 2019, Vol. 39
2019, Vol. 39文章信息
- 薛欣欣, 吴小平, 王文斌, 罗雪华, 张永发, 王大鹏, 赵春梅
- XUE Xinxin, WU Xiaoping, WANG Wenbin, LUO Xuehua, ZHANG Yongfa, WANG Dapeng, ZHAO Chunmei
- 坡度和埋深对橡胶林凋落叶分解及红外光谱特征的影响
- Effects of varying slopes and depths on decomposition and infrared spectrum characteristics of leaf litter in a rubber forest
- 生态学报. 2019, 39(3): 874-883
- Acta Ecologica Sinica. 2019, 39(3): 874-883
- http://dx.doi.org/10.5846/stxb201804070781
-
文章历史
- 收稿日期: 2018-04-07
- 网络出版日期: 2018-11-15





2. 中国热带农业科学院土壤肥料研究中心, 儋州 571737
2. Soil and Fertilizer Center, Chinese Academy of Tropical Agriculture Science, Danzhou 571737, China
植株凋落物的分解是土壤有机质形成、有机养分的矿化以及维持碳平衡的关键过程[1]。森林植被每年从土壤中吸收的养分物质50%以上以枯枝落叶形式归还给土壤, 充分利用森林凋落物, 实现森林的自肥作用, 在森林土壤的培肥与管理上具有十分重要的意义[2]。橡胶林生态系统作为一个开放的人工森林系统, 是我国热带地区主要的人工林生态系统类型之一, 每年有大量的枯枝枯叶凋落, 进而在该系统中分解、循环。研究表明, 在橡胶树33年的生产周期内, 通过枯枝落叶的分解向土壤归还的氮、磷、钾、镁量分别为1350、90、300、270 kg/hm2[3]。可见, 凋落物在橡胶林生态系统养分循环中占较大比重。橡胶树作为阔叶高大乔木, 通常在每年的2—3月份落叶而换发新叶, 研究凋落叶在橡胶树全周期生产中的养分循环过程及其影响因素对指导橡胶林合理施肥具有重要的实践意义。
我国橡胶林种植地型多变、气候环境复杂, 土壤动物、立地条件、温度、大气沉降、植被类型等因素均可能对凋落物分解产生一定的影响。研究表明, 凋落物分解受到土壤微生物生活代谢的影响较大, 土壤中的环境条件越利于微生物生长则凋落物的分解就越迅速, 而土壤微生物活性在特定环境下与所处的深度密切相关[4]。不同立地条件下土壤水分、径流强度均有差异, 也会对凋落物的分解产生较大的影响。研究表明, 适宜的土壤水分可提高分解者的活性, 调控微生物氧活性, 并可能通过破坏有机质结构为微生物提供可利用的有机碳[5]。另外, 坡度大小也会影响地表径流及凋落物的积累进而导致矿质养分的流失或聚集[6]。目前研究橡胶林凋落物分解方式多数集中在地表覆盖, 而忽视了立地条件等因素[7]。再者, 凋落物在土壤中分解转化所形成的中间产物是非常复杂的, 并且很难分离出来, 而红外光谱技术则能够区分个体结构上的差异, 同时具有操作简便及快速检测等特点, 可用于作物残体结构的变化。将傅里叶变换红外光谱(fourier transform infrared spectroscopy, FTIR)应用于农业研究是近些年来关注的热点, 探索森林生态系统凋落叶在土壤中分解的结构变化, 明确凋落物归还后分解变化机制, 有利于为森林生态系统的物质循环提供理论参考[5, 8-9]。
