 2019, Vol. 39
2019, Vol. 39文章信息
- 苏宇, 何瑞, 尹海锋, 郭茂金, 李贤伟, 范川, 刘思泽.
- SU Yu, HE Rui, YIN Haifeng, GUO Maojin, LI Xianwei, FAN Chuan, LIU Size.
- 开窗补植银木对柏木低效林分生态化学计量特征的影响
- Effects of Cinnamomum septentrionale in hilly area of central sichuan basin on ecological stoichiometry characteristics of Cupressus funebris forest
- 生态学报. 2019, 39(2): 590-598
- Acta Ecologica Sinica. 2019, 39(2): 590-598
- http://dx.doi.org/10.5846/stxb201712212289
-
文章历史
- 收稿日期: 2017-12-21
- 网络出版日期: 2018-10-18






2. 剑阁县林业和园林局, 广元 628317
2. Administration of Forestry and Gardening of Jian'ge, Guangyuan 628317, China
森林生态系统中的基础元素(C、N、P)对植物的发育过程和生理功能都起着重要的作用[1-2]。植物体内C储量受N、P可获得量控制, C、N、P元素的耦合作用强于其他元素[3, 4]。现有理论认为植物体的生长速率以及植物体对生境中提供养分的利用效率可以通过对比C:N和C:P比值得知[5], 而N:P比值被更多地用来确定植物所处的种群或者群落受这两种化学元素限制的情况[6-9]。植物根系在土壤中吸收养分, 通过叶片的光合作用积累贮存C, 以凋落物的形式再将C、N等养分元素归还到土壤中, 这就形成了植物-凋落物-土壤之间的养分循环过程[10-11]。凋落物成为养分循环的桥梁, 不同植物类型的凋落物养分归还过程的异质性使得森林生态系统中的C、N、P化学计量特征关系复杂, 已成为研究的重点[12-13]。
在川中丘陵区的低效柏木(Cupressus funebris)纯林中, 群落结构单一, 林分稳定性降低及地力衰退等问题亟待解决。在柏木纯林中“开窗补阔”团块状配置乡土阔叶树种, 构建复合森林生态系统——针阔混交林, 可以有效的促进生态系统循环, 恢复退化林分[14]。林窗内次生植被生物种类和养分分配方式的转变, 随着演替的进行使得林窗形成前后的C、N、P元素生态化学计量特征产生差异[15], 充分认识养分比例在植物-凋落物-土壤系统中的分配的调节机制, 以及系统中凋落物的养分归还比率对土壤养分循环的影响过程十分重要[16]。所以研究植物生长过程中植物体-凋落物-土壤三者之间的关系对提高该区低效林的经营管理水平具有重要现实意义。本研究集中揭示林窗补植的银木(Cinnamomum septentrionale)改造过程中对植物、土壤、凋落物C:N:P比值的影响及三者之间的相互关系;探讨作为乡土阔叶树种的银木是否从群落上改变了区域环境的元素平衡关系。为川中丘陵区大面积纯林改造丰富实践经验, 实现低效林改造目的, 充分发挥其生态功能, 为进一步建设长江上游优良生态屏障提供参考。
1 材料与方法 1.1 研究区概况研究区位于四川省德阳市旌阳区和新镇永新村(31°5′—31°20′N, 104°15′—104°33′E), 靠近川中丘陵北部边缘。该区柏木人工纯林为20世纪80年代中期营建的长江中上游防护林, 期间基本未采取任何林业经营措施和抚育管理, 林分密度2300—2570株/hm2, 2015年测得平均树高6.5 m, 胸径12 cm, 枝下高3 m, 林分郁闭度多在0.7—0.8, 甚至有达0.9。该区域地貌以低山丘陵为主;土壤类型为碱性紫色土;气候属于亚热带季风性湿润气候, 年均温16.0℃;年平均降雨量905.9 mm。
