 2019, Vol. 39
2019, Vol. 39文章信息
- 周世强, 张晋东, Vanessa HULL, 黄金燕, 刘巅, 周季秋, 孙萌萌, 张和民
- ZHOU Shiqiang, ZHANG Jindong, Vanessa HULL, HUANG Jinyan, LIU Dian, ZHOU Jiqiu, SUN Mengmeng, ZHANG Hemin
- 野生大熊猫与放牧家畜的活动格局比较
- Comparative activity patterns of wild giant pandas and livestock
- 生态学报. 2019, 39(3): 1071-1081
- Acta Ecologica Sinica. 2019, 39(3): 1071-1081
- http://dx.doi.org/10.5846/stxb201804170879
-
文章历史
- 收稿日期: 2018-04-17
- 网络出版日期: 2018-11-15

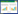




2. 西华师范大学西南野生动植物资源保护教育部重点实验室, 南充 637002;
3. Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, FL, USA
2. Key Laboratory of Southwest China Wildlife Resources Conservation, China West Normal University, Ministry of Education, Nanchong 637002, China;
3. Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, FL, USA
野生大熊猫(Ailuropoda melanoleuca)是世界上生物多样性保护的旗舰物种, 目前间断分布在四川、甘肃和陕西三省的凉山、小相岭、大相岭、邛崃山、岷山和秦岭山系, 据第四次全国大熊猫调查(2011—2014年)统计, 其种群数量约1864只;与第三次调查相比(1999—2003年), 无论是种群数量, 还是栖息地面积均有所增长(种群数量增长了16.8%, 栖息地面积增长了11.8%)[1-3]。然而, 野生大熊猫栖息地的破碎化并未减缓, 局域种群之间的基因交流仍旧隔离严重, 这主要受公路、村庄、水电设施等人为干扰所致[2-5]。在对人为因素(一般干扰类型)的排序中, 以全国第四次大熊猫调查的数据为例, 放牧占据着首要位置, 其海拔跨度大(1500—3600 m)、分布范围广(四川、陕西和甘肃的大熊猫分布区都有出现, 干扰点约3311个)、野外遇见率高(高达33.90%), 是造成大熊猫栖息地质量和数量下降的诱因之一[2-3, 6-12]。放牧不仅导致野生动物生境退化与丧失、疫病扩散、环境污染和气候变化等[13], 而且影响放牧区域森林生态系统的碳储量、栖息地的物种多样性、野生动物的活动格局和草食动物的觅食策略[14-21]。具体到野生大熊猫而言, 放牧活动既破坏栖息地的结构与功能(如草本植物的组成成分变化、藤本植物的人为伐除、灌木植物的践踏死亡、森林的水土保持能力减弱、枯落物的腐烂分解速度趋缓等)、降低大熊猫的生境利用率和加大食物资源(可食竹)的耗损量[11-12, 22], 而且影响大熊猫的空间利用格局和觅食行为策略, 以及伴生动物的共存空间变化[11, 23-25]。
活动格局主要是指动物的昼夜活动节律及其活动强度, 为动物最为重要的行为生态特征, 不仅受制于动物的遗传机制[26-28], 而且反应出动物对环境的适应规律, 比如动物的能量代谢能力、觅食行为策略、进化适应及生理响应[29-31], 动物所处的生理阶段(繁殖期、非繁殖期等)和时空位置[32-35], 以及环境因子(地形地势、气候条件、光照强度、生境结构、食物资源、人为干扰等)对动物活动格局的影响[32-36]。对于野生大熊猫活动节律的研究起始于20世纪80年代, 先后在四川卧龙(1980—1983年)、陕西长青(1989—1997年)和佛坪(1991—1995年)通过无线电项圈遥测技术进行跟踪, 初步破解了竹林隐士的生活奥秘, 为大熊猫的保护管理奠定了基础[37-41];进入本世纪(陕西佛坪2008—2009年、四川卧龙2010—2012年), 人们采用先进的全球定位系统跟踪设备(GPS颈圈)获取了全天候、各时段和长时间的大量定位和活动数据, 进一步洞悉了大熊猫更为系统的活动规律, 深度揭示了大熊猫的生理、生态学特征[25, 42-44]。对于大熊猫分布区域内放牧家畜对大熊猫及其栖息地和主食竹的影响研究较多, 而很少对其活动格局以及与大熊猫相比较的分析[6-11, 19]。由于不同的时间利用规律和不同的空间利用模式形成动物之间不同的生境选择特性、食物利用策略, 进而对其生活的环境造成不同的影响强度和生态后果, 尤其是人为引入的外来物种(如放养家畜)。如果人们在未深入了解引入物种及其与目标物种(如野生大熊猫)之间在空间利用、觅食策略和活动格局等方面的差异性, 就盲目将其释放于目标物种的生活区域, 不仅会导致资源竞争(如栖息地、食物等)、空间萎缩或生境破碎化, 而且影响目标动物的种群发展和基因交流, 甚至具有局部区域的物种灭绝风险[13-25]。因而, 加强引入物种(放牧家畜)与野生大熊猫的生态学比较研究势在必行, 以便为大熊猫的保护管理提供依据。
为此, 本文在Hull等[11, 45]、Zhang等[25, 43]和周世强等[23]的研究基础上, 以卧龙自然保护区“核桃坪”区域放牧的马群(Equus sp.)为例, 通过分析野生大熊猫和放牧家畜的活动强度、活动时间百分率、活动时间片段率和活动片段时长等指标的差异性, 来探讨两者在时间利用方面的内在特征。