本研究拟采用尼龙网袋原位分解法, 探究橡胶林凋落叶在不同坡度及埋深条件下的干物质分解、养分释放特性, 同时运用FTIR技术, 定性分析凋落叶分解前后的组分及结构变化特征, 为橡胶林生态系统的物质循环深入研究及指导科学施肥提供理论基础。
1 材料与方法 1.1 试验点概况试验点位于海南省儋州市中国热带农业科学院试验场, 地处109°49′E, 19°48′N。该地区处于东亚大陆季风气候的南缘, 属热带湿润季风气候, 5—10月为雨季, 11月—次年4月为干季, 年均日照时数2000 h以上, 年均气温为23.5℃, 年均降雨量为1623 mm(图 1)。该地区由丘陵、平原和山地三部分构成, 丘陵占76.50%, 平原占23.13%, 山地占0.37%, 海拔大部分在200 m以下。土壤类型为花岗岩发育的砖红壤, 主要人工林植被类型为天然橡胶林。

|
| 图 1 近20年儋州市平均月降雨量和月气温 Fig. 1 Average monthly rainfall and temperature in Danzhou for recently 20 years |
2017年3月(橡胶树落叶期)在试验区林地收集橡胶树自然凋落叶作为研究对象, 将凋落叶于室内风干备用, 凋落叶分解采用凋落袋法。将凋落叶剪成1 cm ×1 cm的片段, 准确称取10 g, 分别装进10 cm(宽)×20 cm(长)、孔径1 mm2的尼龙网袋中[10]。随机称取5份重量为20 g的风干叶片, 放于烘箱中70℃烘干72 h至恒重, 计算风干系数。
试验从2017年4月1日开始, 选择平地和坡地(坡度约20°)相邻的两块管理一致的橡胶园, 橡胶树栽培密度为3 m×7 m, 两块试验样地面积分别约为200 m2。尼龙网袋均匀置于距每棵树干1 m的位置, 如图 2所示。试验设4个处理:(I)平地胶园地表覆盖(F-Ⅰ), (2)平地胶园埋深10 cm(F-Ⅱ), (3)坡地胶园地表覆盖(S-Ⅰ), (4)坡地胶园埋深10 cm(S-Ⅱ), 每个处理3次重复, 共计96袋;其中地表覆盖处理是将尼龙网袋固定于地表并与土壤平行接触, 并用凋落叶将其覆盖;埋深处理是将尼龙网袋平放于挖好的坑穴后回填原土, 坑穴的长、宽、深分别为15、25、10 cm。试验开始后分别于15、30、60、90、120、150、210、240 d共8次进行采样, 每次每处理各取回3袋, 带回实验室, 将杂物和土块用手挑出, 随后用自来水轻轻冲洗尼龙网袋表面及内部的泥土, 待网袋及凋落叶附着泥土风干后, 用毛刷将其去除干净, 随后将凋落叶装于纸质信封袋中置于70℃烘箱中烘干至恒重, 记录烘干重。烘干样品用陶瓷研钵进行磨细、过筛后用于养分元素含量测定及傅里叶变换红外光谱(fourier transform infrared spectroscopy, FTIR)分析。

|
| 图 2 试验处理示意图 Fig. 2 The diagram of experimental treatments F-Ⅰ:平地胶园地表覆盖处理, Treatment of covered flatland;F-Ⅱ:平地胶园埋深处理, Treatment of buried flatland;S-Ⅰ:坡地胶园地表覆盖处理, Treatment of covered slopes;S-Ⅱ:坡地胶园埋深处理, Treatment of buried slopes |
凋落叶全碳(C)采用重铬酸钾外加热法, 氮(N)和磷(P)均采用H2SO4-H2O2消煮-连续流动分析仪法, 钾(K)采用H2SO4-H2O2消煮-火焰光度计法, 钙(Ca)和镁(Mg)均采用H3ClO4-HNO3消煮-原子吸收分光光度计法[11]。橡胶树凋落叶初始化学性质如表 1所示。
| C/% | N/% | C/N | P/% | K/% | Ca/% | Mg/% | |
