 2019, Vol. 39
2019, Vol. 39文章信息
- 王媚臻, 毕浩杰, 金锁, 刘佳, 刘宇航, 王宇, 齐锦秋, 郝建锋
- WANG Meizhen, BI Haojie, JIN Suo, LIU Jia, LIU Yuhang, WANG Yu, QI Jinqiu, HAO Jianfeng
- 林分密度对云顶山柏木人工林林下物种多样性和土壤理化性质的影响
- Effects of stand density on understory species diversity and soil physicochemical properties of a Cupressus funebris plantation in Yunding Mountain
- 生态学报. 2019, 39(3): 981-988
- Acta Ecologica Sinica. 2019, 39(3): 981-988
- http://dx.doi.org/10.5846/stxb201803170528
-
文章历史
- 收稿日期: 2018-03-17
- 网络出版日期: 2018-11-15





2. 四川农业大学木材工业与家具工程重点实验室, 成都 611130;
3. 四川农业大学水土保持与荒漠化防治重点实验室, 成都 611130
2. Key Laboratory of Wood Industry and Furniture Engineering, Sichuan Agricultural University, Chengdu 611130, China;
3. Key Laboratory of Soil and Water Conservation and Desertification Control, Sichuan Agricultural University, Chengdu 611130, China
柏木林是川中丘陵区主要森林类型之一, 为亚热带常绿针叶林[1], 在气候调节、水土保持和生态恢复等方面有重要功能[2-3]。川中地区大部分柏木人工林因初始高密度连片栽植, 导致乔木层树种单一, 林分结构简单, 林下生物多样性低, 故为低产低效林[4-5]。林下植被是人工林生态系统的重要组成[6-7], 在促进养分循环与能量流动、林地生产力恢复、物种多样性保护等方面有关键作用[8-10], 是反映生态系统稳定性及群落演替规律的重要指标[7]。近年来, 关于人工林林下植被的研究多集中在杉木[11]、桉树[12]、马尾松[13]、油松[7]树种, 对于柏木人工林林下植被的相关研究报道较少, 关于林下植被的研究结果也不尽相同。
林分密度是指单位面积上林木的株数, 是评定立地生产力和影响林木生长发育的重要因子[14]。林分密度可以改变林内的光照、湿度、土壤养分等条件, 造成林下小环境以及灌草植物种类与数量分布的差异[15-16]。合理的林分密度能改善人工林的物种多样性, 优化林分结构, 有助于其向天然林群落方向演替[17]。本实验通过研究林分密度对柏树人工林林下物种多样性和土壤理化性质的影响, 筛选出适宜的林分密度, 以促进林分稳定性和自然更新能力, 为柏木人工林可持续发展与经营提供参考。
1 研究区域概况云顶山(30°71′—30°73′N, 104°47′—104°50′E)位于成都平原东北部金堂县境内龙泉山脉中段, 为金堂山主峰, 海拔982 m, 面积约67 km2, 南北走向, 南缓北陡, 属亚热带湿润季风气候, 四季分明。日照年平均1295.5 h, 年平均气温16.6℃。湿度大, 云雾多, 平均风速小, 无霜期长, 累计年平均285 d, 具有春来较早、夏长、秋冬短的特点。云顶山常年雨量充沛, 降雨时空不均, 平均年降水量920.5 mm。山上水源丰富, 溪流密布, 水动力保持较强。
由于温和的气候特征和丰沛的水资源, 云顶山植被丰富, 但因人为活动和乱砍滥伐的影响, 目前云顶山较完整的自然植被很少, 植被残次林较多, 主要植被类型有常绿阔叶林、常绿针叶林和山地灌丛。该地柏木人工林栽植于20世纪五六十年代, 初始造林密度较高, 大概为1100株/hm2左右, 随着时间的推移, 因自然稀疏、人为间伐和旅游开发干扰等因素, 密度逐渐产生差异。本次调查的柏木林龄基本一致, 平均为50年以上, 林分为公益林, 禁伐。