1 研究方法 1.1 研究区域自然地理特征研究地点位于四川卧龙国家级自然保护区(102°51′—103°24′E、30 °45′—31°25′N)的“核桃坪”及其毗邻区域(简称“核桃坪区域”), 由“牛头山”和“七层楼沟”之间的山脊、森林、灌丛、草地、沟壑和河流(皮条河)组成, 覆盖面积大约40 km2, 海拔范围1510—3200 m(图 1)。该区域山高谷深、沟壑纵横、坡度比降大(坡度变化从20°到50°以上), 是典型的高山峡谷地貌。从低海拔到高海拔, 由于山地气候效应的作用, 气温垂直变化明显, 从而形成了不同的森林植被类型, 递次分布有常绿落叶阔叶林(1510—1800 m)、落叶阔叶林(1800—2300 m)、针阔混交林(2300—2700 m)、亚高山暗针叶林(2700—3200 m)和高山灌丛(仅零星分布)等, 与卧龙整个自然保护区相同地带的植被类型相似。构成各植被结构中灌木层的优势成分除杜鹃(Rhododendron spp.)、悬钩子(Rubus spp.)、栒子(Cotoneaster spp.)、泡花树(Meliosma spp.)、茶藨子(Ribes spp.)、忍冬(Lonicera spp.)、菝葜(Smilax spp.)等灌木植物外, 盖度达85%以上的组成因子是野生大熊猫喜食的亚高山中小径竹。拐棍竹(Fargesia robusta)生长于海拔2600 m以下的森林植被中, 2600—3200 m主要以冷箭竹(Bashania faberi)为主, 短锥玉山竹(Yushania brevipaniculata)在海拔2300—2700 m与拐棍竹或冷箭竹呈镶嵌分布[11, 23, 42, 46-47]。

|
| 图 1 研究区域位置及大熊猫和放牧家畜的GPS颈圈跟踪 Fig. 1 Map of the study area in Wolong Nature Reserve and GPS collar tracking of wild giant pandas and livestock (Horses) |
“核桃坪区域”是卧龙自然保护区野生大熊猫数量分布较为集中的地区, 根据红外数码相机和粪便DNA测定, 大约有22只大熊猫生活其中[43, 48]。其东北面与四川汶川卧龙特别行政区耿达镇的老鸦山牧场相邻, 西南面与卧龙镇三村的鱼丝洞牧场接壤, 两牧场是当地村民的天然放牧地, 主要以马、牦牛(Bos grunniens)、绵羊(Ovis aries)、山羊(Capra hircus)和黄牛(B. tourus)等家畜为主[11, 23]。由于两牧场面积小、载畜量低, 又紧邻村庄和农耕地, 因而放牧家畜数量受限, 2004、2007和2011年两乡镇部分村民擅自将牲畜驱赶至拐棍竹林或冷箭竹林中放养, 导致竹子资源和栖息地受到严重影响。有鉴于此, 2013年当地的保护管理机构和政府部门责令村民将马匹收回并出售[11, 23, 25]。
1.2 研究方法 1.2.1 研究对象概述野生大熊猫的研究对象主要从2010—2012年在“核桃坪区域”佩戴GPS颈圈跟踪监测的5只个体中选择3只作为与放牧家畜进行比较的样本, 选择标准基于监测时间至少在一年以上的成体雌性和雄性大熊猫, 利于分析不同季节、不同月份的活动情况[23, 49]。放牧家畜主要在大熊猫栖息地内活动的马群中选择代表性个体, 以成年健壮马为对象, 雌雄个体兼有, 并佩戴GPS颈圈和命名, 以便野外监测、数据采集和日常饲喂, 选择地点分别位于“趴趴沟”、“鱼丝洞”和“七层楼沟”(表 1)[23]。
| 动物名称Animal Name | 川川 | 美美 | 中中 | 东东 | 娜娜 | 牛牛 |
| 颈圈编号Collar ID | 30856 | 1188 | 30857 | 259 | 1094 | 30859 |
| 年龄Age | 成体 | 成体 | 成体 | 成体 | 成体 | 成体 |
| 性别Gender | 雌性 | 雌性 | 雄性 | 雄性 | 雌性 | 雌性 |
| 开始时间Start Time | 2011-03-31 | 2010-03-29 | 2011-03-16 | 2011-06-02 | 2010-07-26 | 2011-05-07 |
| 结束时间Stop Time | 2012-08-11 | 2012-04-05 | 2012-08-07 | 2012-07-23 | 2012-07-23 | 2012-07-20 |
| 分析时段Duration of Study | 2011-04-01—2012-03-27 | 2011-04-01—2012-03-31 | 2011-04-01—2012-03-31 | 2011-07-01—2012-06-30 | 2011-07-01—2012-06-30 | 2011-07-01—2012-06-30 |
| 数据量Number of Data(n) | 83796 | 100110 | 76469 | 100471 | 100393 | 104019 |
| 传感效率Sensor frequency/% | 79.50 | 94.97 | 97.08 | 95.31 | 95.28 | 98.68 |
| 动物类型Animal Type | 大熊猫 | 大熊猫 | 大熊猫 | 放牧家畜 | 放牧家畜 | 放牧家畜 |
| 监测地点Monitoring Site | 核桃坪 | 核桃坪 | 核桃坪 | 趴趴沟 | 鱼丝洞 | 七层楼 |