| 凋落叶Leaf litter | 43.81 | 1.75 | 25.1 | 0.114 | 1.207 | 1.256 | 0.170 |
凋落叶FTIR分析方法:将样品过100目筛, 在105℃烘箱中烘干恒重后, 在干燥条件下, 放入玛瑙研钵中, 同时加入烘干的已去除结晶水的KBr(光谱纯级)适量, 在红外灯下混匀后反复磨细呈淀粉状, 取少许在压片机上压成透明的薄片, 真空条件下, 用压杆缓慢加压至约为15 Mpa, 维持1 min, 之后将薄片放在Nicolet公司的Inpact-410型FTIR光谱仪上进行测定, 测定区域4000—900 cm-1, 扫描次数32次, 分辨率4 cm-1。
1.4 数据分析[10](1) 凋落叶干物质残留率(%)
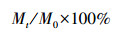
|
式中, Mt为t时刻的凋落叶剩余干重(g), M0为凋落叶初始干重(g)。
(2) 凋落叶分解速率(k)
根据Olsen衰减模型模拟凋落叶分解过程, 并计算凋落叶分解系数(k)
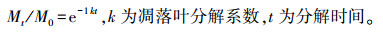
|
(3) 元素残留率(R)
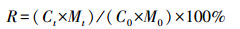
|
式中, Ct为t时刻的凋落叶养分元素含量(%), C0为初始养分元素含量(%)。
1.5 数据分析采用Omnic 8.0软件对FTIR谱图进行数据处理, Origin 8.0软件绘图, SPSS 20.0软件进行统计分析, LSD法进行多重比较。
2 结果与分析 2.1 坡度和埋深对凋落叶干物质残留率和C/N比的影响图 3显示, 随分解时间推进, 各处理的凋落叶干物质残留率呈逐渐下降趋势。分解初始到2个月期间, 地表覆盖处理F-Ⅰ、S-Ⅰ均大于埋深处理F-Ⅱ、S-Ⅱ;从分解3—9个月, 各处理的干物质残留率大小顺序表现为:坡地地表覆盖处理S-Ⅰ>平地地表覆盖处理F-Ⅰ>坡地埋深处理F-Ⅱ、平地埋深处理S-Ⅱ。凋落叶分解9个月后, F-Ⅱ、F-Ⅱ、S-Ⅰ和S-Ⅱ的干物质残留率分别为26.8%、11.2%、39.6%、6.9%。总体来看, 埋深处理的凋落叶干物质残留率较地表覆盖处理快。方差分析显示, 各时期处理间干物质残留率差异显著(P < 0.05)。

|
| 图 3 不同坡度和埋深条件下橡胶树凋落叶干物质残留率和碳氮比的动态变化 Fig. 3 Variation of dry mass remaining rates and C/N in varying slopes and depths for rubber leaf litters *表示同一时间不同处理间差异显著(P < 0.05) |
利用Olsen指数衰减模型对凋落叶干物质分解动态进行拟合, 相关系数R2均达极显著水平(P < 0.01)(表 2)。方程中分解系数k值的生态学意义是k值越大, 凋落叶的分解速率越快。由表 2可以看出各处理的分解常数间差异显著(P < 0.05), 埋深处理F-Ⅱ、S-Ⅱ显著高于地表覆盖处理F-Ⅰ、S-Ⅰ, 而地表覆盖条件下则平地F-Ⅰ明显大于坡地S-Ⅰ。凋落叶干物质分解50%和95%所需要的时间以坡地覆盖处理S-Ⅰ最长, 而埋深处理F-Ⅱ、S-Ⅱ所需要的时间均较短。
| 处理 Treatment |
Olsen指数模型 Olsen exponential model |
相关系数 Correlation coefficient(R2) |
分解常数 Decomposition coefficient(k) |
分解50%所需时间 Time required for leaf litter decomposition 50%/a |
