 2019, Vol. 39
2019, Vol. 39文章信息
- 陈丽, 王东波, 君珊
- CHEN Li, WANG Dongbo, JUN Shan
- 拉萨河流域大型底栖动物群落结构及其与环境因子的关系
- Macroinvertebrate community structure and relationships with environmental factors in the Lhasa River Basin
- 生态学报. 2019, 39(3): 757-769
- Acta Ecologica Sinica. 2019, 39(3): 757-769
- http://dx.doi.org/10.5846/stxb201806011224
-
文章历史
- 收稿日期: 2018-06-01
- 修订日期: 2018-09-27





2. 呼伦贝尔市环境监测中心站, 呼伦贝尔 021008
2. Environmental Monitoring Center of Inner Mongolia, Hulunbeir 021008, China
用于水体生态健康评价的生物类群有细菌、藻类、底栖动物及鱼类, 其中底栖动物因其特有的生物属性, 是应用最为广泛的指示生物[1-2]。大型底栖动物是河流生态系统的重要组成生物, 它们迁移能力有限, 容易受外界环境条件的影响, 类群中有对环境敏感的物种, 也有耐污的物种, 通过对底栖动物特征的分析可以从不同侧面反映出河流水质的好坏, 能够说明河流生态系统的健康状况。大型底栖无脊椎动物参与河流生态系统的物质循环与能量流动, 处于河流生态系统的中间环节, 是联系能量来源和顶级猎食者的主要环节, 对维持河流生态系统的完整性起着重要的作用, 并能较好地反映过去一段时间内水体受干扰和胁迫的累积效应[3]。
拉萨河是雅鲁藏布江最大的支流, 位于雅鲁藏布江中游的左岸, 流域位于29°20′—30°15′N和90°05′—91°09′E之间, 全长551 km。河源海拔约5200 m, 汇入口海拔3580 m, 水流湍急, 水温较低[4]。根据河谷形态和地质状况, 拉萨河大致分为上、中、下游3段。上游河段从源头的麦地藏布、色荣藏布到桑曲汇入口, 长约250 km, 河床平均坡降0.37%, 中游河段从热振藏布到旁多扒曲汇入口, 长约60 km, 河床平均坡降约0.26%, 下游河段从旁多到曲水县境内雅鲁藏布江汇入口, 长约240 km, 河床平均比降约0.19%, 主要支流有麦曲、桑曲、乌鲁龙曲、扒曲、雪绒藏布、玛曲、澎波曲和堆龙曲等[5]。
拉萨河是高原河流, 其河流流域面积仅占西藏自治区总面积的2.7%, 而流域内的人口、耕地却约占全自治区的15%, 由于其特殊的地理位置以及周边的土地利用和社会经济发展状况, 注定拉萨河流域的生态状况研究有着重要的意义。简东等[6]对拉萨河中下游地区底栖动物群落特征进行过研究, 但是对整个拉萨河流域底栖动物群落结构特征以及与环境因子的关系研究未见报道, 本研究于2017年8月对拉萨河流域大型底栖动物群落结构、类群多样性进行调查与分析, 并利用多元排序统计方法来探寻大型底栖动物与环境因子的关系, 为流域的生态健康评估与生态环境管理提供科学支撑。
1 材料与方法 1.1 点位布设根据对拉萨河流域生态健康评估的研究需要, 2017年8月, 在拉萨河流域设置了23个监测点位(拉萨河干流9个, 支流14个)(见图 1), 点位覆盖拉萨河干流、重要支流及流域内重要湿地保护区。干流上游麦地卡湿地、林堤、帕热湿地、旁多电站上下游、直孔电站上下游、拉萨市、才纳9个采样点, 7条支流拉曲、雪绒藏布、玛曲、甲玛沟、彭波河、热泉及堆龙曲共设置14个采样点。

|
| 图 1 拉萨河流域采样点位图 Fig. 1 Sampling sites of river Lhasa 点位代码:拉萨河干流G1—G9;雪绒藏布Z1—Z3;拉曲Z4—Z5;玛曲Z6;甲玛沟Z7—Z8;彭波河Z9—Z10;热泉Z11—Z12;堆龙曲Z13—Z14; (“G”代表干流点位, “Z”代表支流点位) |
使用索伯网在采样断面选择3处采样点进行定量采样, 索伯网采样面积0.05 m2, 采样后将采集的样品进行分拣, 分拣出的节肢动物、软体动物用75%的酒精固定, 环节动物用5%的福尔马林固定, 固定24 h后转入70%的酒精内保存, 带回室内鉴定。大型底栖无脊椎动物在体式显微镜下观察计数, 鉴定至属级水平。
1.3 水环境理化参数测定采用WTW3210便携式pH计现场测定pH值、便携式水质监测仪(YSI 85)现场测定溶解氧DO, 根据GB3838—2002《地表水环境质量标准》, 在实验室内测定采集水样的氨氮(NH3-N)、总磷(TP)、总氮(TN)、氟化物(F-)、硫化物(S2-)、高锰酸盐指数(CODMn)、化学需氧量(CODCr)、铜(Cu)、锌(Zn)、镉(Cd)、铅(Pb)、六价铬(Cr)、汞(Hg)、砷(As), 其中重金属7项均低于检出限, 本次调查中不参与计算。样品分析方法均参照国家环境保护总局《水和废水监测分析方法》测定[7]。
1.4 数据分析采用物种数量、个体数量、优势种、生物多样性指数作为大型底栖动物特征参数, 分析流域底栖动物的群落结构特征。
1.4.1 优势度及多样性分析采用优势度[8](Y)、Margalef丰富度指数[9](R)、Shannon-Wiener多样性指数[10](H)和Pielou均匀度指数[11](J)对底栖动物群落特征进行评价:
