了34.8%的生境变化, 与土壤环境因子之间的相关系数为0.909, 即解释了土壤螨类群落与环境之间关系的34.8%。排序轴Axis 2进一步解释了7.5%的生境变化, 与土壤环境因子之间的相关系数为0.897, 解释了土壤螨类群落与环境之间关系的27.3%。排序轴Axis 1和排序轴Axis 2共同解释了62.1%的土壤螨类群落与土壤环境之间的关系。蒙特卡罗置换检验显示, 十种环境因子与全部排序轴均存在极显著的相关性(F =7.355, P =0.002)。物种组成累积解释量和物种-环境关系累积解释量分别达82.0%和99.5%, 说明排序效果良好。其中排序轴1主要反映了容重和全钾、全磷、pH对土壤螨类分布的影响, 排序轴2主要反映了、土壤温度、全氮、有机质、含水量对土壤螨类分布的影响。
| 环境因子及排序摘要 Environmental factors and ordination summary |
排序轴1 Axis 1 |
排序轴2 Axis 2 |
排序轴3 Axis 3 |
排序轴4 Axis 4 |
|
| 环境因子 Environmental variables |
海拔Altitude | 0.2799** | 0.5058 | 0.5471 | -0.5597 |
| 土壤温度ST Soil temperature | 0.0872** | -0.1474** | -0.1168** | 0.0648** | |
| RDA排序摘要 Ordination summary |
含水量SWC Soil water content | 0.5288 | -0.1518** | 0.0162 | -0.4994 |
| 容重BD Bulk density | -0.6699** | 0.5143 | 0.2263 | 0.1800 | |
| 土壤pH Soil pH | -0.0321** | 0.3116 | 0.1086 | 0.4646 | |
| 有机质SOM Soil organic matter | 0.8062** | -0.1074** | 0.1200** | -0.2046** | |
| 全氮TN Total nitroge | 0.6639** | -0.0652 | -0.1627** | -0.5105 | |
| 全磷TP Total phophorus | -0.1394** | 0.5810 | 0.1441 | -0.2632** | |
| 全钾TK Total potassium | 0.0091** | 0.7995 | -0.1157** | -0.1555 | |
| 总盐量TSC Total salt content | 0.0871** | 0.6766 | -0.4644 | -0.4593** | |
| 特征值Eigenvalues | 0.348 | 0.273 | 0.125 | 0.075 | |
| 物种-环境相关性 Species-environment correlations |
0.909 | 0.897 | 0.784 | 0.755 | |
| 物种累积百分比方差 Cumulative percentage variance of species data |
34.8 | 62.0 | 74.5 | 82.0 | |
| 物种-环境关系方差的累积百分比 Cumulative percentage variance of species- environment relation |
52.6 | 66.5 | 80.1 | 99.5 | |
| 所有典范轴的显著性测验 Significance test for all canonical ordination axes |
F=5.355 | P=0.0020 | |||
研究表明, 不同环境因素对土壤螨类群落多样性的影响具有一定的差异。土壤螨类群落的变化与海拔、湿度、有机质和全氮含量极显著相关(P < 0.01), 其中土壤湿度对土壤螨类的变化影响最大。
从排序图 3可知, 土壤螨类主要类群与环境因子之间的相互关系, 不同类群受环境因子的影响程度明显不同。其中下珠足甲螨属Hypodamaeus、贪颈尖棱甲螨属Jugatala、三皱甲螨属Rhysotritia、龙骨足甲螨属Eremaeus和菌板鳃甲螨属Mycobates受全磷、全氮、有机质含量及pH的影响较大, 而巨须螨属Cunaxa、刀肋甲螨属Cultroribula、四奥甲螨属Quadroppia、瘤前翼甲螨属Peloptulus、盖头甲螨属Tectocepheus、鳞顶甲螨属Lepidozetes、小甲螨属Oribatella、菌甲螨属Scheloribates、小奥甲螨属Oppiella、直卷甲螨属Archoplophora则受全磷、有机质、全氮含量及pH的负面影响较大;爪甲螨属Unguizetes、丽甲螨属Liacarus和革赛螨属Gamasellus受盐总量的影响较大;懒甲螨属Nothrus主要受海拔的影响;表珠甲螨属Epidamaeus和真前翼甲螨属Eupelops受全钾和含水量的影响;虫穴螨属Zercon、点肋甲螨属Punctoribates、全盾螨属Holostaspella、上罗甲螨属Epilohmannia、洼甲螨属Camisia、礼服甲螨属Trhypochthonius受容重和土壤温度的影响较大;角甲螨属Ceratoppia和若甲螨属Oribatula受全钾含量的负面影响。排序图上的距离小, 说明它们在样方中的分布差异较小。