1.2 样地设置与样品采集方法 1.2.1 样地设置2012年3月上旬, 根据前期样地调查, 在试验区内选择坡位(中坡位)、坡度(25—30°)、坡向(西南坡)等立地条件相同、林地质量相近、造林密度相同、生长情况大体一致的柏木林, 进行人工开窗并补植银木。本研究中人工采伐原有柏木林形成200 m2林窗, 共3个, 林窗形状均为南北向长、东西向短的近似椭圆形, 各林窗边界间距不小于10 m。采伐时, 将砍伐的柏木树干、树枝等残体移出, 原林下灌草层植物连根清除, 并进行柏木根掘。在林窗中以“品”字形补植银木, 银木苗株高40—50 cm, 采用规格50 cm×50 cm×50 cm的窝穴栽种, 株行距为1 m×1.5 m。以未采伐开窗的柏木人工林为对照, 设置10 m×20 m的200 m2样方, 重复3次, 一共6个试验区域, 样地基本特征见表 1。
| 处理 Treatment |
乔木平均高 Arbor height/m |
乔木平均胸径 Arbor BDH/cm |
主要植物 Main plants |
植物物种数 Number of species |
枯落物层平均厚度 Litter layer thickness /cm |
| 柏木人工林 Pure cypress forest |
6.5 | 12 | 柏木(Cupressus funebris), 女贞(Ligustrum lucidum), 油桐(Vernicia fordii), 构树(Moraceae broussonetia), 桤木(Alnus cremastogyne), 麻栎(Quercus acutissima) | 12 | 0.4 |
| 银木林窗 Gap with Cinnamomum septentrionale |
4.5 | 7 | 银木(Cinnamomum septentrionale) | 35 | 3.2 |
2015年9月, 待采伐3年后, 在每个银木补植林窗区域中随机各选择生长状况大体一致, 生长位置相近的3株银木做标记, 并放置凋落物网, 共设置9个凋落物收集网袋;柏木林地, 在样地中心随机选择生长状况大体一致, 生长位置相近的3株柏木, 放置个凋落物网, 共设置9个凋落物收集网袋。总共放置18个凋落物网, 凋落物网规格为1 m×1 m, 100目尼龙网, 距地面约0.5 m。
2015年10月(10月)、2015年12月(12月)、2016年4月(4月)及2016年7月(7月), 按照品字布点, 在每块样地中随机选择3株林木, 于林窗内采集银木新鲜叶片, 在未改造柏木纯林样地中心采集柏木新鲜叶片。将叶片带回实验室, 放入烘箱内在105℃杀青15 min, 再在65℃下烘干48 h, 随后将烘干的树叶用粉碎机磨碎, 过2 mm筛子后, 用塑封袋保存起来用于C、N、P含量的测定分析。
同期, 对本实验设置的18个凋落物网进行凋落物收集, 每个样品采用四分法取样500 g入袋。带回实验室后, 去除凋落物中的杂质, 装入尼龙袋中, 再在65℃下烘干48 h, 随后将烘干的树叶片用粉碎机磨碎, 过2 mm筛子后, 用塑封袋保存起来用于C、N、P含量的测定分析。
在银木林窗内与未改造柏木样方下采集0—5 cm(Ⅰ)、5—20 cm(Ⅱ)、20—40 cm(Ⅲ)土层的土壤, 每个样品采用直径为6 cm的土芯取样器采集土壤约0.5 kg土样样品装入封口袋中, 带回实验室。让土壤经自然风干, 挑去活体根系、石块, 磨碎后分别过2 mm土筛、0.15 mm土筛, 然后装入封口袋中, 用于土壤C、N、P含量的分析。
1.3 样品处理及分析测试 1.3.1 样品测试植物样品的C、N、P含的测定分别采用重铬酸钾氧化-外加热法测定(GB 9834-88), 全自动凯氏定N仪进行测定, 硝酸-高氯酸-钼锑抗比色法(6505紫外分光光度计UK)测定(GB 7887-87)。
土壤样品的C、N、P含的测定分别采用重铬酸钾氧化-外加热法测定(LY/T 1237—1999), 全自动凯氏定N仪进行测定, 硝酸-高氯酸-钼锑抗比色法(6505紫外分光光度计UK)测定(LY/T 1232—1999)。
1.3.2 N、P养分回流率凋落物养分回收能力可以用“养分回流率(R)”[3]表示, 可以定义为当秋季来临时, 植物体正常生长器官的养分含量与地面凋落物中养分含量的差值, 即凋落物在其凋落前转移出的养分的量。计算公式如下:
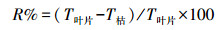
|
式中, R为养分回流率;T叶片为正常生长叶片中的养分含量(g/kg);T枯为凋落物中养分含量(g/kg)。
1.4 数据处理数据采用EXCEL 2010和SPSS 20.0软件进行数据统计, 数据分析显著性水平采用P<0.05。采用单因素方差分析(one-way ANOVA)和Tukey HSDa检验柏木低效林不同季节不同树种植物叶片、凋落物和土壤C、N、P化学元素及化学计量比的显著性差异, 并采用Origin 8.5软件绘制C、N、P含量的季节动态图。采样Pearson相关性分析检验土壤基本性质和土壤C、N、P化学元素及其化学计量比的相关性。