野生大熊猫和放牧家畜(马)均采用内置全球定位系统(GPS)、具有无线电遥测和远程数据下载功能的定位颈圈, 型号Lotek_4400 M、大小120 mm×86 mm×126 mm、总重1200 g[23, 42-43]。该型号颈圈不仅有12个GPS定位频道、还安装了Mini自动脱落装置(启动后2年左右自动从监测对象身体上脱落, 保障了动物的自然生长发育), 而且内设有记录水平和垂直方向活动强度的传感器。传感器每5 min记录1个数据, 包括日期、时间、温度、水平(X)和垂直(Y)方向指标值(0:禁止不动;255:活动最强烈)。此外, 颈圈还按照工作人员设置的定位日程, 每2 h记录1次GPS定位信息, 主要含有日期、时间、东经(°)、北纬(°)、海拔高度(m)、定位质量(PDOP)和导航(NAV)等数据[23, 42-43]。为了保障数据收集的有效性, 了解研究对象的活动区域和生存状况, 每月进行1—2次的无线电监听, 每2个月使用手持终端机(HCU)远程下载1次定位和活动数据。
大熊猫和马的GPS颈圈佩戴过程, 包括人工捕捉、麻醉和安装, 都有中国大熊猫保护研究中心兽医参与, 尤其是大熊猫的麻醉工作主要由专职兽医负责, 而且该项目得到原国家林业局和卧龙自然保护区管理局的审批和准许。大熊猫GPS颈圈的安装于2010年和2011年的3月下旬至4月上旬进行, 放牧家畜则在2010年和2011年5—7月之间佩戴(事先征得牲畜主人的许可, 并一同前往)。具体工作流程参见下图(图 2)及文献所述[23, 42-43]。

|
| 图 2 野生大熊猫GPS颈圈佩戴的工作流程图 Fig. 2 Working flow-chart of wild giant pandas fitted with GPS collars |
分析有关大熊猫活动格局研究的文献, 人们主要从活动强度、活动时间百分率两个方面进行活动节律探讨, 以及外部环境(光照强度、气温等)对野生大熊猫活动节律的影响[25, 37-39, 42-43]。由于动植物的生物钟(活动节律)受其周期基因(Period Gene)的控制[26-28], 同时与其体型、觅食策略和所处环境密切相关[25, 29, 43], 基于GPS颈圈活动记录的数据海量(表 1)和时间较长的特点, 在此引入活动行为连续性(Continuity)与持续性(Durative)概念(亦即活动时间片段率与活动片段时长), 在分析大熊猫与放牧家畜除在能耗性(Energy consumption)和周期性(Periodicity)(即活动强度和活动时间百分率)的基础上, 进一步探讨两者活动格局的差异。
(1) 活动强度 活动强度是指GPS颈圈内活动传感器记录的圆球在水平和垂直方向撞击圆筒的次数(记录值为0—255), 并以此确定动物在某一时刻的活动强度[23, 42-43]。根据研究, 颈圈中传感器的灵敏性在垂直方向的准确度高于水平方向[43, 50], 为此, 本文采用大熊猫和放牧家畜GPS颈圈中的垂直记录值作为活动强度的指标。
(2) 活动时间百分率 经过对圈养大熊猫不同行为类型与GPS颈圈中活动值的对应试验, 显示0值为不活动行为(如休息或禁止不动), 其他值(大于0至255)为活动行为(如采食、饮水、走动等)[43]。因而, 在此的活动时间百分率是指单位时间内(1小时、1天、1月或1年)除去0值的活动时间与总时间的百分比[37, 39, 40, 42-43]。
(3) 活动时间片段率 无论是圈养动物还是野生动物都不可能只有一个行为, 其某一个行为也不可能无限持续, 而呈现出不同行为类型之间的交替进行, 因此在GPS颈圈中显示为0值和其他值的镶嵌分布, 具有活动行为或不活动行为的片段化特征。活动片段率是指单位时间内活动行为的片段数(NFAT)与活动时间(TA)的比率(式1)。活动片段的计数方法是根据活动值的时间连续性进行标记, 例如8:00时活动值大于0, 记为1个片段(编号), 如果8:05仍然大于0, 就记录为相同的片段编号;如果为0值, 则中断此片段编号, 下一时段重新编号, 以此类推。
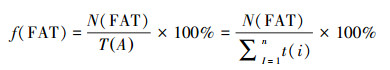
|
(1) |
式中, f(FAT)为活动时间片段率, N(FAT)为活动时间片段数, T(A)为总的活动时间(min), t(i)为每个片段的时长(min)。以24 h为单位时间, GPS颈圈每5 min记录一个活动数据, 那么最多的活动片段数达288个、最少的仅1个, 于此最大的片段率为20.00%, 最小的是0.07%。
(4) 活动片段时长 根据上述的片段计数法, 每个独立片段的时间即为一个活动片段时长(min), 理论上最短的片段时长为5 min, 最长的为1440 min(以24小时为单位时间)。为了比较野生大熊猫与放牧家畜的活动片段时长的差异, 我们对片段时长进行分级, 并统计各级的片段时长的时间密度(%), 其计算公式如下(公式2)。活动片段时长共分成9级, 各级的时间尺度为1:5 min、2:10—30 min、3:35—60 min、4:65—120 min、5:125—180 min、6:185—240 min、7:245—300 min、8:305—360 min和9:≥365 min, 并计算各级的时间密度(以1天为计算单元)。
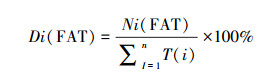
|
(2) |
式中, Di(FAT)为第i级片段时长的时间密度(%), Ni(FAT)为第i级片段时长的活动时间片段数, T(i)为第i级片段时长的活动时间(min), i=1—9。