3 讨论 3.1 土壤螨类群落结构特征土壤螨类群落组成和结构特征在很大程度上受区域气候和小环境的影响[12-14, 25]。天山森林公园土壤螨类资源丰富, 现已鉴定出56科108个属, 其中发现了9个中国新记录属(Pleodamaeus Paschoal, 1983、Plesiodamaeus Grandjean, 1954、Adrodamaeus Paschoal, 1983、Odontodamaeus Paschoal, 1982、Joshuella Wallwork, 1972、Crotonia Thorell, 1876、Neomolgus Oudemans, 1937、Mixozercon Halašková, 1963、Stachyoppia Balogh, 1961)。整个研究区域来说Oribatella属为优势类群, 在不同生境土壤螨类个体数及类群数之间均存在显著差异(P < 0.05), 这与我国其他区域的一些研究结果基本一致[26-34], 但存在一定的差异。Maribie等对不同土地利用类型下土壤螨类群落研究表明菌甲螨属Scheloribates和奥甲螨属Oppia为优势类群[1, 16]。九华山土壤螨类的优势类群为木单翼甲螨属Xylobates, 菌甲螨属Scheloribates和大翼甲螨属Galumna [26]、乌鲁木齐南部山区土壤螨类中小奥甲螨属Oppiella为优势类群[29];新疆喀纳斯国家级自然保护区土壤螨类群落中甲螨属Oppia, 盖头甲螨属Tectocepheus, 钉棱甲螨属Passalozetes和菌甲螨属Scheloribates为优势类群[30]。已有研究表明, 较强的人为干扰会减少稀有类群和特有类群数量, 而对优势类群数量影响不大[25-28]。环境条件的空间差异对土壤螨类群落具有很大的影响, 生境的分异构成了螨类群落分化的基础, 环境的差异与螨类的适应协同作用形成了各个与一定环境相联系, 具有一定内部结构的螨类群落[27-32]。
大量研究表明土壤螨类群落结构和多样性与林地植被成分密切相关, 凋落物为林地土壤生态系统有机质物质的主要来源, 其数量和性质可能对土壤螨类群落产生决定性的影响[33-35]。温度和湿度也是影响土壤螨类群落及多样性的最重要因子, 其种类和数量与土壤温度和有机质存在显著的正相关关系[37-38], 相对较高的地表温度有利于凋落物的分解, 而凋落物分解越快, 土壤螨类获得食物资源越多, 从而使得土壤螨类群落多样性升高[40]。
研究区域七种不同生境土壤螨类群落多样性均有显著差异, 此种差异与土壤环境的利用方式、水热条件, 植被类型以及凋落物层有关[39], 针叶林和苗圃林生境的凋落物较厚, 土壤有机质含量高, 人为干扰相对较少, 因此个体数和类群数多, 群落多样性及丰富度均较高。林中草地生境的多样性最低, 这可能与该生境凋落物较少且人为干扰较大有关。分析表明, 群落内的物种越丰富, 其多样性就越高, 同时多样性指数也受均匀度的影响, 群落内物种分布越均匀, 则优势度越小, 相反群落内物种分布不均匀, 则优势度就越大[39-40]。相似性指数是衡量生境与土壤螨类关系的一个重要指标, 七种生境类型之间土壤螨类群落相似性属于中等不相似, 说明该区域所选生境类型之间差异较大、代表性强, 因而具有实际研究意义。由于土壤螨类对微环境的细微变化特别敏感, 随着土壤环境变化其群落结构及多样性指标都发生较大的变动, 故其群落多样性特征可作为检测环境质量优劣的良好指示指标[39-40]。
3.2 土壤螨类群落与环境因子的相关性土壤螨类在一定的环境条件下生存和繁衍, 因此环境因子与土壤螨类的关系很密切。同一环境因子在不同生境对其土壤螨类有不同的作用和体现, 不同环境因子在同一生境对其土壤螨类也有不同的影响[30, 34]。本研究表明土壤螨类群落多样性与土壤全氮、全磷、有机质及海拔之间的关联作用大, 其中有机质和全氮含量呈正相关关系, 而土壤pH呈负相关关系, 并各生境有机质及全氮含量的变化趋势与土壤螨类群落多样性动态具有一致性, 这与国内外研究结果基本一致[32-34, 40]。研究表明, 植被结构通过改变微气候环境和土壤理化性质影响土壤螨类的种群密度。土壤理化性质中, 土壤容重、水分、养分元素, 甚至是重金属含量均是影响土壤螨类分布的重要因素[13, 41]。螨类密度和Shannon-Wiener多样性指数与土壤pH、湿度、土壤温度呈显著正相关, 而均匀度指数与土壤全钾呈显著正相关, 与土壤全氮呈显著负相关;优势度指数与土壤全氮呈显著正相关。回归分析表明, 土壤有机质、全氮、湿度和海拔是影响土壤螨类群落多样性的主要因素[41-43]。相关分析表明, 土壤螨类群落与环境因子之间的关系是海拔、温度、降水量、凋落物以及人为干扰等诸多因素的综合作用可以通过改变土壤理化性质而对土壤螨类群落多样性产生影响[44-45]。
4 结论(1) 新疆天山森林公土壤螨类资源丰富, 已鉴定出56科108属, 其中9属为中国新纪录属。土壤螨类群落多样性较高, 并在不同生境间螨类的个体数、类群数和多样性指数均存在着显著的差异性, 同一个生境不同季节土壤螨类群落多样性特征存在显著差异(P < 0.05)。
(2) RDA分析表明, 土壤螨类生态分布与环境因子的关系密切, 十种环境因子与全部排序轴间均存在极显著的相关性, 其中海拔、土壤含水量和土壤有机质对土壤螨类群落分布的影响最大。排序轴Axis 1和排序轴Axis 2共同解释了62.1%的土壤螨类群落与土壤环境之间的关系, 物种组成累积解释量和物种-环境关系累积解释量分别达82.0%和99.5%。
| [1] |
Maribie C W, Nyamasyo G H N, Ndegwa P N, Mung'atu J K, Lagerlöf J, Gikungu M. Abundance and diversity of soil mites (Acari) along a gradient of land use types in Taita Taveta, Kenya. Tropical and Subtropical Agroecosystems, 2011, 13(1): 11-26. |
| [2] |
Gormsen D, Hedlund K, Wang H F. Diversity of soil mite communities when managing plant communities on set-aside arable land. Applied Soil Ecology, 2006, 31(1/2): 147-158. |