2 研究方法 2.1 样地设置及调查方法根据实地调查, 按照方精云[18]的研究方法, 采用典型样地法, 选取5种不同的现存林分密度A(500株/hm2)、B(650株/hm2)、C(800株/hm2)、D(950株/hm2)、E(1100株/hm2)柏木人工林, 每个密度各设置4个20 m×30 m的标准地, 共20个样地(表 1)。每个样地的坡向、坡位及海拔等条件基本一致, 对样地的乔木(DBH≥3 cm)进行每木检尺, 记录其胸径、树高和冠幅等数据, 在每块样地内沿对角线设置6个5 m×5 m的灌木(DBH < 3 cm)样方和12个1 m×1 m的草本样方, 分别记录样方内灌木和草本的种名、株数或丛数、高度、盖度等。
| 样地号 Sample plot No. |
海拔 Altitude/m |
坡向 Slope aspect/° |
坡位 Slope positions |
坡度 Slope position/° |
平均树高 Average height/m |
平均胸径 Average diameter/cm |
郁闭度 Canopy density |
密度 Density/(株/hm2) |
密度划分 Density class |
| 1 | 859 | NE50° | 上 | 20° | 14.09±0.72ab | 24.98±2.37a | 0.6 | 650 | B |
| 2 | 858 | NE60° | 上 | 19° | 14.47±0.43a | 19.51±0.80bc | 0.6 | 650 | B |
| 3 | 860 | NE54° | 上 | 21° | 12.33±0.72abcdef | 17.73±0.79bcd | 0.6 | 650 | B |
| 4 | 861 | NE48° | 上 | 26° | 10.09±0.64ef | 14.34±1.50d | 0.6 | 650 | B |
| 5 | 838 | NE66° | 中 | 21.5° | 11.67±0.22bcdef | 18.26±0.48bcd | 0.7 | 800 | C |
| 6 | 841 | NE65° | 中 | 20° | 11.58±0.36bcdef | 16.94±0.72bcd | 0.7 | 800 | C |
| 7 | 835 | 正东 | 中 | 23° | 13.58±1.45abc | 16.53±0.59bcd | 0.8 | 800 | C |
| 8 | 835 | 正东 | 中 | 25° | 11.05±0.44cdef | 15.45±0.59cd | 0.7 | 800 | C |
| 9 | 867 | 正南 | 中 | 28° | 12.56±0.21abcde | 16.70±0.59bcd | 0.5 | 500 | A |
| 10 | 834 | 正南 | 中 | 27° | 11.92±0.79abcdef | 17.02±1.45bcd | 0.5 | 500 | A |
| 11 | 829 | 正南 | 中 | 27° | 13.42±0.33abcd | 21.00±0.94ab | 0.5 | 500 | A |
| 12 | 827 | 正南 | 中 | 25° | 12.52±0.24abcde | 20.14±0.61bc | 0.5 | 500 | A |
| 13 | 830 | 正南 | 中 | 25° | 11.79±0.28abcdef | 17.35±0.28bcd | 0.9 | 1100 | E |
| 14 | 831 | 正南 | 中 | 26° | 12.60±0.39abcde | 18.06±0.81bcd | 0.9 | 1100 | E |
| 15 | 833 | 正南 | 中 | 25° | 13.27±0.29abcd | 20.50±0.36ab | 0.9 | 1100 | E |
| 16 | 830 | 正南 | 中 | 27° | 12.67±0.21abcde | 18.43±0.48bcd | 0.8 | 1100 | E |
| 17 | 835 | 正南 | 中 | 24° | 10.77±0.24def | 17.90±0.54bcd | 0.8 | 950 | D |
| 18 | 834 | 正南 | 中 | 22° | 9.57±0.43f | 17.14±0.76bcd | 0.8 | 950 | D |