分解95%所需时间 Time required for leaf litter decomposition 95%/a |
| F-Ⅰ | Mt/M0=e-0.1464t | 0.9412 | 0.1464 b | 0.39 | 1.71 |
| F-Ⅱ | Mt/M0= e-0.2341t | 0.9166 | 0.2341 a | 0.25 | 1.07 |
| S-Ⅰ | Mt/M0 = e-0.1024t | 0.8777 | 0.1024 c | 0.57 | 2.44 |
| S-Ⅱ | Mt/M0=e-0.2270t | 0.9204 | 0.2270 a | 0.26 | 1.10 |
| t:分解月份t-decomposition time, k:年分解常数k Annual decomposition rate, Mt/M0:凋落叶干物质残留率Dry mass remaining rates of leaf litter, Mt:凋落叶剩余干重Residual dry mass weight of litter leaf, M0:凋落叶初始干重Initial dry mass weight of litter leaf;数据后不同字母表示不同处理间差异显著(P < 0.05) | |||||
凋落叶C/N比也随分解时间推进呈波动性降低的趋势(图 3)。各处理相比, 4个月前坡地覆盖处理S-Ⅰ的C/N比明显高于其他处理(P < 0.05), 而4个月后处理间差异不显著(P < 0.05)。F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ凋落叶C/N比从最初始的25.1下降到9个月后的14.4、16.2、12.7、16.9。
2.2 坡度和埋深对凋落叶分解过程中C、N含量及其残留率的影响由图 4可知, 凋落叶C含量随分解时间推进均呈波动性降低的趋势, 表现为0—0.5个月增加, 0.5—4个月急剧下降, 4—5个月增加, 5—9个月缓慢下降。以坡地埋深处理S-Ⅱ的C含量始终最高, 而平地覆盖处理F-Ⅰ的C含量始终最低, 在分解1、2、4、5、7、9个月时处理间差异显著(P < 0.05)。C残留率随分解时间推进均呈逐渐下降的趋势, 坡地覆盖处理S-Ⅰ的C残留率高于其他处理, 方差分析表明, 1—9个月间, 处理间差异均达显著水平(P < 0.05);分解9个月后, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ处理C残留率分别为6.4%、5.9%、10.9%和3.8%。

|
| 图 4 不同坡度及埋深条件下橡胶树凋落叶C、N含量及其残留率的动态变化 Fig. 4 Variation of C, N contents and their remaining rate under varying slopes and depths for rubber leaf litters |
凋落叶N含量的变化在2个月前呈现增加趋势, 2—9个月呈持续降低的趋势;埋深处理F-Ⅱ和S-Ⅱ的氮含量均高于覆盖处理F-Ⅰ和S-Ⅰ, 处理间差异显著(P < 0.05)。N残留率随分解时间推进均呈下降趋势, 坡地覆盖处理S-Ⅰ的N残留率高于其他处理, 方差分析表明, 处理间差异显著(P < 0.05);分解9个月后, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ处理N残留率分别为11.8%、9.1%、21.6%和6.5%。