| [3] |
Bedano J C, Cantú M P, Doucet M E. Influence of three different land management practices on soil mite (Arachnida:Acari) densities in relation to a natural soil. Applied Soil Ecology, 2006, 32(3): 293-304. DOI:10.1016/j.apsoil.2005.07.009 |
| [4] |
Gulvik M. Mites (Acari) as indicators of soil biodiversity and land use monitoring:a review. Polish Journal of Ecology, 2007, 55(3): 415-440. |
| [5] |
Badejo M A, Ola-Adams BA. Abundance and diversity of soil mites of fragmented habitats in a Biosphere reserve in southern Nigeria. Pesquisa Agropecuária Brasileira, 2000, 35(11): 2121-2128. DOI:10.1590/S0100-204X2000001100001 |
| [6] |
Wu D H, Zhang B, Bu Z Y, Chen P. The community characteristics of soil mites under different land uses in Changchun metropolitan area, China. Acta Ecologica Sinica, 2006, 26(1): 16-25. DOI:10.1016/S1872-2032(06)60002-4 |
| [7] |
Yin X Q, Li X Q, An J C, Wang F B. Characteristics of ecological distribution of soil microarthropod communities in the Wetlands of the Lhasa river on the Qinghai-Tibet Plateau. Wetlands, 2015, 35(3): 589-596. DOI:10.1007/s13157-015-0649-z |
| [8] |
N'Dri J K, Hance T, André H M, Lagerlöf J, Tondoh J E. Microarthropod use as bioindicators of the environmental state:case of soil mites (Acari) from Côte d'Ivoire. Journal of Animal & Plant Sciences, 2016, 29(2): 4622-4637. |
| [9] |
Lin L, Gao M X, Liu D, Zhang X P, Wu H T, Wu D H. Co-occurrence patterns of above-ground and below-ground mite communities in farmland of Sanjiang Plain, Northeast China. Chinese Geographical Science, 2014, 24(3): 339-347. DOI:10.1007/s11769-014-0683-4 |
| [10] |
郑经鸿, 王丽真, 张兴亚, 王新华, 薄新文. 新疆草地甲螨分布规律研究. 生态学报, 1993, 13(2): 171-176. DOI:10.3321/j.issn:1000-0933.1993.02.008 |
| [11] |
Zhang Y, Jin D C, Zhou Y F, Yang G P, Liang W Q. Community composition and diversity of soil oribatid mites(Acari:Oribatida) in karst forests in Shibing, Guizhou, Southwestern China. Acta Entomologica Sinica, 2015, 58(7): 791-799. |
| [12] |
Illig J, Norton R A, Scheu S, Maraun M. Density and community structure of soil-and bark-dwelling microarthropods along an altitudinal gradient in a tropical montane rainforest. Experimental and Applied Acarology, 2010, 52(1): 49-62. DOI:10.1007/s10493-010-9348-x |
| [13] |
高梅香, 刘冬, 张雪萍, 吴东辉. 三江平原农田地表和地下土壤螨类丰富度与环境因子的空间关联性. 生态学报, 2016, 36(6): 1782-1792. |
| [14] |
Omar A, Ajar Q, Mukaram H, Wu S L. Community diversity and its seasonal dynamics of soil mites in Oasis of the Sangong river watershed of Xinjiang, China. Journal of Xinjiang University:Natural Science Edition, 2016, 33(1): 6-19. |