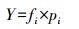
|
(1) |
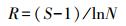
|
(2) |
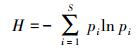
|
(3) |

|
(4) |
式中, S为总物种数;N为总个体数;pi=ni/N, pi为i种在总数量中的比例, ni为第i种个体数;fi为i种在采样点中出现频率, Y>0.02时, 定为优势种。
1.4.2 水质生物学评价利用Pielou均匀度指数J(对应水质等级划分标准见表 1)和生物指数BI进行水质生物学评价。
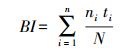
|
(5) |
| 生物指数 Biological index |
最清洁(Ⅰ) Cleanest(Ⅰ) |
清洁(Ⅱ) Clean(Ⅱ) |
轻污染(Ⅲ) Light pollution(Ⅲ) |
中污染(Ⅳ) Moderate pollution(Ⅳ) |
重污染(Ⅴ) Heavy pollution(Ⅴ) |
| Pielou均匀度指数J Evenness index J |
>0.80 | 0.61—0.80 | 0.41—0.60 | 0.21—0.40 | <0.20 |
式中, ni是第i种个体数;ti是第i个分类单元的耐污值;N是样本总个体数。
参照Karr等使用的水质生物评价指数值与分值的转换方法, 计算BI指数水质评价分级标准。具体方法为:应用所有样点的BI值, 通过频数分析, 以5%分位数对应的值作为标准, 小于该值表示水质最清洁, 大于该值至最大值分布范围四等分, 分值从小到大依次分别代表清洁、轻污染、中污染和重污染。
1.4.3 多元分析采用主成分分析(PCA)中的因子分析法, 将各采样点的环境数据进行标准化处理(其中溶解氧为逆指标, 故将其先进行倒数变换, 然后在SPSS 13.0中完成数据标准化), 为了使每个主成分的意义更加明确, 采用Varimax最大方差法对因子进行旋转, 按照特征值λ大于1的原则提取出3个主成分, 选择主成分的因子载荷量>0.7的环境因子用于底栖动物与环境因子的关系分析。
本次对拉萨河流域底栖动物与环境因子的关系分析采用多元排序方法进行。底栖动物数据与环境因子数据均进行对数转换(pH除外), 在排序前先对底栖动物相对丰度矩阵进行除趋势对应分析(detrended correspondence analysis, DCA), 若计算结果中最大的梯度长度大于3, 则选择单峰模型典范对应分析(canonical correspondence analysis, CCA)进行大型底栖动物群落结构数据与环境因子之间的关系分析, 若计算结果中最大的梯度长度小于3, 则选择线性模型RDA(redundancy analysis)进行分析。本研究的DCA结果表明, 最大的梯度长度为4.335, 因此运用CCA方法进行排序分析。采用向前引入法筛选能通过Monte Carlo置换检验的显著的环境因子变量。
底栖动物与环境因子的多元分析在Canoco for windows 4.5中完成, 成图利用Canoco Draw for windows 4.0及ArcMap 9.3完成。
2 结果与分析 2.1 大型底栖动物物种组成及分布调查期间在拉萨河流域所有干、支流23个点位共发现大型底栖动物41种, 隶属于9目30科41属(表 2)。其中水生昆虫分属6目25科35个分类单元, 占85.4%;软体动物和环节动物种类较少, 共2目4科5属, 分别为2属和3属, 各占总数的4.9%和7.3%, 铁线虫纲蠕虫1目1科1属, 占2.4%。对拉萨河流域所有点位的大型底栖动物类群相对丰度进行统计, 各样点共出现了20种优势类群, 各样点优势类群差别较大。优势种有四节蜉属、纹石蛾属、短石蛾属、水丝蚓属等。四节蜉属以及大蚊属在所有点位出现的频率高于50%, 是广泛分布物种。
| 类别 Categories |
名称 Name |
干流点位Main stream sites | 支流点位Tributary sites | ||||||||||||||||||||||||||||
| 雪绒藏布 | 拉曲 | 玛曲 | 甲玛沟 | 彭波河 | 热泉 | 堆龙曲 | |||||||||||||||||||||||||
| G1 | G2 | G3 | G4 | G5 | G6 | G7 | G8 | G9 | Z1 | Z2 | Z3 | Z4 | Z5 | Z6 | Z7 | Z8 | Z9 | Z10 | Z11 | Z12 | Z13 | Z14 | |||||||||
| 水生昆虫 Aquatic insect |
四节蜉属 Baetis spp.* |
a | a | a | a | a | a | a | a | a | a | a | a | a | a | a | |||||||||||||||
| 高地蜉属 Caucasiron sp. * |
a | a | a | a | a | a | a | a | |||||||||||||||||||||||
| 高翔蜉属 Epeorus sp. |
a | a | a | ||||||||||||||||||||||||||||
| 赞蜉属 Paegniodes sp. * |