1.2.4 数据处理和统计分析对从野外通过HCU远程下载的试验动物所佩戴的GPS颈圈中储存的活动数据, 利用GPS_plus 3.9软件转存电脑, 同时转化为数据库文件, 并在Microsoft Excel 2013中进行时间矫正(因GPS颈圈存储数据的时间格式有格林威治时间(GMT)和当地时间(LMT), 有时在GPS设置时可能存在时间未加上北京时区值)、数据校核(可能由于环境条件的影响, 传感器记录的日期或时间发生错误, 比如小于或大于佩戴颈圈的日期、两相邻时间重叠等), 以及活动时间片段的编码等。由于野生大熊猫和放牧家畜佩戴GPS颈圈的年度和日期有差异, 加之野外环境的影响(尤其是大熊猫颈部的特殊性, 常导致大熊猫的颈圈跟踪时间长短不齐, 最短的仅43 d, 最长的达731 d), 所以为了有效分析放牧家畜和野生大熊猫活动格局的差异, 我们选择2011年和2012年之间的监测数据作为分析样本, 且保证监测时间都在一年以上, 并尽量避免佩戴颈圈对动物行为的影响[23](表 1)。每个试验动物的活动数据汇总后, 采用SPSS 20.0和Origin 2016进行统计分析与绘图。因采用One-Sample Kolmogorov-Smirnov Test方法检验每个动物的活动数据, 表明所有数据都遵循正态分布和方差齐性, 故使用方差分析(ANOVA)和多重比较(Fisher LSD法)进行个体或物种间(大熊猫与马)的显著性检验[51-52]。平均值以平均数±标准差(Mean±SD)表示, 显著性水平设置为0.05。
2 结果与分析 2.1 野生大熊猫与放牧家畜的活动强度通过对3只野生大熊猫和3只放牧家畜1年内垂直方向活动数据的整合统计, 结果表明放牧家畜的活动强度极显著地大于野生大熊猫(df1=1, df2=2133, F=522.077, P < 0.0001), 前者平均是28.93±20.13、后者为14.99±17.86(含活动水平0值)或34.28±8.73和25.18±9.67(不含活动水平0值)。
从日活动强度的时间分布来看(图 3), 野生大熊猫和放牧家畜的活动水平都在10.00以上, 白天(6:00—18:00)高于夜间(0:00—6:00和18:00—24:00), 具有明显的活动高峰期, 且不同时间之间表现出明显的统计学意义(大熊猫:df1=23, df2=273127, F=527.725, P < 0.0001;放牧家畜:df1=23, df2=304859, F=1258.127, P < 0.0001)。从不同月份和季度活动水平的动态分析(图 3), 大熊猫不同月份之间变化明显, 呈现出季节性波动(df1=11, df2=273139, F=778.772, P < 0.0001), 春季(4—6月)和冬季(11月至次年3月)活动强度大于夏秋季;而放牧家畜从4月份逐渐增强, 6月份达到峰值后, 一直持续下降, 不同月份、不同季节之间均具有极显著性差异(P < 0.0001)。

放牧家畜的活动时间百分率极大地高于野生大熊猫, 是后者的1.51倍, 据统计野生大熊猫的活动时间百分率为(53.91±12.64)%、放牧家畜是(81.34±5.93)%, 且两者之间具有极显著性差异(df1=1, df2=2133, F=4195.413, P < 0.0001)。从活动节律分析, 野生大熊猫和放牧家畜都表现出3个明显的活动高峰和2个活动低谷, 活动高峰分别是黎明(6:00—9:00)、黄昏(16:00—19:00)和子夜(23:00—1:00), 活动低谷都在5:00和22:00两个时间段(图 4)。从不同月份的活动时间百分率变化规律来看, 野生大熊猫的曲线起伏较大具有明显的季节性(P < 0.0001), 放牧家畜则较为平缓, 其春季和夏秋季与冬季之间差异显著(P < 0.0001)、春季与夏秋季之间无明显的统计学意义(P=0.3900)(图 4)。

|
| 图 4 不同时间和月份野生大熊猫与放牧家畜的活动时间百分率 Fig. 4 Mean hourly and monthly percentage of time active for wild pandas and livestock |
统计野生大熊猫与放牧家畜整年内日活动时间的片段率平均为野生大熊猫(1.62±0.86)%、放牧家畜(1.40±0.45)%, 野生大熊猫明显大于放牧家畜的活动时间片段率(F=73.956, P < 0.0001)(图 5)。不同月份两者之间野生大熊猫的5—11月都大于放牧家畜, 仅1—4月和12月略低(图 5)。不同季节两者之间均具有极显著性差异(P < 0.0001), 春季和夏秋季野生大熊猫高于放牧家畜, 冬季则低于放牧家畜(图 5)。

|
| 图 5 不同月份野生大熊猫与放牧家畜的活动片段率 Fig. 5 Mean monthly ratio of fragments of time active for wild pandas and livestock |
从图 6中的曲线显示, 活动时间片段率(图 5)与活动片段时长呈相反的规律。比较整年日平均活动时长, 野生大熊猫与放牧家畜之间无显著性差异(F=0.667, P=0.4107), 分别为野生大熊猫平均(77.63±37.67)min, 最大均值223 min, 极值440 min;放牧家畜平均(79.59±28.49)min, 最大均值271 min, 极值425 min。从不同月份活动时长的统计结果来看, 野生大熊猫5—9月略低于放牧家畜, 1—4月和10—12月较后者为高(图 6)。将两者分不同季节进行比较, 野生大熊猫与放牧家畜之间有明显的差异(F=7.069, P=0.0079), 野生大熊猫各季节的活动时长分别是春季(70.23±31.97)min、夏秋季(66.30±32.97)min、冬季(93.32±40.08)min, 放牧家畜依次是春季(82.81±29.98)min、夏秋季(84.87±29.23)min、冬季(73.39±25.73)min(图 6)。

|