| [15] |
Gao M X, Liu D, Lin L, Wu D H. The small-scale structure of a soil mite metacommunity. European Journal of Soil Biology, 2016, 74: 69-75. DOI:10.1016/j.ejsobi.2016.03.004 |
| [16] |
Bolger T, Arroyo J, Kenny J, Caplice M. Hierarchical analysis of mite community structures in Irish forests——A study of the relative contribution of location, forest type and microhabitat. Applied Soil Ecology, 2014, 83: 39-43. DOI:10.1016/j.apsoil.2013.06.004 |
| [17] |
Minor M A, Ermilov S G. Effects of topography on soil and litter mites (Acari:Oribatida, Mesostigmata) in a tropical monsoon forest in Southern Vietnam. Experimental and Applied Acarology, 2015, 67(3): 357-372. DOI:10.1007/s10493-015-9955-7 |
| [18] |
Magurran A E. Ecological Diversity and Its Measurement. New Jersey:Princeton University Press, 1988, 1-179. |
| [19] |
殷秀琴, 宋博, 董炜华, 辛未冬. 我国土壤动物生态地理研究进展. 地理学报, 2010, 65(1): 91-102. |
| [20] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998: 163-242, 527-562.
|
| [21] |
青木淳一. 日本土壤动物检索图鉴. 东京: 东海大学出版社, 1991: 173-436.
|
| [22] |
江原昭三. 日本蜱螨类检索图鉴. 东京: 全国农村教育协会出版社, 1980: 1-562.
|
| [23] |
Balogh J, Balogh P. The Oribatid Mites Genera of the World(Vol. 1 and 2). Budapest: The Hungarian National Museum Press, 1992: 1-263.
|
| [24] |
Krantz G W, Walter D E. A Manual of Acarology. 3rd ed. Texas: Texas Tech University Press, 2009: 1-806.
|
| [25] |
Kardol P, Reynolds W N, Norby R J, Classen A T. Climate change effects on soil microarthropod abundance and community structure. Applied Soil Ecology, 2011, 47(1): 37-44. DOI:10.1016/j.apsoil.2010.11.001 |
| [26] |
王宗英, 路有成, 王慧芙. 九华山土壤螨类的生态分布. 生态学报, 1996, 16(1): 58-64. |
| [27] |
吾玛尔·阿布力孜, 阿布都如苏力·吐逊, 木卡热木·阿吉木, 张卫红. 乌鲁木齐地区不同生境土壤捕食性螨类群落结构及其多样性研究. 新疆农业科学, 2012, 49(9): 1748-1758. |
| [28] |
吾玛尔·阿布力孜, 阿布都如苏力·吐逊, 木开热木·阿吉木, 吴松林. 准噶尔盆地东南缘绿洲-荒漠交错带土壤螨类的群落多样性与相似性. 干旱区资源与环境, 2013, 27(6): 75-79. |
| [29] |
吾玛尔·阿布力孜, 阿提坎木·吾布力喀斯木, 祖胡玛尔·阿里. 乌鲁木齐南山小渠子土壤螨类群落结构研究. 安徽农业科学, 2014, 42(24): 8191-8197. |
| [30] |
吾玛尔·阿布力孜, 阿提坎木·吾布力喀斯木, 吴松林. 新疆喀纳斯国家级自然保护区土壤螨类群落特征. 干旱区资源与环境, 2015, 29(2): 99-104. |
| [31] |
阿布都如苏力·吐逊, 吾玛尔·阿布力孜, 木开热木·阿吉木. 准噶尔盆地东南缘绿洲-荒漠交错带土壤螨类群落特征初步研究. 干旱区资源与环境, 2013, 27(3): 160-166. |
| [32] |
排孜丽耶·合力力, 吾玛尔·阿布力孜, 阿加尔·恰肯. 干旱区不同土地利用类型下土壤螨类群落多样性比较. 新疆农业科学, 2017, 54(12): 2267-2281. |
| [33] |
阿加尔·恰肯, 吾玛尔·阿布力孜, 排孜力耶·合力力. 新疆玛纳斯河流域土壤螨类群落多样性. 生态学报, 2017, 37(24): 8385-8396. |
| [34] |