| 19 | 834 | 正南 | 中 | 22° | 10.94±0.45cdef | 17.07±0.88bcd | 0.8 | 950 | D |
| 20 | 833 | 正南 | 中 | 21° | 10.14±0.39ef | 17.45±0.69bcd | 0.8 | 950 | D |
| NE:东北, Northeast;A:500株/hm2, 500 tree/hm2;B:650株/hm2, 650 tree/hm2;C:800株/hm2, 800 tree/hm2;D:950株/hm2, 950 tree/hm2;E:1100株/hm2, 1100 tree/hm2;不同小写字母表示在0.05水平上具有显著差异 | |||||||||
本实验采用α多样性指数综合评估群落物种多样性水平, 公式如下:
灌木层、草本层重要值:IV=(相对密度+相对盖度+相对频度) /3
丰富度指数(D):用相对物种丰富度来表示:D = S
Simpson指数(优势度指数): 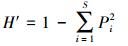
Shannon-Wiener指数: 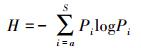
Pielou均匀度指数: 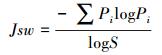
式中, Pi为第i种的个体数ni占调查物种个体总数n的比例, 即Pi=ni /n;i=1, 2, 3, …, S, S为物种数。
2.3 土壤取样及测定方法采用对角线五点法, 在调查区取0—20 cm混合土样带回实验室, 待土样风干后, 先剔除较大的植物残体和石碴, 再取100 g土样过2 mm孔径尼龙筛, 接着用四分法取出部分, 连续研磨直至使其全部通过0.25 mm孔径尼龙筛, 并将研磨过后的样品混匀、装袋保存、密封待测。
测定方法:烘干法测土壤含水量;电位法测土壤pH;高氯酸-硫酸硝化凯氏定氮法测土壤全氮含量;碱熔-铝锑抗比色法测全磷含量;盐酸-硫酸浸提法测速效磷含量;重铬酸钾-硫酸消化法测有机质含量;酸溶-火焰光度法测全钾含量;乙酸铵提取-火焰分光光度法测速效钾含量[19]。
2.4 数据处理采用SPSS 17.0软件中的单因素方差分析(One-Way ANOVA)和最小显著差异法(LSD)检验不同林分密度对柏树人工林林下植被物种多样性指数和土壤理化性质的差异显著性(P=0.05), 采用Excel 2010软件制作图表。
3 结果分析 3.1 不同林分密度下的物种组成在调查的20个样地中, 共记录到维管植物168种, 隶属75科140属, 其中以菊科(Compositae)、蔷薇科(Rosaceae)、百合科(Liliaceae)、禾本科(Gramineae)、忍冬科(Caprifoliaceae)等植物为主。如图 1所示, 随着林分密度由E到A变化, 灌木层物种数分别为26、28、45、45、43, 草本层物种数分别为33、43、40、51、33, 灌木层与草本层物种数均呈先增后减的趋势, 且在密度B出现最大值, 草本层物种数在密度D有所升高。高密度下草本层物种数较灌木层多, 随着乔木层郁闭度降低, 灌木层植物占据优势地位, 物种数逐渐多于草本层。

|
| 图 1 不同林分密度柏木人工林林下物种组成 Fig. 1 The understory species composition of Cupressus funebris plantanion with different densities A:500株/hm2;B:650株/hm2;C:800株/hm2;D:950株/hm2;E:1100株/hm2 |
不同林分密度对不同植物群落影响不同(图 2), 随着林分密度的变化, 灌木层Shannon-Wiener多样性指数H、Simpson优势度指数H′、Pielou均匀度指数Jsw均无显著差异(P>0.05), 基本在密度B为最大值, 而物种丰富度指数D存在显著差异(P<0.05), 并随林分密度降低, 呈先增后减的趋势, 在密度B出现峰值。草本层多样性指数变化较复杂, 差异性显著, 其物种丰富度指数D、Shannon-Wiener多样性指数H、Simpson优势度指数H′随林分密度降低均呈先增后减再增再减的双峰型变化趋势, 在密度D和B均出现峰值, 而Pielou均匀度指数Jsw则呈先增后减的趋势, 在密度D出现最大值。

|