a | a | a | ||||||||||||||||||||||||||||
| 短丝蜉属 Siphlonurus sp. |
a | a | a | ||||||||||||||||||||||||||||
| 小蜉属 Ephemerella sp. |
a | ||||||||||||||||||||||||||||||
| 晚蜉属 Teloganodes sp. |
a | ||||||||||||||||||||||||||||||
| 假二翅蜉属 Pesudocloeon sp. * |
a | a | a | a | a | a | |||||||||||||||||||||||||
| 纹石蛾属 Hydopsyche sp. * |
a | a | a | a | a | a | a | ||||||||||||||||||||||||
| 多距石蛾属 Polyplectropus sp. |
a | a | a | ||||||||||||||||||||||||||||
| 短石蛾属 Brachycentrus sp. * |
a | a | a | a | a | a | a | a | a | ||||||||||||||||||||||
| 长角石蛾属 Leptocerus sp. |
a | ||||||||||||||||||||||||||||||
| 舌石蛾属 Glossosoma sp. |
a | ||||||||||||||||||||||||||||||
| 原石蛾属 Rhyacophila sp. |
a | a | |||||||||||||||||||||||||||||
| Clostoeca sp. | a | ||||||||||||||||||||||||||||||
| 襟石蝇属 Togoperla sp. * |
a | a | a | a | a | a | |||||||||||||||||||||||||
| 狭石蝇属 Stavsolus sp. * |
a | a | a | a | a | a | |||||||||||||||||||||||||
| 叉石蝇属 Nemoura sp. * |
a | a | a | a | a | ||||||||||||||||||||||||||
| Agnetina sp. | a | ||||||||||||||||||||||||||||||
| Suwallia sp. | a | ||||||||||||||||||||||||||||||
| 大石蝇属 Pteronarcys sp. |
a | ||||||||||||||||||||||||||||||
| 冬石蝇属 Taeniopteryx sp. * |
a | a | |||||||||||||||||||||||||||||
| 狭溪泥甲属 Stenelmis sp. * |
a | a | a | a | a | ||||||||||||||||||||||||||
| 钩虾属 Gammarus sp. |
a | a | a | a | |||||||||||||||||||||||||||
| 直突摇蚊属 Orthocladius sp. * |
a | a | a | a | a | a | |||||||||||||||||||||||||
| 多足摇蚊属 Polypedilum sp. * |
a | a | a | a | a | a | a | ||||||||||||||||||||||||
| 摇蚊属 Chironomus sp. * |
a | a | |||||||||||||||||||||||||||||
| 齿斑摇蚊属 Stictochironomus sp. |
a | a | a | a | |||||||||||||||||||||||||||
| 特突摇蚊属 Thienemanninyia sp. |
a | a | a | a | |||||||||||||||||||||||||||
| 环足摇蚊属 Cricotopus sp. * |
a | a | a | a | a | a | |||||||||||||||||||||||||
| 隐摇蚊属 Cryptochironomus sp.* |
a | a | a | a | a | ||||||||||||||||||||||||||
| 锥长足虻属 Rhaphium sp. |
a | a | |||||||||||||||||||||||||||||
| 大蚊属 Tipula sp.* |
a | a | a | a | a | a | a | a | a | a | a | a | a | ||||||||||||||||||
| 蚋属 Simulium sp. * |
a | a | a | a | a | a | a | a | a | ||||||||||||||||||||||
| 拟网蚊属 Deuterophlebia sp. |
a | ||||||||||||||||||||||||||||||
| 环节动物 Annelida |
水丝蚓属 Limnodrilus sp. * |
a | a | a | a | a | a | a | a | a | |||||||||||||||||||||
| 颤蚓属 Tubifex sp. * |