Dirilgen T, Arroyo J, Dimmers W J, Faber J, Stone D, da Silva P M, Carvalho F, Schmelz R, Griffiths B S, Francisco R, Creamer R E, Sousa J P, Bolger T. Mite community composition across a European transect and its relationships to variation in other components of soil biodiversity. Applied Soil Ecology, 2016, 97: 86-97. DOI:10.1016/j.apsoil.2015.06.008 |
| [35] |
Alatalo J M, Jägerbrand A K, Juhanson J, Michelsen A, Łuptáčik P. Impacts of twenty years of experimental warming on soil carbon, nitrogen, moisture and soil mites across alpine/subarctic tundra communities. Scientific Reports, 2017, 7: 44489. DOI:10.1038/srep44489 |
| [36] |
韩雪梅, 李丹丹, 梁子安, 陈云峰, 胡诚. 北方常见农业土地利用方式对土壤螨群落结构的影响. 生态学报, 2013, 33(16): 5026-5034. |
| [37] |
张丽梅, 高梅香, 刘冬, 张雪萍, 吴东辉. 环境筛选和扩散限制在地表和地下螨群落物种共存中的调控作用. 生态学报, 2016, 36(13): 3951-3959. |
| [38] |
N'Dri J K, Seka F A, Pokou P K, N'Da R A G, Lagerlöf J. Abundance and diversity of soil mite (Acari) communities after conversion of tropical secondary forest into rubber plantations in Grand-Lahou, Côte d'Ivoire. Ecological Research, 2017, 32(6): 909-919. DOI:10.1007/s11284-017-1499-3 |
| [39] |
Alatalo J M, Jägerbrand A K, Molau U. Impacts of different climate change regimes and extreme climatic events on an alpine meadow community. Scientific Reports, 2016, 6: 21720. DOI:10.1038/srep21720 |
| [40] |
Zaitsev A S, Chauvat M, Wolters V. Spruce forest conversion to a mixed beech-coniferous stand modifies oribatid community structure. Applied Soil Ecology, 2014, 76: 60-67. DOI:10.1016/j.apsoil.2013.12.009 |
| [41] |
Fischer B M, Schatz H. Biodiversity of oribatid mites (Acari:Oribatida) along an altitudinal gradient in the Central Alps. Zootaxa, 2013, 3626(4): 429-454. DOI:10.11646/zootaxa.3626.4 |
| [42] |
Minor M A, Ermilov S G, Anichkin A E. Biodiversity of soil oribatid mites (Acari:Oribatida) in a tropical highland plateaux, Bi Doup-Nui Ba National Park, Southern Vietnam. Tropical Ecology, 2017, 58(1): 45-55. |
| [43] |
Gao M X, He P, Zhang X P, Liu D, Wu D H. Relative roles of spatial factors, environmental filtering and biotic interactions in fine-scale structuring of a soil mite community. Soil Biology and Biochemistry, 2014, 79: 68-77. DOI:10.1016/j.soilbio.2014.09.003 |
| [44] |
Kamczyc J, Urbanowski C, Pers-Kamczyc E. Mite communities (Acari:Mesostigmata) in young and mature coniferous forests after surface wildfire. Experimental and Applied Acarology, 2017, 72(2): 145-160. DOI:10.1007/s10493-017-0148-4 |
| [45] |
Minor M A, Babenko A B, Ermilov S G. Oribatid mites (Acari:Oribatida) and springtails (Collembola) in alpine habitats of southern New Zealand. New Zealand Journal of Zoology, 2017, 44(1): 65-85. DOI:10.1080/03014223.2016.1251950 |