| 图 6 不同月份野生大熊猫与放牧家畜的活动片段时长及各级活动时间的密度 Fig. 6 Mean monthly duration of fragmentation active and mean fragmentation time density of each duration scales of action time segment for wild pandas and livestock 片段时长分级标准(min)::1—9分别代表:5, 10—30, 35—60, 65—120, 125—180, 185—240, 245—300, 305—360, ≥365 min |
为了更加清晰地了解野生大熊猫与放牧家畜活动片段时长的差异性, 将活动片段时长进行分级, 并计算各级的时间密度(%)。经统计分析, 两者均呈现为正态分布图形, 其片段时长都集中于4—6级, 野生大熊猫达到66.76%、放牧家畜约61.99%(图 6)。从两者不同时间尺度级条形图的分布格局来看, 野生大熊猫的1—6级(除3级外)均略高于放牧家畜, 相差范围在0.44%—2.43%之间;而7—9级则为前者低于后者, 差值达到-1.51%—-2.94%(图 6)。统计检验表明, 两者总体之间的差异无统计学意义(F=0.7274, P=0.3938), 而相同时间尺度级的野生大熊猫与放牧家畜除1、2、3、6之间没有明显的差异外(P=0.2954—0.6330), 其余都差异极显著(P < 0.01)。
3 讨论 3.1 野生大熊猫与放牧家畜活动的能耗性与周期性统计卧龙自然保护区“核桃坪区域”内野生大熊猫个体和自由散放家畜的活动强度, 结果表明野生大熊猫的活动水平明显低于放牧家畜(图 3), 是能量低耗性的物种, 这与圈养和野生大熊猫能量研究的结论一致[31]。分析大熊猫与放牧家畜能耗性差异的内在机制, 这不仅与动物的质量(体重)、体型大小、活动规律和觅食策略有关, 而且与其生存环境和遗传基因相联[26-29, 31-36]。虽然成年大熊猫的体重平均在85—110 kg, 体长为80—90 cm, 体高约60 cm, 并非属于小体型动物[37-38](本次估测数据, 2010—2012年), 但其行为独特, 独居和隐蔽生活, 一般情况下步行缓慢、移动距离短(日移动距离仅200 m左右), 采食时通常以坐立方式进行[23, 37-38, 43, 53];生境结构简单, 主要生存于落叶阔叶林、针阔混交林和亚高山暗针叶林, 乔灌草的物种多样性较低[46-47, 54];地形地势复杂, 处在四川盆地向青藏高原过渡地带, 山高谷深、沟壑纵横、坡陡径险[46];食性特异, 通过几百万年的进化适应, 大熊猫特化成以亚高山中小径竹为食物来源, 并且资源相对储量较大、易于获取[1, 12, 55];以及具有独一无二的基因使其能依赖于低营养的竹子而长期生存[31]。相对于大熊猫而言, 放牧家畜(马)不仅个体高大(体重约400—800 kg、体长150—200 cm、体高100—150 cm;本次估测数据, 2010—2012年)、营群体生活、善于奔跑、人为干预较多, 颈部较长、且站立采食;生境变化不定, 通常往来于森林植被和草地灌丛之间;食物种类较多, 除利用森林内竹子资源外、林中的灌草植物和牧场草本亦被采食[44](野外观察, 2010—2012年)。
与活动强度相似, 野生大熊猫的活动时间较放牧家畜为少, 其休息时间显著长于马群(图 4), 这也是大熊猫节省能量的策略之一[37-38, 43]。从活动节律来看, 野生大熊猫与放牧家畜差异不十分明显, 都具有3个活动高峰和两个活动低谷, 仅在时间节点上略有不同(图 4), 形成两者活动节律的影响因素与活动强度的机制相似。
不同月份或季节的活动变化规律是与动物所居环境的气候变化、动物的生理阶段和食物类型的时间更替密切相关, 因而形成了野生大熊猫和放牧家畜的季节活动格局(图 4、图 5)。春季(4—6月)气温逐渐回暖、大熊猫处于发情交配期以及主食竹笋(主要是拐棍竹)的萌发生长, 致使野生大熊猫的活动范围扩大、活动水平增高(寻找配偶、撵食竹笋等), 为成功交配、妊娠育幼等积累能量;夏秋时节(7—10月), 一方面因较低海拔的气温偏高, 多数熊猫生活在高海拔地区, 活动范围相对缩小, 二是受孕母兽在树洞或岩穴产仔育幼, 活动强度降低, 三是营养丰富的嫩枝嫩叶量大质优, 不用远程觅食;而寒冷的冬季(11月至次年3月), 大熊猫不仅需要从高海拔生境迁移至较低的区域、长距离跋涉至不冻溪沟或河流饮水、厚厚积雪艰难爬行, 而且冬季的食物数量和质量相较于其他季节, 采食难度较大[37-38, 40, 43, 47, 54, 56]。放牧家畜除在春季处于发情交配期, 活动强度均大于其他季节外, 食物资源的多样性(灌木、竹子和草本植物等)、采食策略的广谱性(嫩枝嫩叶、干草、老竹等), 以及人为干预的频繁性(饲喂食盐、添加辅料或转移采食场等)、活动空间的局域性(很少长距离迁徙、活动范围较为固定), 都不同程度地抑制了活动水平和活动时间的剧烈波动, 因而其不同月份或季节之间的绝对值相差远小于大熊猫[11, 23, 25](野外观察, 2010—2012)。
3.2 野生大熊猫与放牧家畜活动的连续性与持续性野生大熊猫的“分散多核域”活动模式、随季节和食物类型的变化而垂直移动(如邛崃山系的“撵笋”现象、秦岭山系的“夏居地”与“冬居地”之分), 以及低能耗的生理特性(能量消耗量为6.2 MJ/d)、微强度的活动格局(活动水平仅14.99或25.18、休息时间高达46%)(图 3、图 4), 从而形成了较高的活动时间片段率(1.62%)、较短的活动时长(77.63 min)和不同季节的变化规律(图 5、图 6)[23, 31, 37-38, 40-41, 43]。放牧家畜的活动时间片段率显著低于野生大熊猫(P < 0.0001), 但绝对值相差较小(1.40%), 活动片段时长也短(79.59 min)、且与大熊猫无明显的统计学意义(P=0.4107)(图 5、图 6), 表明放牧家畜的片段化程度依然很高, 这主要是放牧家畜营群体生活, 幼体与成体之间的体能和活动模式相差较大, 导致整个群体走走停停、活动与休息频繁交替。