| 图 2 不同林分密度下柏木人工林林下物种多样性指数 Fig. 2 Diversity index of understory vegetation in Cupressus funebris plantations with different densities |
对重要值的研究发现(表 2), 不同林分密度下灌木层与草本层优势种变化较小, 灌木层主要为石海椒(Reinwardtia indica)、黄荆(Vitex negundo)、荚蒾(Viburnum dilatatum)等植物, 草本层主要为莩草(Setaria chondrachne)、求米草(Oplismenus undulatifolius)、白接骨(Asystasiella neesiana)等植物, 优势种大多为阴性、耐荫性或适应性极强的植物, 说明上层乔木对林下植物抑制较强, 其群落优势种演替进行较为缓慢。在密度D和B, 灌木层中出现珊瑚朴(Celtis julianae)、蕊帽忍冬(Lonicera pileata)、构树(Broussonetia papyrifera)等阳生性树种且长势较好, 草本层中出现对光照要求较高的植物—麦冬(Ophiopogon japonicas), 说明这两种密度下林内光照资源较好, 阳生性植物逐渐生长并开始显现优势。除此之外, 在乔木层与灌木层的物种调查中均没有发现柏木幼苗, 说明当地柏木人工林乔木层几乎不存在自然更新。
| 层次 Layer |
密度 Density |
各层次优势种及重要值之和 The dominant species and sum of important value at all levels |
| 灌木层Shrub | E | 石海椒+黄荆+荚蒾+六道木+豆腐柴 Reinwardtia indica+Vitex negundo+Viburnum dilatatum+Abelia biflora+Premna microphylla(0.6614) |
| D | 黄荆+珊瑚朴+荚蒾+蕊帽忍冬+豆腐柴 Vitex negundo+Celtis julianae+Viburnum dilatatum+Lonicera pileata+ Premna microphylla(0.6395) |
|
| C | 黄荆+梧桐+石海椒+六道木+荚蒾 Vitex negundo+Firmiana platanifolia+Reinwardtia indica+Abelia biflora+Viburnum dilatatum(0.6731) |
|
| B | 构树+石海椒+珊瑚朴+荚蒾+黄荆 Broussonetia papyrifera+Reinwardtia indica+Celtis julianae+Viburnum dilatatum+Vitex negundo(0.6881) |
|
| A | 黄荆+石海椒+豆腐柴+荚蒾+六道木 Vitex negundo+Reinwardtia indica+Premna microphylla+Viburnum dilatatum+Abelia biflora(0.6241) |
|
| 草本层Herb | E | 莩草+沿阶草+求米草+白接骨+过路黄 Setaria chondrachne+Ophiopogon bodinieri+Oplismenus undulatifolius+Asystasiella neesiana+Lysimachia christinae(0.5548) |
| D | 莩草+白接骨+三花莸+求米草+麦冬 Setaria chondrachne+Asystasiella neesiana+Caryopteris terniflora+Oplismenus undulatifolius+Ophiopogon japonicas(0.4137) |
|
| C | 莩草+莎草蕨+求米草+过路黄+韩信草 Setaria chondrachne+Schizaea digitata+Oplismenus undulatifolius+Lysimachia christinae+Scutellaria indica(0.5995) |
|
| B | 莩草+过路黄+白接骨+麦冬+韩信草 Setaria chondrachne+Lysimachia christinae+Asystasiella neesiana+Ophiopogon japonicas+Scutellaria indica(0.5039) |