2 结果分析 2.1 柏木纯林与银木林分的化学计量特征 2.1.1 柏木与银木叶片元素含量季节变化特征柏木和银木两种树种的C含量随季节变化的趋势不同, 而在同一树种上叶片与凋落物养分含量在季节变化上便呈现一直的变化规律。柏木的叶片和凋落物的C含量均显著大于银木(P<0.01), 在4月呈现峰值, 在7月呈现最低值。同时, 季节对于柏木的叶片C的含量影响显著, 而凋落物C则只在4月和7月表现出显著的差异(P<0.01)。同树种同时期的凋落物C含量低于鲜叶的C含量, 银木表现显著。银木鲜叶的C在年际中并没有显著的变化, 但凋落物C在12月份最低, 并在次年的7月呈现显著的增加(P<0.01), 见图 1。

|
| 图 1 柏木与银木叶片和凋落物的C、N、P含量季节变化 Fig. 1 Nutrient concentration of leaves and litter of Cupressus funebris and Cinnamomum septentrionale |
如图 1, 柏木和银木的N含量随着季节的变化皆呈现显著性的差异, 但年际变化趋势表现一致。两种树种都表现为凋落物N含量显著低于鲜叶, 其中12月份含量最低, 次年的4月呈现峰值。P含量与N含量表现出一致的变化规律, 皆在12月份含量最低, 在次年的4月含量表现出年际的峰值(图 1)。且凋落物的P含量低于鲜叶, 柏木的P含量高于银木(P<0.01)。
2.1.2 柏木与银木凋落物养分回流率如图 2所示, 柏木纯林与林窗补植银木的凋落物在养分回流率存在一定的差异。C、N的回流率银木>柏木。P的回流率在不同的采样季节表现出不同幅度的波动, 柏木凋落物P的季节波动强于银木, 年际回流率在10月、12月高于银木, 而在次年的4月显著的低于银木。在N的养分回流规律上, 柏木与银木表现相对一致, 在12月达到峰值, 在次年的4月最低。年平均回流率的表现为银木凋落物C回流率为5.85%, 柏木凋落物C的回流率为2.72%。银木凋落物N的回流率为28.60%, 柏木凋落物N的回流率为18.35%。银木凋落物P的回流率为51.69, 柏木凋落物P的回流率为50.00%。

|
| 图 2 凋落物养分回流率 Fig. 2 Litter nutrient return rate |
柏木与银木叶片鲜叶、凋落物和土壤的C、N、P的化学计量特征可以看出, 柏木的凋落物的C:N, C/P, N:P均显著的大于叶片, 在未改造的柏木人工林中不同土层的C:N随着土层的加深而在20—40 cm土层呈现最大值(19.23±1.46), 同时变异系数(CV)也随之增加。林窗补植银木在C:N计量特征上的表现与柏木呈现一致性的规律, 但银木叶片与凋落物的C:N都显著的低于柏木, 凋落物C:N的变异系数大于叶片, 见表 2。
| 处理 Treatment |
层次 Layer |
碳氮比 C:N |
变异系数 C.V/% |
碳磷比 C/P |
变异系数 C.V/% |
氮磷比 N:P |
变异系数 C.V/% |
| 柏木人工林 | 植物叶片 | 99.14±2.91b | 2.94 | 1067.74±83.67b | 7.84 | 10.77±0.76b | 7.02 |
| Pure cypress forest | 植物凋落物 | 118.28±5.88a | 4.97 | 2106.03±22.23a | 1.06 | 17.83±0.77a | 4.3 |
| 土壤(0—5 cm) | 9.93±0.18D | 1.85 | 85.77±21.77A | 25.38 | 8.64±2.22A | 25.66 | |
| 土壤(5—20 cm) | 12.55±0.24B | 1.89 | 71.91±7.90A | 10.99 | 5.73±0.53A | 9.29 | |
| 土壤(20—40 cm) | 19.23±1.46A | 7.6 | 87.36±14.13A | 16.17 | 4.59±0.99A | 21.49 | |
| 土壤(0—40 cm) | 12.30±0.22C | 1.78 | 79.36±2.46A | 3.1 | 6.45±0.17A | 2.67 | |