a | a | a | a | a | a | a | a | |||||||||||||||||||||||
| 软体动物 Mollusca |
膀胱螺属 Physa sp. |
a | a | a | |||||||||||||||||||||||||||
| 萝卜螺属 Radix sp. |
a | a | a | ||||||||||||||||||||||||||||
| 赤豆螺属 Bithynia sp. |
a | ||||||||||||||||||||||||||||||
| 线虫动物 Nematoda |
铁线虫属 Gordius sp. |
a | |||||||||||||||||||||||||||||
| a代表在该采样点出现;标注*为优势度y>0.02的优势种;“G”代表干流点位, “Z”代表支流点位 | |||||||||||||||||||||||||||||||
拉萨河各点位物种数量和组成差别也较大, 平均物种数为8种, 最低在G8点位, 仅出现大型底栖动物1种, 而Z3采集到的大型底栖动物种类最多, 为18种。
2.2 大型底栖动物群落密度与分布从各点位的采样调查及鉴定结果看来, 点位间群落密度相差较大。最大值出现在拉萨河干流上游G2点, 该点位生态环境状况较好, 植被覆盖度较高, 河水清澈, 底质为卵石, 物种类型主要为摇蚊幼虫, 底栖动物密度为1250只/m2;最低值出现在拉萨市下游干流G8点位, 底栖动物密度为3只/m2。总体来看, 支流底栖动物密度高于干流, 支流中, 除甲玛沟和热泉(堆龙曲支流)外, 其他支流底栖动物密度均较高, 其中彭波河底栖动物密度最高, 而且上下游比较稳定。底栖动物密度空间分布情况见图 2。

|
| 图 2 拉萨河流域大型底栖动物密度空间分布特征 Fig. 2 Spatial patterns of abundance for macroinvertebrate in the Lhasa River Basin |
从拉萨河流域各点位大型底栖动物多样性的计算结果图 3来看, Margalef丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数在拉萨河干流、各支流表现出较为一致的变化趋势。拉萨河干流上游至下游, 多样性指数呈现下降趋势, 但下降幅度较小, 自直孔电站开始指数下降明显;支流中, 除雪绒藏布自上游至下游呈现明显上升趋势, 其余直流均呈现下降或者微小变化。Margalef丰富度指数与Shannon-Wiener多样性指数变化幅度较大, 而Pielou均匀度指数变化幅度相对较小;Shannon-Wiener多样性指数在拉萨河干流平均值为0.99, 最大值出现在G4旁多电站下游(1.47);支流雪绒藏布直孔电站上游Z3点Margalef指数和Shannon-Wiener指数最高, 分别为3.12和2.51。支流中甲玛沟Margalef指数和Shannon-Wiener多样性指数最低, 但Pielou均匀度指数经过比物种数的运算又弱化了这种现象。

|
| 图 3 生物多样性指数计算结果 Fig. 3 Results of biodiversity indices |
(1) 基于Pielou均匀度指数的水质评价
如图 4, 根据计算的Pielou均匀度指数按照水质等级进行划分的结果显示, 在拉萨河流域23个点位中, 多数点位处于清洁(Ⅱ)至中污染(Ⅳ)状态。清洁(Ⅱ)、轻污染(Ⅲ)和中污染(Ⅳ)点位数量分别为6个、10个、6个, 占总点位的比例分别为26.1%、43.5%、26.1%;水质评价等级为重污染(Ⅴ)的点位只有1个, 占4.3%。从拉萨河流域来看, 干流上游、下游以及彭波河水质状况较差, 水质等级为中污染(Ⅳ)和重污染(Ⅴ);干流中游以及其他支流水质状况较好, 水质等级为清洁(Ⅱ)和轻污染(Ⅲ)。

|
| 图 4 水质评价结果 Fig. 4 Water quality assessment results 水质未检出项目:Cu、Zn、Cr、Cd、Pb、Hg、As;海拔单位为m,pH无量纲,样本数单位为个,其他项目单位为mg/L |
(2) 基于底栖动物BI指数的水质评价
由前述的BI指数水质评价方法, 计算出的BI指数最小值为2.41, 最大值为6.00, 5%分位数为2.46, 根据计算结果确定的水质评价分级标准见表 3。
| 生物指数 Biological index |
最清洁(Ⅰ) Cleanest(Ⅰ) |
清洁(Ⅱ) Clean(Ⅱ) |
轻污染(Ⅲ) Light pollution(Ⅲ) |
中污染(Ⅳ) Moderate pollution(Ⅳ) |
重污染(Ⅴ) Heavy pollution(Ⅴ) |
| BI指数BI index | <2.46 | 2.46—3.35 | 3.35—4.23 | 4.23—5.12 | >5.12 |
如图 4, 在拉萨河流域23个点位中, 1个为最清洁(Ⅰ), 占总点位的比例为4.3%;4个点位为清洁(Ⅱ), 比例为17.4%;5个点位为轻污染(Ⅲ), 占21.8%;4个为中污染(Ⅳ), 占17.4%;其余9个为重污染点位, 占39.1%。从全流域来看, 基本呈现出支流以及干流的上游的水质状况优于下游的趋势。拉萨河干流从直孔电站下游起至入雅鲁藏布江前以及彭波河、甲玛沟的水质状况较差, 水质等级均是重污染(Ⅴ);干流的上游以及支流上游的水质状况较好, 水质等级为最清洁(Ⅰ)、清洁(Ⅱ)和轻污染(Ⅲ)。