|
| A | 莩草+求米草+韩信草+过路黄+白接骨 Setaria chondrachne+Oplismenus undulatifolius+Scutellaria indica+Lysimachia christinae+Asystasiella neesiana(0.6962) |
如表 3所示, 林分密度对土壤pH、含水量、全磷、速效磷无显著影响(P>0.05), 对全氮、全钾、速效钾、有机质影响显著(P<0.05)。土壤pH值均大于7, 林地土壤为弱碱性, 与柏木生长特性相符。随着林分密度的降低, 土壤全氮、全钾、速效钾、有机质含量总体呈先增后减趋势, 在密度B出现峰值或较大值。
| 密度 Density |
pH | 含水量 Water content/% |
全氮 Total Nitrogen/(g/kg) |
全磷 Total phosphorus/(g/kg) |
速效磷 Available Phosphorus/(mg/kg) |
全钾 Total Potassium/(g/kg) |
速效钾 Available Potassium/(mg/kg) |
有机质 Organic Matter/(g/kg) |
| E | 7.70±0.04a | 0.16±0.01a | 1.33±0.03bc | 4.63±0.65a | 0.22±0.11a | 16.02±0.42ab | 90.18±7.20c | 46.46±11.48ab |
| D | 7.73±0.11a | 0.18±0.03a | 1.02±0.05c | 6.09±1.23a | 0.12±0.03a | 18.95±2.09ab | 133.85±16.86c | 33.18±0.57ab |
| C | 7.65±0.17a | 0.14±0.02a | 2.45±0.17a | 6.18±0.86a | 0.25±0.10a | 19.40±0.71a | 212.93±11.44ab | 58.91±3.65a |
| B | 7.73±0.09a | 0.19±0.01a | 2.51±0.04a | 7.44±0.41a | 0.34±0.05a | 18.74±0.97ab | 250.93±14.55a | 62.66±6.32a |
| A | 7.73±0.03a | 0.17±0.01a | 1.70±0.19b | 5.40±0.47a | 0.18±0.04a | 14.38±0.79b | 147.95±21.33bc | 28.21±6.76b |
土壤速效钾指标变异程度显著大于全钾指标, 全氮含量变化规律与有机质含量变化趋势基本一致。土壤理化性质与灌木层、草本层多样性指数的相关分析(表 4)表明:土壤pH值与灌木层和草本层物种多样性指数不存在明显的相关关系, 土壤含水量、全磷、全钾与草本层D呈显著或极显著正相关, 土壤速效磷与灌木层H呈显著负相关, 土壤全氮与灌木层D呈极显著正相关, 与草本层H、H′、Jsw呈显著或极显著负相关, 土壤速效钾与灌木层H、D和草本层D呈显著或极显著正相关, 与草本层Jsw呈显著负相关, 土壤有机质与灌木层H、Jsw呈显著正相关。
| 因子 Factor |
灌木层Shrub Layer | 草本层Herb Layer | |||||||
| H | H′ | D | Jsw | H | H′ | D | Jsw | ||
| pH | 0.18 | 0.2 | 0.23 | 0.14 | -0.09 | -0.1 | -0.11 | -0.03 | |
| SW | -0.13 | -0.14 | 0.16 | -0.19 | 0.32 | 0.18 | 0.48* | 0.12 | |
| TN | 0.38 | 0.1 | 0.71** | 0.11 | -0.45* | -0.50* | 0.32 | -0.67** | |
| TP | 0.04 | -0.09 | 0.35 | -0.13 | -0.02 | -0.14 | 0.49* | -0.3 | |
| AP | -0.28 | -0.55* | 0.19 | -0.43 | -0.07 | -0.03 | 0.23 | -0.18 | |
| TK | 0.23 | 0.19 | 0.01 | 0.27 | 0.4 | 0.25 | 0.57** | 0.13 | |