| 银木林窗 | 植物叶片 | 52.59±2.07b | 3.93 | 559.73±20.06b | 3.58 | 10.64±0.17b | 1.61 |
| Gap with Cinnamomum | 植物凋落物 | 69.44±3.04a | 4.37 | 1089.55±28.38a | 2.6 | 15.72±1.06a | 6.73 |
| septentrionale | 土壤(0—5 cm) | 18.44±0.58B | 3.17 | 65.70±3.82A | 5.82 | 3.56±0.18AA | 5.18 |
| 土壤(5—20 cm) | 27.82±1.22A | 4.39 | 58.70±0.63B | 1.07 | 2.11±0.10C | 4.61 | |
| 土壤(20—40 cm) | 34.31±2.19A | 6.38 | 43.81±2.59B | 5.91 | 1.28±0.10D | 7.68 | |
| 土壤(0—40 cm) | 23.51±0.25A | 1.08 | 57.25±1.95B | 3.41 | 2.43±0.06B | 2.37 | |
| 不同小写字母表示同一植物叶片与凋落物化学计量比之间差异显著;不同大写字母表示同一林地不同土壤层次的化学计量比差异显著 | |||||||
两种树种的凋落物C/P是叶片的2倍, 同时在叶片呈现出高变异系数。在两种树种的对应土壤C/P表现出了异质性, 在柏木人工林分中5—20 cm土层显著的低于表层土和20—40 cm土层, 但银木林窗的土壤则打破了该规律, 呈现出随着土层的加深C/P降低的变化趋势。
柏木与银木叶片的N:P显著的低于凋落物的含量, 对应土壤中的N:P则都呈现出随着土层深度增加而减小的变化趋势。
2.3 植物-凋落物-土壤化学计量特征的相关性柏木人工林中叶片和凋落物的N, P含量呈现正相关的线性关系(R2=0.7961, R2=0.8677)。但在土壤中的C, P含量呈现负相关的线性关系。在银木林窗的土壤中C, P则正与柏木人工林土壤相反, 呈现正相关的相关关系。柏木人工林土壤的N与C:N并没有呈现线性的相关关系(R2=0.0299), 而除此以外柏木与银木在叶片, 凋落物和土壤的N与C:N, P含量与C:P的线性关系上都呈现正相关关系, 见表 3。
| 处理 Treatment |
y为P含量 x为N含量 |
y为C:N x为N含量 |
y为C:P x为P含量 |
|
| 柏木人工林 | 植物叶片 | y=0.1186x - 0.1542 | y =-17.658x+209.23 | y=-2018.7x+2272.7 |
| Pure cypress forest | R2=0.7961 | R2=0.9122 | R2=0.9383 | |
| 植物凋落物 | y=0.0855x - 0.1451 | y=-27.973x + 270.63 | y=-10235x + 5728.2 | |
| R2=0.8677 | R2=0.9228 | R2=0.8388 | ||
| 土壤(0—40 cm) | y=-0.2209x + 0.6293 | y=-2.044x + 15.721 | y=-501.35x + 209.47 | |
| R2=0.9855 | R2=0.0299 | R2=0.7066 | ||
| 银木林窗 | 植物叶片 | y=0.1025x - 0.0817 | y=-5.2013x + 105.43 | y=-600.94x + 1144.7 |
| Gap with Cinnamomum | R2=0.9097 | R2=0.9856 | R2=0.9669 | |
| septentrionale | 植物凋落物 | y=0.0818x - 0.1228 | y=-10.298x + 144.9 | y=-10.89x + 21.073 |
| R2=0.9488 | R2=0.9667 | R2=0.6561 | ||
| 土壤(0—40 cm) | y=1.6276x - 1.559 | y=-21.397x + 50.929 | y=-118.12x + 119.45 | |
| R2=0.9585 | R2=0.6811 | R2=0.9768 |