2.3 坡度及埋深对凋落叶分解过程中P、K、Ca、Mg含量及其残留率的影响由图 5可知, 凋落叶P含量随分解时间的推进, 各处理变化有所不同, 其中坡地覆盖处理S-Ⅰ在0.5个月前呈降低的趋势, 0.5—4个月呈持续上升的趋势, 而埋深处理F-Ⅱ和S-Ⅱ在0.5个月前均呈上升的趋势, 0.5—4个月呈降低的趋势, 各处理在4—9个月均呈持续降低的趋势;分解期间, 埋深处理F-Ⅱ、S-Ⅱ的P含量均高于覆盖处理F-Ⅰ、S-Ⅰ, 方差分析显示处理间差异显著(P < 0.05)。凋落叶P残留率随分解时间推进变化趋势与P含量基本一致, 分解3个月后的坡地覆盖处理S-Ⅰ的残留率明显高于坡地埋深处理S-Ⅱ, 处理间差异显著(P < 0.05)。分解9个月后, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ处理P残留率分别为5.7%、5.3%、10.7%和3.4%。

|
| 图 5 不同坡度及埋深条件下橡胶树凋落叶P、K、Ca、Mg含量及其残留率的变化 Fig. 5 Variation of P, K, Ca and Mg contents in varying slopes and depths for rubber leaf litters |
凋落叶K含量在2个月前呈急剧降低的趋势, 2—4个月呈缓慢上升的趋势, 4—9个月呈缓慢下降的趋势, 9个月之前处理间差异显著(P < 0.05)。凋落叶K残留率与K含量变化趋势相似, 分解期间以坡地地表覆盖处理S-Ⅰ的K残留率最高, 在0—2个月急剧下降到较低水平, 分解2个月时, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ各处理K残留率分别为29.3%、19.5%、32.7%和17.1%;分解9个月后, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ各处理K残留率分别仅为4.8%、3.1%、9.7%和2.3%。处理间差异显著(P < 0.05)。
凋落叶Ca含量随分解时间的推进, 覆盖和埋深处理的变化有所差别, 平地覆盖处理F-Ⅰ和平地埋深处理F-Ⅱ均呈持续降低的趋势, 而坡地覆盖处理S-Ⅰ和坡度埋深处理S-Ⅱ均呈先增加后降低的趋势, 在分解2个月时达到最大值, 之后呈持续降低的趋势, 其中坡地埋深处理S-Ⅱ在2个月后由各处理中最高水平急剧下降到最低水平;方差分析显示, 处理间差异显著(P < 0.05)。凋落叶Ca的残留率以坡面覆盖处理S-Ⅰ较高, 坡面埋深处理S-Ⅱ最低, 方差分析表明, 处理间差异均显著(P < 0.05)。分解9个月后, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ各处理Ca残留率分别为5.5%、1.4%、10.4%和0.8%。
凋落叶Mg含量随分解时间的推进呈上升—下降的趋势, 在分解0.5个月时达到最大值;分解第9个月时处理间无显著差异(P < 0.05)。就凋落叶Mg残留率而言, 坡地条件下的Mg残留率呈上升—下降的趋势, 在0.5个月时达到最大, 而平地条件下的Mg残留率则呈持续下降的趋势, 坡地覆盖处理S-Ⅰ的残留率高于其他处理。方差分析表明, 整个分解过程中处理间差异显著(P < 0.05)。分解9个月后, F-Ⅰ、F-Ⅱ、S-Ⅰ和S-Ⅱ各处理Mg残留率分别为4.7%、3.4%、7.9%和2.1%。