| AK | 0.53* | 0.34 | 0.72** | 0.29 | -0.19 | -0.28 | 0.55** | -0.52* | |
| OM | 0.50* | 0.36 | 0.28 | 0.44* | -0.08 | -0.05 | 0.19 | -0.16 | |
| H:Shannon多样性指数, Shannon diversity index;H′:Simpson优势度指数, Simpson dominance index;D:物种丰富度指数, Richness index;Jsw:Pielou均匀度指数, Pielou evenness index;pH:土壤pH值, Soil pH value;TN:全氮, Total nitrogen;TP:全磷, Total potassium;AP:有效磷, Available potassium;OM:土壤有机质, Soil organic matter;AK:速效钾, Available potassium;TK:全钾, Total potassium;SW:土壤含水量, Soil water content | |||||||||
物种多样性是反映群落物种组成、林分结构、动态演替等方面的重要指标[19]。林分密度通过控制乔木层截留的光照和水分影响林下植被的生长与分布[18]。雷相东等[20]认为郁闭度是影响天然林下层植物多样性的主要因子。在本实验中, 灌木层与草本层物种多样性对林分密度响应不同, 灌木层物种多样性指数随林分密度降低呈先增后减趋势, 在密度B出现峰值, 这是因为高密度的柏木人工林冠层结构对下层灌木有直接抑制作用[21], 随着林分密度降低, 林隙增多, 林下光照增强, 阴生性和中生性植物逐渐向阳生性植物方向发生演替, 故林下物种多样性增加, 同时植物光合作用增强, 林下可利用空间增大, 因此植物长势更优且分布均匀。草本层物种多样性指数呈双峰型变化趋势, 在密度D和B均出现峰值, 当林分密度为D时, 草本层物种数大幅上升, 灌草层中珊瑚朴、蕊帽忍冬、麦冬等阳生性植物生长较好并为优势种, 说明此密度下林内光照资源较好, 乔灌层对草本层的抑制减弱, 因而促进阳生性草本植物快速生长, 在密度B亦是因光照条件对下层植被生长有利, 物种丰富度达到最高, 由此说明光照是影响林下植被组成的重要因素, 这与Alem[22]等的研究结果相似。当林分密度降低到A时, 灌木层多样性指数依然保持较大值, 而草本层多样性指数下降, 这与鲁绍伟等[23]的低密度林分生物多样性优于高密度林分的研究结果不完全相同, 其原因可能是研究区域和调查所选林分密度的不同。在本实验中虽然密度降低在一定程度上使林下生境状况变好, 但灌木层较草本层生活力更强, 冠幅更大, 能吸收更多的光照, 因而加剧乔灌层以下的草本植物种内对生存空间与养分的竞争, 故其多样性指数降低且分布不均匀。因此, 林分过于稀疏时也不利于林下物种的更新和种群拓展[24]。
4.2 林分密度对柏木人工林土壤理化性质的影响林分密度影响林下生境的形成, 导致土壤空间异质性和理化性质差异[11]。在针叶树人工林中, 乔木层养分分解归还速率慢, 因而林下植被的作用更为重要[7]。在本实验中, 土壤理化性质随林分密度降低表现为土壤养分含量先增后减的变化趋势, 在密度B各养分含量均为较大值。柏木为针叶树种, 自然更新凋落速度较慢, 且高密度的柏木林对土壤养分吸收量大, 而光照和空间的不足导致林下植被稀疏, 凋落物较少且分解速度慢, 土壤腐殖质不足, 故有机质与其他养分含量偏低。由表 4可知, 土壤有机质与灌木层多样性指数呈显著正相关, 两者变化趋势基本一致, 说明灌木层植物为影响有机质的主要因子, 其枯枝落叶层相较于草本植物更为丰富[25]。土壤有机质是氮的重要来源[13], 有机质的积累有利于氮含量的增加, 故氮含量与灌木层D呈极显著正相关, 在本实验中全氮含量与有机质变化规律相似, 这与崔宁洁等[13]的研究结果一致。但全氮与草本层多样性指数呈显著负相关, 可能是因为草本层植物侧重对碳的吸收, 也可能是草本层多样性高, 因而从土壤中吸收的氮素较多, 造成土壤氮含量下降。速效钾随林分密度变化表现出显著差异, 且与灌木层和草本层D呈极显著正相关, 全钾与草本层D呈极显著正相关。植物群落与土壤为有机整体, 林下植被的丰富和稳定能够改善土壤结构, 减弱钾的淋溶作用, 故其含量增加, 而土壤钾肥的提高能够促进植物生长发育, 增加其物种多样性。同时发现, 速效钾与草本层Jsw呈显著负相关, 这可能与对钾吸收量较大的草本植物分布以及钾元素较强的移动性相关[26]。