针对柏木和银木叶片与之所对应土壤C、N、P含量相关分析表明, 在柏木人工林中, 土壤P含量与叶片与凋落物中的C含量都表现为极显著的正相关关系, 而与叶片N含量呈正相关, 与凋落物呈现负相关。在林窗处理的土壤氮含量与银木叶片、凋落物的C、N、P含量含量都显著正相关。土壤P含量与叶片N含量、凋落物N含量呈现极显著的负相关关系。在进行林窗混交银木的改造之后, 土壤的C含量与银木叶片的C含量呈现极显著的正相关关系, 并进一步强化了叶片中的N、P, 凋落物的C、N、P与土壤C之间的相互关系, 见表 4。
| 处理 Treatment |
指标 Index |
叶片碳 Leaf C |
叶片氮 Leaf N |
叶片磷 Leaf P |
凋落物碳 Litter C |
凋落物氮 Litter N |
凋落物磷 Litter P |
| 柏木人工林 | 土壤C | 0.471 | 0.751 | 0.900** | 0.719 | -0.727 | 0.327 |
| Pure cypress forest | 土壤N | -0.485 | -0.762 | -0.893** | -0.730 | 0.738 | -0.312 |
| 土壤P | 0.997** | 0.906** | -0.038 | 0.925** | -0.920** | -0.735 | |
| 银木林窗 | 土壤C | 0.853** | 0.889** | 0.873** | 0.821** | 0.864** | 0.880** |
| Gap with Cinnamomum | 土壤N | -0.462 | -0.583 | -0.521 | -0.449 | -0.537 | -0.518 |
| septentrionale | 土壤P | -0.739 | -0.810** | -0.718 | -0.720 | -0.764** | -0.777** |
| 柏木人工林的土壤指标与柏木叶片的C、N、P含量进行相关性分析, 银木林窗的土壤指标与银木叶片的CNP含量进行相关性分析 | |||||||
当对柏木人工林中植物、凋落物、土壤的C:N、C:P、N:P进行对数转换之后进行Pearson进行相关性分析, 土壤的N:P与凋落物的C:N呈现显著的正相关(P<0.05), 而与凋落物的N:P呈现显著的负相关关系(P<0.05)。但在柏木叶片的化学计量特征则表现不同, 仅在叶片C:N与C:P表现出显著的正相关关系(P<0.05), 见表 5。
| 土壤碳氮比 Soil C:N |
土壤 碳磷比 Soil C:P |
土壤 氮磷比 Soil N:P |
叶片 碳氮比 Leaf C:N |
叶片 碳磷比 Leaf C:P |
叶片 氮磷比 Leaf N:P |
凋落物 碳氮比 Litter C:N |
凋落物 碳磷比 Litter C:P |
|
| 叶片碳氮比Leaf C:N | -0.713 | -0.067 | 0.191 | |||||
| 叶片碳磷比Leaf C:P | -0.264 | 0.000 | 0.513 | 0.824* | ||||
| 叶片氮磷比Leaf N:P | 0.730 | -0.306 | -0.271 | -0.652 | -0.474 | |||
| 凋落物碳氮比Litter C:N | 0.256 | 0.613 | 0.825* | -0.185 | 0.173 | 0.273 | ||
| 凋落物碳磷比Litter C:P | 0.323 | 0.474 | 0.349 | -0.547 | -0.286 | -0.194 | 0.247 | |
| 凋落物氮磷比Litter N:P | -0.218 | -0.291 | -0.822* | -0.309 | -0.765 | 0.071 | -0.700 | -0.086 |
| 对植物-凋落物-土壤计量比进行了对数转换。*.在0.05水平(双侧)上显著相关 | ||||||||