野生大熊猫与放牧家畜在季节动态方面, 春季和夏秋季的规律一致, 片段率前者高于后者、片段时长前者小于后者;冬季两者之间与其他季节呈相反变化, 野生大熊猫片段率低于放牧家畜、片段时长大于后者(图 5、图 6)。分析原因, 可能是冬季竹子资源的数量和质量不及春季、夏秋季丰富与营养, 以及冰雪封冻, 致使野生大熊猫需要不停地走动寻找最佳食物、长距离移动至水源处饮水、持续采食以补充能量[37-38, 56-59]。从活动片段时长的不同尺度级分布图式来看(图 6), 野生大熊猫的小径级(1—6级)时间密度高于放牧家畜, 而大径级(7—9级)则明显为低, 显示大熊猫主要以小片段的活动时长为主, 这与其低能耗的生理特征相符[31]。
3.3 野生大熊猫与放牧家畜活动的相关性动物的活动格局虽受环境条件、生理阶段等因素影响[32-33, 36], 但主要是其遗传基因(生物钟的周期基因)所控制[26-28], 因而不同动物之间(除捕食者-被食者外)在活动格局方面的关联性是通过生境利用(食物和栖息地的资源竞争)、空间扩散或退缩等间接方式表现出来的[6-7, 11-12, 23-24]。况且野生大熊猫与放牧家畜(马)之间不属于捕食者-被食者关系, 两者具有相互回避、不直接发生冲突的策略, 因此, 野生大熊猫与放牧家畜之间对其各自的时间利用规律(活动格局)直接影响甚微。
野生动物或放牧家畜不同的空间利用格局对栖息地、食物资源和伴生动物的利用方式、影响强度、空间分布均截然相异, 比如放牧家畜(马群)“步步为营”的空间扩展模式不仅阻断了野生大熊猫的移动通道, 缩小了适宜栖息地的范围, 改变了伴生动物的空间分布格局, 而且导致栖息地生境结构组分的变化、食物资源的质量降低、主食竹的种群衰退, 甚至可能造成水土流失、疫病传染等生态后果[23-24]。从活动格局分析放牧家畜与野生大熊猫, 除了活动时间片段率外, 放牧家畜的其他指标都明显高于野生大熊猫, 尤其是活动强度和活动时间百分率, 都远远大于后者, 分别达到1.37倍和1.51倍, 这充分表明放牧家畜对大熊猫栖息地和主食竹的利用时间长、采食强度大、影响程度深[11, 24]。
致谢: 数据收集过程中得到卧龙特区卧龙镇杨文兵和杨洪先生的帮助, 中国大熊猫保护研究中心科学研究与动物管理处邓林华、李才武协助进行大熊猫麻醉和GPS颈圈的佩戴, 特此致谢。| [1] |
国家林业局. 全国第三次大熊猫调查报告. 北京: 科学出版社, 2006: 1-355.
|
| [2] |
中华人民共和国国务院新闻办公室.国家林业局举行全国第四次大熊猫调查结果新闻发布会. (2015-03-01)[2018-03-20] http://www.gov.cn/xinwen/2015-03/01/content_2823605.htm
|
| [3] |
唐小平, 贾建生, 王志臣, 张德辉, 遇宝成, 岳建兵, 龚明昊, 刘洋. 全国第四次大熊猫调查方案设计及主要结果分析. 林业资源管理, 2015(1): 11-16. |
| [4] |
Kang D W, Li J Q. Premature downgrade of panda's status. Science, 2016, 354(6310): 295. |
| [5] |
周世强, 屈元元, 黄金燕, 黄炎, 李德生, 张和民. 野生大熊猫种群动态的研究综述. 四川林业科技, 2017, 38(2): 17-30. |
| [6] |
冉江洪, 刘少英, 王鸿加, 孙治宇, 曾宗永, 刘世昌. 小相岭大熊猫与放牧家畜的生境选择. 生态学报, 2003, 23(11): 2253-2259. DOI:10.3321/j.issn:1000-0933.2003.11.007 |
| [7] |
冉江洪, 刘少英, 王鸿加, 孙治宇, 曾宗永, 刘世昌. 放牧对冶勒自然保护区大熊猫生境的影响. 兽类学报, 2003, 23(4): 288-294. DOI:10.3969/j.issn.1000-1050.2003.04.003 |
| [8] |
冉江洪, 刘少英, 王鸿加, 曾宗永, 孙治宇, 刘世昌. 小相岭大熊猫栖息地干扰调查. 兽类学报, 2004, 24(4): 277-281. DOI:10.3969/j.issn.1000-1050.2004.04.001 |
| [9] |
王学志.岷山地区人类活动干扰对大熊猫生境利用的影响研究[D].北京: 中国科学院生态环境研究中心, 2008. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1627194
|
| [10] |
康东伟, 赵联军, 宋国华. 四川王朗国家级自然保护区大熊猫与家畜竞争关系. 东北林业大学学报, 2011, 39(7): 74-76. DOI:10.3969/j.issn.1000-5382.2011.07.021 |
| [11] |
Hull V, Zhang J D, Zhou S Q, Huang J Y, Viňa A, Liu W, Tuanmu M N, Li R G, Liu D, Xu W H, Huang Y, Ouyang Z Y, Zhang H M, Liu J G. Impact of livestock on giant pandas and their habitat. Journal for Nature Conservation, 2014, 22(3): 256-264. DOI:10.1016/j.jnc.2014.02.003 |
| [12] |
四川省林业厅. 四川的大熊猫:四川省第四次大熊猫调查报告. 成都: 四川科学技术出版社, 2015: 129-159.