2.4 拉萨河流域环境特征此次调查共计监测水质指标17项, 其中7项指标在各采样点水样中未检出, 监测结果数据特征见表 4。
| 环境因子 Environmental factors |
海拔 Altitude |
pH | 溶解氧 DO |
氨氮 NH3-N |
总氮 TN |
总磷 TP |
氟化物 F- |
高锰酸盐指数 CODMn |
硫化物 S2- |
石油类 Oil |
化学需氧量 CODCr |
样本数 nSample number |
| 平均值Mean | 4137.32 | 7.76 | 6.20 | 0.35 | 0.87 | 0.16 | 0.59 | 1.15 | 0.02 | 0.56 | 12.88 | 23 |
| 标准偏差Standard deviation | 420.09 | 0.27 | 0.56 | 0.20 | 0.48 | 0.10 | 0.37 | 0.43 | 0.01 | 0.43 | 10.19 |
流域海拔从干流最低的才纳点位3590 m—支流雪绒藏布点位5040 m, 海拔跨度较大;从水质测定结果综合来看, 拉萨河流域水体呈碱性, 监测项目均能满足《地表水环境质量标准》(GB3838—2002)Ⅲ类水体或更高水体功能标准。
2.5 大型底栖动物与环境因子的关系用主成分分析(PCA)提取特征值大于1的主成分, 共有6个, 累计贡献率达到81.9%;选取载荷超过0.7的指标作为主要影响因子, 并对相关性较强的因子组进行比较剔除, 筛选出海拔(H)、pH、总磷(TP)、总氮(TN)、硫化物(S2-)、化学需氧量(CODCr)6个主要环境因子。
大型底栖动物优势类群与主要环境因子的CCA的分析结果表 5显示, 前两个排序轴的特征值较大, 分别为0.432和0.300, 分别解释了8.7%和6.1%的底栖动物优势物种数据方差变异以及28.6%和19.8%的物种-环境关系变异。第3轴和第4轴分别解释5.3%和4.1%的底栖动物优势物种数据方差变异以及17.3%和13.6%的物种-环境关系变异;两个物种排序轴近似垂直, 相关系数为-0.02, 两个环境排序轴的相关系数为0, 说明排序轴与环境因子间线性结合的程度较好地反映了物种与环境之间的关系, 排序结果可靠[12]。从19个底栖动物优势类群和环境因子的排序结果图 5来看, 第1轴与环境因子TN、TP、S2-的相关性较高, 第2轴与CODCr的相关性较高, pH、H与两个轴的相关度相近。从各因子进入CCA分析的顺序及其解释率可以看出, H、CODCr、S2-的解释率相对较高, 对拉萨河流域大型底栖动物优势种丰度影响较显著;而pH、TP和TN的解释率相对较低, 六项环境因子之间的相关性较小。
| 项目 Item |
轴1 Axis1 |
轴2 Axis2 |
轴3 Axis3 |
轴4 Axis4 |
总惯量 Total inertia |
| 特征值Eigenvalues | 0.432 | 0.300 | 0.262 | 0.206 | 4.952 |
| 物种-环境相关系数Species-environment correlations | 0.952 | 0.911 | 0.891 | 0.819 | |
| 物种数据方差变异累计百分比/% Cumulative percentage variance of species data |
8.7 | 14.8 | 20.1 | 24.2 | |
| 物种-环境关系变异累计百分比/% Cumulative percentage variance of species-environment relation |
28.6 | 48.4 | 65.7 | 79.3 | |
| 总磷TP | -0.26 | -0.12 | 0.30 | -0.07 | |
| 总氮TN | 0.11 | 0.03 | -0.37 | 0.21 | |
| 硫化物S2- | 0.57 | -0.27 | 0.29 | 0.41 | |
| pH | -0.25 | 0.35 | -0.09 | 0.67 | |
| 化学需氧量CODCr | -0.13 | -0.62 | -0.53 | 0.17 | |
| 海拔H | -0.71 | -0.55 | 0.20 | 0.10 |

|
| 图 5 大型底栖动物群落结构与环境因子的典范对应分析排序图 Fig. 5 CCA plots of macroinvertebrate assemblages and environmental factors 图中pH为酸碱度;TP为总磷;TN为总氮;H为海拔;COD为化学需氧量;S为硫化物; 物种代码——S1:四节蜉属, Baetis spp.;S2:短石蛾属, Brachycentrus sp.;S3:纹石蛾属, Hydopsyche sp.;S4:颤蚓属, Tubifex sp.;S5:水丝蚓属, Limnodrilus sp.;S6:蚋属, Simulium sp.;S7:直突摇蚊属, Orthocladius sp.;S8:多足摇蚊属, Polypedilum sp.;S9:大蚊属, Tipula sp.;S10:摇蚊属, Chironomus sp.;S11:环足摇蚊属, Cricotopus sp.;S12:冬石蝇属, Taeniopteryx sp.;S13:隐摇蚊属, Cryptochironomus sp.;S14:狭石蝇属, Stavsolus sp.;S15:襟石蝇属, Togoperla sp.;S16:叉石蝇属, Nemoura sp.;S17:假二翅蜉属, Pesudocloeon sp.;S18:狭溪泥甲属, Stenelmis sp.;S19:高地蜉属, Caucasiron sp. |