2.4 坡度及埋深对凋落叶红外光谱特征的影响凋落叶分解前后的傅里叶变换红外光谱特征(FTIR)如图 6所示。FTIR吸收峰的归属如下:红外光谱吸收峰发生变化的主要有3387 cm-1(一部分为碳水化合物中—OH形成的氢键的伸缩振动, 另一部分为纤维素、半纤维素、淀粉及其他多糖和单糖等的成分, 也包括氨基酸中N—H伸缩振动);2920 cm-1和2852 cm-1(分别为对称和非对称的油脂中—CH伸缩振动, 主要是膜脂和细胞壁果胶中常见的酯类化合物, 反映膜透性);1735 cm-1(酯化果胶中酯基—COOR的伸缩振动);1638 cm-1(细胞壁蛋白质上的C=O酰胺(ⅳ带)伸缩振动, 反映蛋白构象);1512 cm-1(蛋白质上N-H酰胺基(ⅶ带)的伸缩振动, 酰胺化合物的特征吸收峰);1439 cm-1(木质素和碳水化合物中饱和C—H的弯曲振动);1375 cm-1(具有脂肪族特征化合物中—CH3的对称变形振动, 说明存在纤维素);1153 cm-1(C—O的伸缩振动, 蛋白质分子氨基酸残基、纤维素糖苷等多糖吸收峰);1033 cm-1和1050 cm-1(碳水化合物和多糖结构中的C—O伸缩振动)。

|
| 图 6 不同坡度及埋深条件下橡胶树凋落叶FTIR(Fourier transform infrared spectroscopy)光谱特征 Fig. 6 FTIR (Fourier transform infrared spectroscopy) spectra of litter leaf initial and after decomposition in varying slopes and depths |
与分解初始相比, 分解9个月后的3387 cm-1处吸收峰强度有不同程度减弱, 表明橡胶树凋落叶中的纤维素、半纤维素、多糖等碳水化合物得到分解, 使—OH基团含量减少, 凋落叶中的残留C量也随之显著减少;各处理相比较, 坡地埋深处理S-Ⅱ的吸收峰最弱, 说明其分解速率较其他处理快。2920 cm-1、2853 cm-1在分解后吸收峰强度有所减弱, 说明分解后凋落叶细胞壁中的蛋白质、纤维素和果胶等组分有所减少;各处理相比较, F-Ⅰ的吸收峰最强, S-Ⅱ的吸收峰较弱。1734 cm-1处的吸收峰在分解后基本消失, 说明羧酸酯类化合物基本分解。1638 cm-1处的吸收峰强度有所减弱, 说明细胞壁中C=O基含量减少, 蛋白质含量降低;各处理相比较, 平地吸收峰强度强于坡地、覆盖强于埋深。1439、1375 cm-1处的吸收峰在分解后消失, 即木质素C—H弯曲振动和具脂肪族特征的化合物中—CH3的对称变角振动消失, 表明橡胶树凋落叶中木质素、纤维素、半纤维素或具脂肪族特征的化合物产生了分解。1050 cm-1处的吸收峰向低频方向位移了17 cm后变为1033 cm-1, 说明凋落物在分解过程中凋落叶原有的可溶性糖和纤维素C—C键和C—O键伸缩振动遭到破坏;各处理相比较, 埋深处理F-Ⅱ和S-Ⅱ的吸收峰较F-Ⅰ和S-Ⅰ处理弱, 也说明埋深处理加速了凋落叶的分解, 碳残留碳量随之降低。1153 cm-1处的吸收峰消失, 说明蛋白质分子氨基酸残基、纤维素糖苷等多糖降解。
3 讨论 3.1 埋放位置对凋落叶干物质分解速率的影响本研究表明, 凋落叶埋深处理的干物质分解速率明显比地表覆盖快;Olsen指数方程模拟显示, 埋深处理分解50%和95%所需要的时间比地表覆盖处理快1.7—3.7个月和7.7—16.1个月。有研究表明, 埋深处理明显加速了杉木凋落叶分解, 主要因为地下土壤水热条件要好于地表, 有利于微生物活动分解[12];金龙等[13]认为埋于橡胶林地下的凋落叶与土壤接触面积更大, 土壤生物参与凋落叶分解的机会也更大;陈晓丽等[14]研究表明, 随土层深度加深, 过氧化氢酶活性增大, 进而促进凋落物的分解。本研究结果还显示, 平地覆盖处理的凋落叶干物质分解速率比坡地覆盖处理快, 与吕瑞恒等[15]研究结果相似, 其研究表明, 坡度越小, 越有利于凋落物的分解。海拔、坡度的变化导致林下水热条件具有不同程度的差异, 进而影响凋落物分解速率。本研究还认为, 坡度主要影响土壤水分, 坡度越大, 土壤地表保水能力越差, 越不利于微生物的活动, 从而不利于凋落物的分解。