|
| [13] |
Steinfeld H, Gerber P, Wassenaar T, Castel V, Rosales M, de Haan C. Livestock's long shadow: environmental issues and options. Rome: Food and Agricultural Organization of the United Nations, 2006.[2018-03-20]. http://www.fao.org/docrep/010/a0701e/a0701e.pdf
|
| [14] |
Adler P, Raff D, Lauenroth W. The effect of grazing on the spatial heterogeneity of vegetation. Oecologia, 2001, 128(4): 465-479. DOI:10.1007/s004420100737 |
| [15] |
Madhusudan M D. Recovery of wild large herbivores following livestock decline in a tropical Indian wildlife reserve. Journal of Applied Ecology, 2004, 41(5): 858-869. DOI:10.1111/jpe.2004.41.issue-5 |
| [16] |
van Uytvanck J, Hoffmann M. Impact of grazing management with large herbivores on forest ground flora and bramble understorey. Acta Oecologica, 2009, 35(4): 523-532. DOI:10.1016/j.actao.2009.04.001 |
| [17] |
Bai Y F, Wu J G, Clark C M, Pan Q M, Zhang L X, Chen S P, Wang Q B, Han X G. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 2012, 49(6): 1204-1215. DOI:10.1111/jpe.2012.49.issue-6 |
| [18] |
Evans D M, Villar N, Littlewood N A, Pakeman R J, Evans S A, Dennis P, Skartveit J, Redpath S M. The cascading impacts of livestock grazing in upland ecosystems:a 10-year experiment. Ecosphere, 2015, 6(3): 1-15. |
| [19] |
Li W H, Zhan S X, Lan Z C, Wu X B, Bai Y F. Scale-dependent patterns and mechanisms of grazing-induced biodiversity loss:evidence from a field manipulation experiment in semiarid steppe. Landscape Ecology, 2015, 30(9): 1751-1765. DOI:10.1007/s10980-014-0146-4 |
| [20] |
Ren H Y, Taube F, Stein C, Zhang Y J, Bai Y F, Hu S J. Grazing weakens temporal stabilizing effects of diversity in the Eurasian steppe. Ecology and Evolution, 2018, 8(1): 231-241. DOI:10.1002/ece3.3669 |
| [21] |
Erb K H, Kastner T, Plutzar C, Bais A L S, Carvalhais N, Fetzel T, Gingrich S, Haberl H, Lauk C, Niedertscheider M, Pongratz J, Thurner M, Luyssaert S. Unexpectedly large impact of forest management and grazing on global vegetation biomass. Nature, 2018, 553(7686): 73-76. |
| [22] |
黄金燕, 刘巅, 张明春, 谢浩, 周世强, 李仁贵, 张晋东, 周小平, 黄炎, 李德生, 张和民. 放牧对卧龙大熊猫栖息地草本植物物种多样性与竹子生长影响. 竹子学报, 2017, 36(2): 57-64. DOI:10.3969/j.issn.1000-6567.2017.02.009 |
| [23] |
周世强, Hull V, 张晋东, 黄金燕, 刘巅, 黄炎, 李德生, 张和民. 野生大熊猫与放牧家畜的空间利用格局比较. 兽类学报, 2016, 36(2): 138-151. |
| [24] |
Li B V, Pimm S L, Li S, Zhao L J, Luo C P. Free-ranging livestock threaten the long-term survival of giant pandas. Biological Conservation, 2017, 216: 18-25. DOI:10.1016/j.biocon.2017.09.019 |
| [25] |
Zhang J D, Hull V, Ouyang Z Y, Li R G, Connor T, Yang H B, Zhang Z J, Silet B, Zhang H M, Liu J G. Divergent responses of sympatric species to livestock encroachment at fine spatiotemporal scales. Biological Conservation, 2017, 209: 119-129. DOI:10.1016/j.biocon.2017.02.014 |
| [26] |
Allada R, White N E, So W V, Hall J C, Rosbash M. A mutant Drosophila homolog of Mammalian clock disrupts circadian rhythms and transcription of Period and Timeless. Cell, 1998, 93(5): 791-804. DOI:10.1016/S0092-8674(00)81440-3 |
| [27] |
Dunlap J C. Molecular bases for circadian clocks. Cell, 1999, 96(2): 271-290. DOI:10.1016/S0092-8674(00)80566-8 |
| [28] |
韦芳, 王正荣. 哺乳动物的生物钟分子机制研究进展. 四川生理科学杂志, 2002, 24(1): 1-4. DOI:10.3969/j.issn.1671-3885.2002.01.001 |
| [29] |
孙儒泳. 动物生态学原理(第三版). 北京: 北京师范大学出版社, 2001: 91-98.
|
| [30] |
McNAB B K. Ecological factors influence energetics in the order Carnivora. Acta Zoologica Sinica, 2005, 51(4): 535-545. |
| [31] |
Nie Y G, Speakman J R, Wu Q, Zhang C L, Hu Y B, Xia M H, Yan L, Hambly C, Wang L, Wei W, Zhang J G, Wei F W. Exceptionally low daily energy expenditure in the bamboo-eating giant panda. Science, 2015, 349(6244): 171-174. DOI:10.1126/science.aab2413 |
| [32] |
Beier P, McCullough D R. Factors influencing white-tailed deer activity patterns and habitat use. Wildlife Monographs, 1990, 109: 3-51. |
| [33] |
Beltrán J F, Delibes M. Environmental determinants of circadian activity of free-ranging Iberian lynxes. Journal of Mammalogy, 1994, 75(2): 382-393. DOI:10.2307/1382557 |
| [34] |
Schmidt K. Variation in daily activity of the free-living Eurasian lynx (Lynx lynx) in Bialowieza Primeval Forest, Poland. Journal of Zoology, 1999, 249(4): 417-425. |
| [35] |
Kolbe J A, Squires J R. Circadian activity patterns of Canada lynx in western Montana. Journal of Wildlife Management, 2007, 71(5): 1607-1611. DOI:10.2193/2005-727 |
| [36] |
Garshelis D L, Pelton M R. Activity of black bears in the Great Smoky Mountains national park. Journal of Mammalogy, 1980, 61(1): 8-19. DOI:10.2307/1379952 |
| [37] |
胡锦矗, 乔治·夏勒, 潘文石, 朱靖. 卧龙的大熊猫. 成都: 四川科学技术出版社, 1985: 33-116.