如图 5所示, TP沿着CCA第一轴从左至右逐渐降低, 而TN和S2-沿第1轴从左往右逐渐增加。沿CCA第2轴从下向上, pH逐渐增大, CODCr逐渐减小。
排序结果表明, 不同物种对环境因子的要求不同, 因而在CCA排序图中空间位置分布不同。襟石蝇属、颤蚓属、冬石蝇属、高地蜉属以及狭溪泥甲属位于排序图的外围, 说明这几种底栖动物对特定的环境因子有响应, 如冬石蝇属分布在海拔极高的点位、颤蚓属和襟石蝇属分布在pH较高的环境中、高地蜉属分布在硫化物含量较高的环境中。而四节蜉属、纹石蛾属、大蚊属及摇蚊属等位于排序图中部, 也广泛分布于各采样点, 说明这些物种在拉萨河流域受环境影响较小, 适应性强, 为广布种。
3 讨论西藏被称为地球第三极, 平均海拔4000 m以上, 生态环境相对脆弱和敏感。拉萨河属于中纬度高海拔河流, 底栖动物的群落结构和特征既有普遍分布的特性又有区域独特的特点。
3.1 拉萨河流域大型底栖动物群落结构特征有关大型底栖动物群落的研究在低海拔地区开展较多[13-15], 高海拔地区的研究相对较少, 2010年赵伟华和刘学勤[16]以及2012年马宝珊等[17]对雅鲁藏布江底栖动物进行了初步研究, 李斌等[18]对怒江西藏段大型底栖动物群落进行了调查研究。拉萨河是雅鲁藏布江的支流, 拉萨河流域有关大型底栖动物的研究报道较少, 可参照对比的资料也较少, 徐梦珍等[19]对雅鲁藏布江流域底栖动物多样性进行研究中在拉萨河设置了3个采样点位。2013年简东等[6]对拉萨河中下游底栖动物群落结构特征进行了研究, 共设置16个采样断面, 干流和支流共采集到31种底栖动物, 本次调查为全面调查拉萨河流域的底栖动物状况, 采样点布设范围涵盖拉萨河上中下游, 共设置了26个点位, 共采集到41种底栖动物, 对拉萨河流域大型底栖动物分布特征掌握了更加丰富的信息。
纬度和海拔因子本身对河流生物不会产生直接影响, 而一般是通过温度和降水等因素间接作用于河流生态系统[20]。在本研究所设的点位中, 所有干支流上游的底栖动物种类数较下游的种类数丰富, 上游设置的采样点位受人类干扰极少, 生物多样性指数相对较高, 但远低于平原地区[21-22], Jacobsen的研究结果表明随着纬度和海拔的增加, 河流的大型底栖动物的多样性是降低的[23]。从采样过程以及调查结果来看, 拉萨河流域干支流底栖动物生物多样性较匮乏, 物种数量较少, 物种数最多的点位为支流雪绒藏布的相对上游点位Z3, 数量为18种, 物种数在十种以上的也仅有G1、G2、Z2、Z4、Z6和Z9上游点位。李宁等[24]在湟水河的研究也有同样的现象, 在47个采样点位中总分类单元数最高的为17, 且局限于源头的几个样点, 平均分类单元数为8.04;简东等对拉萨河中下游大型底栖动物采样研究中种类最丰富的布冈点位有8个物种;本研究中拉萨市下游G8点位仅发现1个物种, 各点位的平均分类单元数为7.26, 均表现出了高海拔地区河流大型底栖动物的特有的群落特征。
3.2 拉萨河流域水质评价本次研究选取了水质的一些理化指标与大型底栖动物采样调查同步进行, 以期为解释底栖动物的群落结构特征等探寻影响因素。从时间角度看, 水质理化指标的测定对象是采集到的水体样品, 具有瞬时的特点, 监测结果受外部气候环境如风力、湿度、温度等以及人为活动因子影响也较大, 受采样过程及分析过程的影响也较大。相比之下, 底栖动物的状况是环境效应累积的表现, 表征的是采样点长期的水质状况, 是长时间环境因子作用逐步形成的。拉萨河流域水质理化指标分析结果显示, 所有指标监测结果均能满足《地表水环境质量标准》(GB3838—2002)Ⅲ类水体或更高水体功能标准, 也就说明水样采集时段从理化角度看水质较好。
由于拉萨河整条河流均位于高海拔地区, 现场调查结果发现底栖动物生物多样性较低, 为了探讨应用生物指数对采样点水质进行生物学评价, 选取了能够弱化物种影响的Pielou均匀度指数以及基于物种耐污值的BI指数作为本次水质生物学评价研究的方法。
利用Pielou均匀度指数和BI生物指数进行水质生物学评价的结果显示分别有30.5%和56.5%的采样点位为中污染至重污染状态。从评价结果看来与理化指标评价的结果有出入, 究其原因是由于研究区域整体的生物多样性水平较低造成的, 也使得生物多样性指数的计算结果偏小。一般情况下, 河流上游位于林地, 并且是高海拔地区, 水温较下游偏低, 水流较快, 由于水温低, 物种会出现由于温度低、营养物质匮乏等因素而单一和稀少, 出现多样性偏低的情况, 但拉萨河流域仅一条河里呈现该现象, 为雪绒藏布支流, 比较符合内陆河流的变化趋势;而干流和其余支流均呈现上游较下游偏低或变化较小的情况, 本文认为拉萨河整条河流均位于高海拔地区, 由于下游水体水温也较低, 并且下游水流变缓, 底质不适于底栖动物生存, 故多样性指数偏低, 就会出现下游多样性指数偏低的情况。参与BI生物指数计算的物种耐污值来自国内辽河流域、安徽以及南京等地的样本, 本研究区域具有一定的地理特殊性, 物种耐污值应用的准确性及适用性有待深入的研究。通过本研究中理化手段和生物学方法对水质进行评价的过程及结果, 表明在高海拔地区使用生物学方法对水质进行评价还需进一步的修正, 才能与长久以来使用的理化手段相结合, 更好的应用于水生态系统的健康评价工作中。