本研究中, 柏木和银木叶片的年均C含量高于全球陆生植物叶片的C含量(464 g/kg), 其中柏木林C含量(603.56 g/kg)高于珠江三角洲以马尾松为主要建群种的针叶林(517.85), 银木(499.92g/kg)高于米老排(Mytilaria laosensis)等常绿阔叶林[1, 17]。而N、P含量显著的低于全国平均水平(20.2 g/kg和1.46 g/kg)[18], 植物叶片的C:N和C:P在一定程度上表征这单位养分供应量所能产生的生产力或者表示植物对养分的利用效率[19]。这种高C低N、P的养分储存形式, 使得研究区林木生长呈现高C:N与C/P特征, 也就意味着植物呈现高C同化能力与高养分利用能力[20]。由于树种间不同的生长发育需求, 元素需求比例也不同, 柏木叶片的C:N与C/P都显著的高于银木叶片, 这一规律普遍表现在全球范围内的常绿针叶树种与常绿阔叶树种的比较中[9]。常绿的针叶树种叶片中富碳次生物多于阔叶树种, 而常绿阔叶树种养分的分配往往会集中于非溶性蛋白以增强包围结构, 从而提高叶寿命[21]。叶片营养元素的含量与自身组织结构特点和生长节律关系显著[22]。在本研究中, 柏木C:N在12月份为132.79, 而在次年的4月下降到了84.91, 这种大幅度的下降正是由于4月份的高生长发育期与12月的低生长休眠期对比形成的(图 1)。与柏木叶片相比, 银木叶片的低C:N, 和C:P都体现出银木在幼龄阶段快速生长的特点。旺盛的代谢活动, 伴随酶合成大量的蛋白, 这个过程需要大量的N参与, 而同时核糖体中P的需求能更好的完成这一过程。生长代谢过程可以通过叶片C、N、P化学计量特征来解释[23]。
植物的生长限制条件也可以通过化学计量比值来确定, 生态化学计量学理论认为正常的生物体的养分组成基本稳定, 若超出该范围则说明受到某一元素的限制[17, 24]。通常植物叶片的N:P比值来反映植物受N或P的限制作用, 当植物叶片N:P<14时, 植物生长主要受N限制;当N:P>16时, 植物生长主要受P限制;当14<N:P<16时, 植物生长受N和P共同限制[25]。但柏木与银木叶片N:P分别为10.77、10.64都明显处于N限制的阈值以下, 本研究区的植物生长主要受到了N元素的限制, N是该地区树木生长的主要限制因子[26]。生长速率假说认为个体生长速率与N:P呈现负相关的相关关系[27], 但在本研究中柏木与银木的显著生长速率差异性并没有在该化学计量比中得到体现, 这并不能用假说来解释, 这与不同林龄马尾松的研究一致[28]。
3.2 柏木人工林与银木林窗凋落物与土壤C、N、P含量及化学计量特征凋落物的养分来源于叶片, 植被类型的改变可以显著的改变凋落物的化学计量特征。植物叶片在凋落前将营养元素转移到了其他的组织结构, 这种养分的重吸收, 再分配能力可以有效的提高养分的利用效率, 增长营养元素在植物体内存留的时间[29]。不同的植物叶片对于养分的重吸收作用并不一致, 当对柏木纯林进行林窗式的补植银木后, 两种林分的凋落物养分回流率表现出了显著的差异(图 2), 银木N、P元素回流率都显著高于柏木。但本研究中N、P再吸收率均明显低于全球N、P再吸收率(57.4%, 60.7%), 表明土壤养分供应相对充足的情况下, 植物可能主要以提高养分吸收能力而非养分再吸收能力的方式适应环境[30-31]。柏木与银木凋落物较叶片在C:N、C:P和N:P特征上都显著的增加(表 2), 银木叶片与凋落物C:N较柏木较低, 一般认为这种比例下的植物残体更易分解, 分解速率更快, 同时凋落物N含量的增加或C:N比的降低都会增加凋落物的分解速率[32-33]。N:P的增加表明了植物对于N的再吸收率大于P, 且分解过程收P的限制越强[22]。银木的植入可以有效的降低凋落物的N:P, 这说明阔叶树种的引入有利于提高分解的相对速率, 有利于养分的循环。
柏木人工林土壤C:N为12.3, 稍高于我国土壤(10—12)的平均值, 当进行银木植入后的土壤C:N显著增高达到23.5, 已达到全球平均水平[34-35]。土壤的生态化学计量特征随着地上植物和凋落物的养分变化而改变, 但这种关系十分复杂, 并非单一、线性的相关关系(表 2)。土壤的化学计量指标是反应土壤资源有效性的重要指标[36], 土壤C:N比值与有机质分解速率呈反比, 而土壤C:P比值能够指示P有效性的高低。在柏木人工林分中土壤C:N比随着土层的深度而增加, 而土壤的C/P比并没有在土层的深度变化下表现规律性的变化, 在不同土层P有效性都表现的较为稳定。C:N在柏木土壤中低于补植银木土壤, 而土壤C/P银木混交林分则显著的低于柏木, 这印证了银木的较柏木优化的养分分解速率, 且土层内变异系数的增加也说明土壤异化过程的进展。