|
| [38] |
Schaller G B, Hu J C, Pan W S, Zhu J. The Giant Pandas of Wolong. Chicago:University of Chicago Press, 1985, 47-107. |
| [39] |
雍严格, 王宽武, 汪铁军. 佛坪大熊猫的移动习性. 兽类学报, 1994, 14(1): 9-14. |
| [40] |
潘文石, 吕植, 朱小健, 王大军, 王昊, 龙玉, 付达莉, 周欣. 继续生存的机会. 北京: 北京大学出版社, 2001: 1-522.
|
| [41] |
Liu X H, Skidmore A K, Wang T J, Yong Y G, Prins H H T. Giant panda movements in Foping Nature Reserve, China. The Journal of Wildlife Management, 2002, 66(4): 1179-1188. DOI:10.2307/3802951 |
| [42] |
张晋东, Hull V, 黄金燕, 周世强, 李仁贵, 刘巅, 黄炎, 李德生, 徐卫华, 张和民, 刘建国, 欧阳志云. 大熊猫取食竹笋期间的昼夜活动节律和强度. 生态学报, 2011, 31(10): 2655-2661. |
| [43] |
Zhang J D, Hull V, Huang J Y, Zhou S Q, Xu W H, Yang H B, McConnell W J, Li R G, Liu D, Huang Y, Ouyang Z Y, Zhang H M, Liu J G. Activity patterns of the giant panda (Ailuropoda melanoleuca). Journal of Mammalogy, 2015, 96(6): 1116-1127. DOI:10.1093/jmammal/gyv118 |
| [44] |
Zhang Z J, Sheppard J K, Swaisgood R R, Wang G, Nie Y G, Wei W, Zhao N X, Wei F W. Ecological scale and seasonal heterogeneity in the spatial behaviors of giant pandas. Integrative Zoology, 2014, 9(1): 46-60. DOI:10.1111/inz2.2014.9.issue-1 |
| [45] |
Hull V, Zhang J D, Zhou S Q, Huang J Y, Li R G, Liu D, Xu W H, Huang Y, Ouyang Z Y, Zhang H M, Liu J G. Space use by endangered giant pandas. Journal of Mammalogy, 2015, 96(1): 230-236. DOI:10.1093/jmammal/gyu031 |
| [46] |
卧龙自然保护区管理局, 南充师范学院生物系, 四川省林业厅保护处. 卧龙植被及资源植物. 成都: 四川科学技术出版社, 1987: 11-194.
|
| [47] |
周世强, 黄金燕, 张亚辉, 李德生, 黄炎, 周小平, 王鹏彦, 张和民. 卧龙自然保护区大熊猫栖息地植物群落多样性Ⅴ:不同竹林的物种多样性. 应用与环境生物学报, 2009, 15(2): 180-187. |
| [48] |
Huang J, Li Y Z, Du L M, Yang B, Shen F J, Zhang H M, Zhang Z H, Zhang X Y, Yue B S. Genome-wide survey and analysis of microsatellites in giant panda (Ailuropoda melanoleuca), with a focus on the applications of a novel microsatellite marker system. BMC Genomics, 2015, 16(1): 61. DOI:10.1186/s12864-015-1268-z |
| [49] |
Hull V, Zhang J D, Huang J Y, Zhou S Q, Viña A, Shortridge A, Li R G, Liu D, Xu W H, Ouyang Z Y, Zhang H M, Liu J G. Habitat Use and Selection by Giant Pandas. PLoS One, 2016, 11(9): e0162266. DOI:10.1371/journal.pone.0162266 |
| [50] |
Coulombe M L, Massé A, Côté S D. Quantification and accuracy of activity data measured with VHF and GPS telemetry. Wildlife Society Bulletin, 2006, 34(1): 81-92. DOI:10.2193/0091-7648(2006)34[81:QAAOAD]2.0.CO;2 |
| [51] |
张红兵, 贾来喜, 李潞. SPSS宝典. 北京: 电子工业出版社, 2007: 25-116.
|
| [52] |
叶卫平. Origin 9.1科技绘图及数据分析. 北京: 机械工业出版社, 2015: 1-462.
|
| [53] |
周小平, 王鹏彦, 张和民, 魏荣平, 张贵权, 李德生, 黄炎, Ronald Swaisgood. 圈养大熊猫行为研究及其方法. 成都: 四川科学技术出版社, 2005: 1-23.
|
| [54] |
黄金燕, 周世强, 谭迎春, 周小平, 王鹏彦, 张和民. 卧龙自然保护区大熊猫栖息地植物群落多样性研究:丰富度、物种多样性指数和均匀度. 林业科学, 2007, 43(3): 73-78. |
| [55] |
周世强, 张和民, 李德生. 大熊猫觅食行为的栖息地管理策略. 四川动物, 2010, 29(3): 340-345. |
| [56] |
Li Y X, Swaisgood R R, Wei W, Nie Y G, Hu Y B, Yang X Y, Gu X D, Zhang Z J. Withered on the stem:is bamboo a seasonally limiting resource for giant pandas?. Environmental Science and Pollution Research, 2017, 24(11): 10537-10546. DOI:10.1007/s11356-017-8746-6 |
| [57] |
Zhang Z J, Zhan X J, Yan L, Li M, Hu J C, Wei F W. What determines selection and abandonment of a foraging patch by wild giant pandas (Ailuropoda melanoleuca) in winter. Environmental Science and Pollution Research, 2009, 16(1): 79-84. DOI:10.1007/s11356-008-0066-4 |
| [58] |
李亚军, 蔡琼, 刘雪华, Songer M, 武鹏峰, 贾晓东, 何祥博. 海拔对大熊猫主食竹结构、营养及大熊猫季节性分布的影响. 兽类学报, 2016, 36(1): 24-35. |
| [59] |
王丹林, 郭庆学, 王小蓉, 梁春平, 张远彬. 海拔对岷山大熊猫主食竹营养成分和氨基酸含量的影响. 生态学报, 2017, 37(19): 6440-6447. |