3.3 优势种及其与环境因子的关系拉萨河流域干流和支流以及支流间群落组成差别较大, 不同点位的优势种不同。干流点位由于支流的汇入, 水文条件发生改变加上不同类型不同程度的人为干扰致使优势种差别较大, 最上游G1优势种为四节蜉属和短石蛾属, 到中游G3优势种为摇蚊幼虫和纹石蛾属, 进入雅鲁藏布江前的G9优势种为摇蚊幼虫。宏观看来, 随着海拔下降与人类活动干扰的增加, 干流底栖动物的优势种由较清洁的类群过渡到较耐污的类群。雪绒藏布支流从上游Z1到下游Z3耐污种类逐渐增多, 不过每个点位都有襀科清洁种存在, 表明河流水质还是较好的;拉曲支流上游Z4点位优势种由指示较清洁的四节蜉属、短石蛾属、大蚊属以及中等耐污的摇蚊幼虫等组成, 下游Z5点位优势种清洁种减少, 较耐污的纹石蛾属成为优势种;甲玛沟、彭波河、热泉以及堆龙曲几条支流的优势种也是相似的状况。从不同支流优势种看来, 不同支流的优势种不同, 雪绒藏布有襀科物种、热泉有赞蜉、拉曲有毛翅目物种、彭波河有假二翅蜉, 根据李宁等[24]的研究成果, 青藏高原高山大川, 地理隔绝明显, 不同河段海拔高度不同, 底栖动物单独进化, 使得不同溪流底栖动物的优势种及群落结构差别较大。
大型底栖动物不仅受到底质影响, 也受到底层水环境的影响[25]。CCA结果表明拉萨河流域大型底栖动物群落结构受海拔、总氮、pH值、化学需氧量、硫化物、总磷影响较多, 其中海拔是影响底栖动物优势种分布的关键因子, 其次为水体化学物质含量以及总磷总氮营养盐。海拔决定了一个地区的温度和光照等环境因素的变化, 属于宏观尺度的环境因子, 因此从根本上决定了河流底栖动物的群落结构组成[21]。CODCr和硫化物作为判别底栖动物优势种的指标说明了不同点位污染来源和程度的不同。水体中总磷、总氮等营养盐在一定程度上也影响着拉萨河流域大型底栖动物优势种的空间分布格局, 在对太子河流域、西苕溪[26]以及鱼山渔场[27]等的研究中, 发现总氮对底栖动物的影响较为严重, 张又等[28]在巢湖流域的研究中也有相似的结果, 营养盐的含量与水体中其他化学物质也是有关联的, 根据deJonge等[29]的研究结果, 在营养盐浓度较高的水体中, 致使底层水环境溶氧较少并增加底质中和水体中硫化物的含量, 限制了敏感种的分布。从以上结果与讨论得出, 拉萨河属高原河流, 流域底栖动物优势种的组成分布与自然因素海拔密切相关, 同时污染及营养物质输入等人为干扰因素也是关注的重点。
鉴于拉萨河流域特殊的地理区位及本次研究的结果, 今后应加强区域大型底栖动物与更多环境因子的调查、监测及研究, 完善水环境评价体系, 通过找出影响底栖动物分布的因素, 再反过来从因素的角度出发探讨改善流域生态环境, 为相关部门提供决策参考, 为拉萨河水生态健康的评估及保护提供指导, 用以推动区域自然、经济和社会的和谐发展。
4 结论(1) 拉萨河流域调查共发现大型底栖动物41个分类单元, 各点位间底栖动物物种数量和密度差别较大, 物种数量变化范围为1—18种, 物种密度变化范围为3—1250只/ m2。
(2) Margalef丰富度指数R、Shannon-Wiener多样性指数H和Pielou均匀度指数J在拉萨河干流、各支流表现出的趋势较为一致。利用Pielou均匀度指数法和底栖动物BI指数法进行了水质评价, 两种评价结果显示分别有30.3%和56.5%的采样点水质为中污染至重污染状态。
(3) 拉萨河流域大型底栖动物空间分布主要受到海拔、硫化物、化学需氧量、总磷、总氮和pH等6个因子影响。
| [1] |
Karr J R. Assessment of biotic integrity using fish communities. Fisheries, 1981, 6(6): 21-27. DOI:10.1577/1548-8446(1981)006<0021:AOBIUF>2.0.CO;2 |
| [2] |
Bonada N, Prat N, Resh V H, Statzner B. Developments in aquatic insect biomonitoring:a comparative analysis of recent approaches. Annual Review of Entomology, 2006, 51: 495-523. DOI:10.1146/annurev.ento.51.110104.151124 |
| [3] |
李镇, 张岩, 袁建平, 卢路. 大型底栖无脊椎动物在河流健康评价中的发展趋势. 南水北调与水利科技, 2011, 9(4): 96-101. |
| [4] |
中国科学院青藏高原综合科学考察队. 西藏河流与湖泊. 北京:科学出版社, 1984, 108-142. |
| [5] |
布多, 许祖银, 吴坚扎西, 李明礼, 旦增, 德吉. 拉萨河流域选矿厂分布及其对环境的影响. 西藏大学学报:自然科学版, 2009, 24(2): 33-38. |
| [6] |
简东, 黄道明, 常秀岭, 张庆, 谢山, 陈峰, 陈胜. 拉萨河中下游底栖动物群落结构特征分析. 水生态学杂志, 2015, 36(1): 40-46. |
| [7] |
国家环境保护总局. 水和废水监测分析方法 (第四版). 北京: 中国环境科学出版社, 2002.