3.2 埋放位置对凋落叶养分元素分解的影响本研究表明, 凋落叶分解过程中C和N含量呈波动性下降的趋势, 但其残留率呈持续下降的趋势。该现象说明, 分解过程中凋落叶对C和N的富集作用较弱。赵春梅等[7]研究表明, 叶片氮的富集出现在4月份, 富集率仅为2.75%, 其余时间几乎呈下降的趋势;也可以说明, 枯落叶在分解过程中释放的C、N对土壤的补充贡献较高。王春阳等[16]研究表明, 凋落物可显著提高土壤微生物碳、氮含量。坡地覆盖处理S-Ⅰ的C、N残留率均较高, 埋深处理均较低, 说明埋深处理加快了营养元素的释放速率, 与吕瑞恒等[14]的研究结果相似。与C、N分解有所不同的是, P、K、Ca、Mg矿质营养元素在分解过程中均呈“释放-富集”模式, 富集现象出现时间分别为:埋深处理的P在0.5个月和3—4个月之间, 所有处理的K在4个月时, 坡地覆盖处理S-Ⅰ的Ca在2个月时, 坡地条件下的Mg在0.5个月时。各营养元素的富集率均较低且持续时间均较短, 总体均呈显著释放的趋势。K含量急剧下降, 之后趋于平缓, 这与K在植物体内主要以离子态存在形式有很大关系[17]。各营养元素的分解速率均以坡地覆盖处理S-Ⅰ最慢且残留量最大, 以埋深处理均较快且残留量较小, 造成该现象的主要原因与坡地覆盖处理干物质分解速率慢而埋深处理干物质分解较快有较大的关系。
3.3 埋放位置对凋落叶红外光谱特征的影响以往的研究表明, 油菜秸秆腐解过程中最明显的变化在3430—3410、2930 cm-1处, 其吸收峰强度减弱, 脂族性下降[5]。王文全等[18]研究表明, 随时间推移, 牛粪腐解在3430、2925 cm-1和2855 cm-1处的吸收峰逐渐减弱, 说明在分解过程中碳水化合物、脂肪族和蛋白质等有机物逐渐在分解。本试验结果表明, 分解270 d之后, 在3387、2920、2853、1439 cm-1和1375 cm-1处的吸收峰有所减弱甚至消失, 说明凋落叶碳水化合物、脂肪族碳、蛋白质等进行了部分的分解, 也应证了凋落叶在分解后全C含量及C/N大幅下降的原因。主要官能团FTIR中反映细胞壁多糖信息的是指纹区900—1200 cm-1(糖链的特征峰)。本研究表明, 在1033 cm-1处的吸收峰强度均表现为地表覆盖强于埋深处理, 也说明, 埋深加快了凋落叶碳水化合物的分解。因此, 在橡胶林生态系统中, 可以采取适当的或局部的将凋落物与表土掺混, 以加强土壤微生物的分解活动, 促进凋落物的分解, 加速养分的释放, 供橡胶树根系吸收利用。
4 结论橡胶树凋落叶干物质残留率随分解时间的推进符合Olsen指数衰减模型, 相关系数达极显著水平(P < 0.01)。凋落叶干物质和养分元素分解速率均受坡度和埋深处理显著影响(P < 0.05), 埋深处理较地表覆盖处理加快了凋落叶干物质分解速率;经过9个月的分解, 坡地覆盖处理S-Ⅰ的凋落叶C、N、P、K、Ca和Mg的元素残留率均较其他处理高, 而埋深处理F-Ⅱ和S-Ⅱ的元素残留率均较低。傅里叶红外光谱分析显示凋落叶分解后的碳水化合物、果胶、蛋白质、木质素等含量不同程度减少, 羧酸酯类化合物基本分解, 埋深处理分解程度均较覆盖处理高。综上所述, 凋落叶分解过程中, 平地覆盖较坡地覆盖分解快, 而埋深处理较覆盖处理分解快。建议橡胶生产中采取凋落叶与地表土掺混或压青的农艺措施以加快凋落叶的分解, 提高养分循环效率, 同时还能降低养分流失及森林火灾风险。
| [1] |
Austin A T, Ballaré C L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(10): 4618-4622. DOI:10.1073/pnas.0909396107 |
| [2] |
张鹏.亚热带森林土壤分解酶活性的季节变化及凋落物分解的研究[D].南京: 南京大学, 2007.