4.3 该地柏木人工林可持续发展的建议与思考第八次全国森林资源清查结果显示, 我国人工林面积约占全国森林总面积的31.2%, 居世界首位[27]。与天然林相比, 人工林普遍存在结构简单、自然更新缓慢、生态效益低等特点[28]。云顶山风景区为旅游胜地, 其上有石城—宋元战争遗址和慈云寺—始建于南朝齐梁, 之后将建立的龙泉山城市森林公园“十园”, 云顶山森林公园便为其中之一, 云顶山很有可能会成为市民游览圣地, 这必然带来更大程度的人为干扰甚至导致生境破碎化, 因此除通过抚育间伐调整林分密度, 改善土壤质量外, 还应采取近自然林业的方法, 实施带状或块状改造[3], 补植生态与经济效益均较高的乡土树种, 营造异龄混交复层林, 从而提高森林的抗干扰和生态系统的自我修复能力, 以期发挥更大的生态价值和经济效益。
4.4 结论灌木层与草本层多样性指数均在密度B(650株/hm2)出现峰值, 表明此密度下光热、水分、大气等环境因子较有利于林下植物生长, 柏木人工林林下物种多样性和群落稳定性水平能够得到有效提升。对土壤相关数据分析可得, 全氮、钾、有机质为影响柏木人工林林下物种多样性的主要因子。本实验通过比较不同林分密度下柏木人工林林下物种多样性与土壤理化性质, 发现林分密度B能够为林下植物提供良好的生长空间并维持土壤养分特征, 因而可作为最适密度, 为该地柏木人工林的可持续经营提供参考。
| [1] |
陈丝露, 赵敏, 李贤伟, 范川, 肖宝茹. 柏木低效林不同改造模式优势草本植物多样性及其生态位. 生态学报, 2018, 38(1): 143-155. |
| [2] |
刘奕琳, 万福绪, 娄晓瑞. 盐胁迫对10个墨西哥柏种源幼苗生理生化的影响. 南京林业大学学报:自然科学版, 2013, 37(4): 29-33. |
| [3] |
龚固堂, 黎燕琼, 朱志芳, 陈俊华, 慕长龙, 吴雪仙, 郑绍伟. 川中丘陵区人工柏木防护林适宜林分结构及水文效应. 生态学报, 2012, 32(3): 923-930. |
| [4] |
李平, 李凤汀, 范川, 李贤伟, 张健, 黄明莉. 川中丘陵区柏木低效林改造模式植物多样性对土壤有机碳的影响. 生态学报, 2015, 35(8): 2667-2675. |
| [5] |
范川, 周义贵, 李贤伟, 张健, 廖洪流, 李凤汀, 冯茂松. 柏木低效林改造不同模式土壤抗蚀性对比. 林业科学, 2014, 50(6): 107-114. |
| [6] |
巢林, 洪滔, 李键, 陈灿, 洪伟, 吴承祯. 中亚热带不同林龄杉木人工林径级结构与林下物种多样性分析. 植物资源与环境学报, 2015, 24(2): 88-96. DOI:10.3969/j.issn.1674-7895.2015.02.13 |
| [7] |
李国雷, 刘勇, 于海群, 吕瑞恒, 李瑞生. 油松(Pinus tabulaeformis)人工林林下植被发育对油松生长节律的响应. 生态学报, 2009, 29(3): 1264-1275. DOI:10.3321/j.issn:1000-0933.2009.03.022 |
| [8] |
杨承栋, 焦如珍, 屠星南, 陈仲庐, 熊有强. 发育林下植被是恢复杉木人工林地力的重要途径. 林业科学, 1995, 31(3): 275-283. |
| [9] |
Carr C A, Krueger W C. Understory vegetation and ponderosa pine abundance in Eastern Oregon. Rangeland Ecology & Management, 2011, 64(5): 533-542. |
| [10] |
Rivaie A A. The effects of understory vegetation on P availability in Pinus radiata forest stands:a review. Journal of Forestry Research, 2014, 25(3): 489-500. DOI:10.1007/s11676-014-0488-4 |
| [11] |
盛炜彤. 不同密度杉木人工林林下植被发育与演替的定位研究. 林业科学研究, 2001, 14(5): 463-471. DOI:10.3321/j.issn:1001-1498.2001.05.001 |