|
| [8] |
徐兆礼. 东海亚强真哲水蚤种群生态特征. 生态学报, 2006, 26(4): 1151-1158. DOI:10.3321/j.issn:1000-0933.2006.04.024 |
| [9] |
Margalef R. Diversidad de especies en las comunidades naturales. Publicacions del Institute de Biologia Aplicada, 1951, 9(5): 5-27. |
| [10] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1963.
|
| [11] |
Pielou E C. Ecological Diversity. New York: John Wiley, 1975.
|
| [12] |
Ter Braak C J F. Canonical correspondence analysis:a new eigenvector technique for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167-1179. |
| [13] |
殷旭旺, 韩洁, 王博涵, 金文, 杨璐, 陈海, 吴丹. 太子河流域底栖动物群落结构及其与环境因子的关系. 水产学杂志, 2017, 30(3): 40-44. DOI:10.3969/j.issn.1005-3832.2017.03.008 |
| [14] |
段学花, 王兆印, 程东升. 典型河床底质组成中底栖动物群落及多样性. 生态学报, 2007, 27(4): 1664-1672. DOI:10.3321/j.issn:1000-0933.2007.04.047 |
| [15] |
杨莲芳, 李佑文, 戚道光, 孙长海, 田立新. 九华河水生昆虫群落结构和水质生物评价. 生态学报, 1992, 12(1): 8-15. DOI:10.3321/j.issn:1000-0933.1992.01.010 |
| [16] |
赵伟华, 刘学勤. 西藏雅鲁藏布江雄村河段及其支流底栖动物初步研究. 长江流域资源与环境, 2010, 19(3): 281-286. |
| [17] |
马宝珊, 谢从新, 杨学峰, 霍斌. 雅鲁藏布江谢通门江段着生生物和底栖动物资源初步研究. 长江流域资源与环境, 2012, 21(8): 942-950. |
| [18] |
李斌, 徐丹丹, 刘绍平, 王志坚. 怒江西藏段大型底栖动物群落结构及多样性研究. 淡水渔业, 2015, 45(2): 43-48. DOI:10.3969/j.issn.1000-6907.2015.02.008 |
| [19] |
徐梦珍, 王兆印, 潘保柱, 巩同梁, 刘乐. 雅鲁藏布江流域底栖动物多样性及生态评价. 生态学报, 2012, 32(8): 2351-2360. |
| [20] |
Dudgeon D. Responses of benthic macroinvertebrate communities to altitude and geology in tributaries of the Sepik River (Papua New Guinea):the influence of taxonomic resolution on the detection of environmental gradients. Freshwater Biology, 2012, 57(9): 1794-1812. DOI:10.1111/fwb.2012.57.issue-9 |
| [21] |
渠晓东, 曹明, 邵美玲, 黎道丰, 蔡庆华. 雅砻江(锦屏段)及其主要支流的大型底栖动物. 应用生态学报, 2007, 18(1): 158-162. DOI:10.3321/j.issn:1001-9332.2007.01.027 |
| [22] |
曹艳霞, 蔡德所, 张杰, 赵湘桂, 王备新. 漓江水系大型无脊椎底栖动物多样性现状调查. 广西师范大学学报:自然科学版, 2009, 27(2): 118-123. |
| [23] |
Jacobsen D, Schultz R, Encalada A. Structure and diversity of stream invertebrate assemblages:the influence of temperature with altitude and latitude. Freshwater Biology, 1997, 38(2): 247-261. DOI:10.1046/j.1365-2427.1997.00210.x |
| [24] |
李宁, 陈阿兰, 杨春江, 马琪, 马国良, 孙瑜旸, 白露超. 湟水河上游大型底栖动物多样性及水体理化因子调查分析. 中国农学通报, 2016, 32(23): 43-48. DOI:10.11924/j.issn.1000-6850.casb16050046 |
| [25] |
张莹, 吕振波, 徐宗法, 刘义豪, 靳洋. 山东半岛南部海湾底栖动物群落生态特征及其与水环境的关系. 生态学报, 2011, 31(15): 4455-4467. |
| [26] |
吴东浩, 于海燕, 吴海燕, 周斌, 王备新. 基于大型底栖无脊椎动物确定河流营养盐浓度阈值——以西苕溪上游流域为例. 应用生态学报, 2010, 21(2): 483-488. |
| [27] |
谢旭, 俞存根, 郑基, 夏陆军, 毕耜瑶, 张平, 邓小艳, 苗露. 鱼山渔场披山海域虾类群落结构及其与环境因子的关系. 大连海洋大学学报, 2017, 32(6): 713-723. |
| [28] |
张又, 程龙, 尹洪斌, 高俊峰, 张志明, 蔡永久. 巢湖流域不同水系大型底栖动物群落结构及影响因素. 湖泊科学, 2017, 29(1): 200-215. |
| [29] |
de Jonge M, Dreesen F, de Paepe J, Blust R, Bervoets L. Do acid volatile sulfides (AVS) influence the accumulation of sediment-bound metals to benthic invertebrates under natural field conditions?. Environmental Science & Technology, 2009, 43(12): 4510-4516. |