|
| [3] |
曹建华, 蒋菊生, 赵春梅, 陈俊明. 橡胶林生态系统养分循环研究进展. 热带农业科学, 2007, 27(3): 48-56. DOI:10.3969/j.issn.1009-2196.2007.03.015 |
| [4] |
李海涛, 于贵瑞, 李家永, 陈永瑞, 梁涛. 亚热带红壤丘陵区四种人工林凋落物分解动态及养分释放. 生态学报, 2007, 27(3): 898-908. DOI:10.3321/j.issn:1000-0933.2007.03.009 |
| [5] |
王景, 魏俊岭, 章力干, 常江, 郜红建. 厌氧和好气条件下油菜秸秆腐解的红外光谱特征研究. 中国生态农业学报, 2015, 23(7): 892-899. |
| [6] |
Koarashi J, Atarashi-Andoh M, Takeuchi E, Nishimura S. Topographic heterogeneity effect on the accumulation of Fukushima-derived radiocesium on forest floor driven by biologically mediated processes. Scientific Reports, 2014, 4: 6853. |
| [7] |
赵春梅, 曹建华, 李晓波, 兰国玉, 彭宗波, 蒋菊生. 橡胶林枯落物分解及其氮素释放规律研究. 热带作物学报, 2012, 33(9): 1535-1539. DOI:10.3969/j.issn.1000-2561.2012.09.002 |
| [8] |
曹莹菲, 张红, 赵聪, 刘克, 吕家珑. 秸秆腐解过程中结构的变化特征. 农业环境科学学报, 2016, 35(5): 976-984. |
| [9] |
Soong J L, Calderón F J, Betzen J, Cotrufo M F. Quantification and FTIR characterization of dissolved organic carbon and total dissolved nitrogen leached from litter:a comparison of methods across litter types. Plant and Soil, 2014, 385(1/2): 125-137. |
| [10] |
文海燕, 傅华, 郭丁. 黄土高原典型草原优势植物凋落物分解及养分释放对氮添加的响应. 生态学报, 2017, 37(6): 2014-2022. |
| [11] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2005.
|
| [12] |
马祥庆, 刘爱琴, 何智英, 俞立煊. 杉木幼林生态系统凋落物及其分解作用研究. 植物生态学报, 1997, 21(6): 564-570. DOI:10.3321/j.issn:1005-264X.1997.06.009 |
| [13] |
金龙, 吴志祥, 杨川, 管利民, 赖华英. 不同林龄橡胶凋落物叶分解特性与有机碳动态研究. 热带作物学报, 2015, 36(4): 698-705. DOI:10.3969/j.issn.1000-2561.2015.04.011 |
| [14] |
陈晓丽, 王根绪, 杨燕, 杨阳. 山地森林表层土壤酶活性对短期增温及凋落物分解的响应. 生态学报, 2015, 35(21): 7071-7079. |
| [15] |
吕瑞恒, 李国雷, 刘勇, 贾黎明, 江萍, 林娜. 不同立地条件下华北落叶松叶凋落物的分解特性. 林业科学, 2012, 48(2): 31-37. |
| [16] |
王春阳, 周建斌, 夏志敏, 刘瑞. 黄土高原区不同植物凋落物搭配对土壤微生物量碳、氮的影响. 生态学报, 2011, 31(8): 2139-2147. |
| [17] |
何洁, 杨万勤, 倪祥银, 李晗, 徐李亚, 吴福忠. 雪被斑块对川西亚高山森林凋落物冬季分解过程中钾和钠动态的影响. 植物生态学报, 2014, 38(6): 550-561. |
| [18] |
王文全, 赵秀玲, 罗艳丽, 施宠, 马荷, 余雄, 贾宏涛. 牛粪发酵过程中的红外光谱分析. 中国牛业科学, 2011, 37(2): 15-19. DOI:10.3969/j.issn.1001-9111.2011.02.004 |