| [12] |
李伟, 张翠萍, 魏润鹏. 广东中西部桉树人工林植物多样性与林龄和土壤因子的关系. 生态学报, 2014, 34(17): 4957-4965. |
| [13] |
崔宁洁, 张丹桔, 刘洋, 张健, 欧江, 张捷, 邓超, 纪托未. 不同林龄马尾松人工林林下植物多样性与土壤理化性质. 生态学杂志, 2014, 33(10): 2610-2617. |
| [14] |
罗素梅, 何东进, 谢益林, 李树忠, 董永平, 王其炳, 罗建, 林美浩, 张中瑞. 林分密度对尾赤桉人工林群落结构与生态效应的影响研究. 热带亚热带植物学报, 2010, 18(4): 357-363. |
| [15] |
何艺玲, 傅懋毅. 人工林林下植被的研究现状. 林业科学研究, 2002, 15(6): 727-733. DOI:10.3321/j.issn:1001-1498.2002.06.015 |
| [16] |
王树森, 余新晓, 罗于洋, 赵广亮, 杨君, 海龙. 人工造林对八达岭森林植被木本植物生物多样性的影响. 北京林业大学学报, 2008, 30(S2): 155-159. |
| [17] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [18] |
周树平, 梁坤南, 杜健, 李碧均, 周再知, 黄桂华. 不同密度柚木人工林林下植被及土壤理化性质的研究. 植物研究, 2017, 37(2): 200-210. |
| [19] |
王继丰, 韩大勇, 王建波, 付晓玲, 朱道光, 刘赢男, 曹宏杰, 黄庆阳, 谢立红, 钟海秀, 隋心, 倪红伟. 三江平原湿地小叶章群落沿土壤水分梯度物种组成及多样性变化. 生态学报, 2017, 37(10): 3515-3524. |
| [20] |
雷相东, 唐守正, 李冬兰, 陈宝升, 张则路. 影响天然林下层植物物种多样性的林分因子的研究. 生态学杂志, 2003, 22(3): 18-22. DOI:10.3321/j.issn:1000-4890.2003.03.004 |
| [21] |
郝建锋, 王德艺, 李艳, 姚小兰, 张逸博, 朱云航, 齐锦秋. 不同林分密度下川北白云山地区喜树人工林的群落结构和物种多样性. 植物研究, 2015, 35(5): 772-778. |
| [22] |
Alem S, Pavlis J, Urban J, Kucera J. Pure and mixed plantations of Eucalyptus camaldulensis and Cupressus lusitanica:their growth interactions and effect on diversity and density of undergrowth woody plants in relation to light. Open Journal of Forestry, 2015, 5(4): 375-386. DOI:10.4236/ojf.2015.54032 |
| [23] |
鲁绍伟, 刘凤芹, 余新晓, 王树森, 杨新兵, 李春平. 华北土石山区不同造林密度的油松林结构与功能研究. 干旱区资源与环境, 2007, 27(9): 144-149. DOI:10.3969/j.issn.1003-7578.2007.09.029 |
| [24] |
康冰, 刘世荣, 蔡道雄, 卢立华. 马尾松人工林林分密度对林下植被及土壤性质的影响. 应用生态学报, 2009, 20(10): 2323-2331. |
| [25] |
杨秉珣, 刘泉, 王彬. 嘉陵江流域不同类型植被多样性与土壤养分和酶活性的关系. 水土保持研究, 2016, 23(6): 45-51, 58-58. |
| [26] |
董艳红, 王火焰, 周健民, 任正文. 不同土壤钾素淋溶特性的初步研究. 土壤, 2014, 46(2): 225-231. |
| [27] |
国家林业局.第八次全国森林资源清查结果.北京: 国家林业局, 2014.
|
| [28] |
孙千惠, 吴霞, 王媚臻, 张柳桦, 姚小兰, 齐锦秋, 郝建锋. 林分密度对马尾松林林下物种多样性和土壤理化性质的影响. 应用生态学报, 2018, 29(3): 732-738. |