3.3 柏木人工林与银木林窗植物-凋落物-土壤C、N、P化学计量比的相关性分析在土壤的养分变化过程中影响因素众多, 基于C、N、P生态化学计量特征的研究, 有助于了解基础元素在森林生态系统土壤-凋落物-植被界面的循环过程。本研究中两种树种的C、N含量表现为叶片>凋落物>土壤, P含量大体表现为叶片>土壤>凋落物, 这说明叶片中养分储存量是最高的, 凋落物和土壤次之, 这一规律性的分布在树种间表现出一致性。叶的C:N:P的变化能力影响土壤养分关系, 反过来土壤对植物养分有效性也会产生反馈[37]。凋落物中C、N、P含量明显低于叶片, 这说明叶在衰落前后养分的再吸收状况, 其实也是植物在养分供应有限的环境中进化出来的一种机制[38]。
柏木与银木叶片N含量与C:N以及P含量与C:P都表现为负相关关系, 植物体对N、P元素的利用效率会随着N、P含量的升高而不断降低, 这与Hedin[39]的研究结果一致。同时本研究区柏木与银木叶片与凋落物的N、P含量表现出显著的线性相关关系, 这与喀斯特森林群落植物表现一致[40]。地上植被的改变可以有效的触发森林土壤养分循环过程, 当对柏木纯林进行林窗式阔叶树种的引入后, 土壤营养元素与对应植物叶片与凋落物间的相关性增加(表 4)。研究区域林分叶片-凋落物-土壤的C:N、C:P、N:P间的相关性并不显著, 但在对计量比进行了对数转换后, 可以发现柏木人工林土壤N:P与凋落物C:N、N:P上的相关关系, 这说明了生态系统内部C、N、P循环是在植物、凋落物和土壤3个库之间运输和转换的。
4 结论(1) 银木的生长速率高于柏木, 但生长速率假说并不能有效说明柏木与银木生产速率之间的差异。
(2) 柏木与银木叶片N:P分别为10.77, 10.64, 都明显处于N限制的阈值以下, 生长过程主要受到了N元素的限制, N即为川中丘陵区树木生长的主要限制因子。
(3) 林窗中银木凋落物N含量高于柏木, C:N、N:P小于柏木, 土壤与“植物-凋落物”间的相关性增加。因此, 阔叶树种的引入有利于提高分解的相对速率, 触发森林土壤养分循环过程。
| [1] | 吴统贵, 陈步峰, 肖以华, 潘勇军, 陈勇, 萧江华. 珠江三角洲3种典型森林类型乔木叶片生态化学计量学. 植物生态学报, 2010, 34(1): 58–63. DOI:10.3773/j.issn.1005-264x.2010.01.009 |
| [2] | Niklas K J, Owens T, Reich P B, Cobb E D. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth. Ecology Letters, 2005, 8(6): 636–642. DOI:10.1111/ele.2008.8.issue-6 |
| [3] | 郭子武, 陈双林, 杨清平, 李迎春. 密度对四季竹叶片C、N、P化学计量和养分重吸收特征的影响. 应用生态学报, 2013, 24(4): 893–899. |
| [4] | Knecht M F, Göransson A. Terrestrial plants require nutrients in similar proportions. Tree Physiology, 2004, 24(4): 447–460. DOI:10.1093/treephys/24.4.447 |
| [5] | Ågren G I. The C:N:P stoichiometry of autotrophs-theory and observations. Ecology Letters, 2004, 7(3): 185–191. |
| [6] | Aerts R, Chapin Ⅲ F S. The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30: 1–67. DOI:10.1016/S0065-2504(08)60016-1 |
| [7] | Güsewell S, Jewell P L, Edwards P J. Effects of heterogeneous habitat use by cattle on nutrient availability and litter decomposition in soils of an Alpine pasture. Plant and Soil, 2005, 268(1): 135–149. DOI:10.1007/s11104-004-0304-6 |
| [8] | Güsewell S, Koerselman W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology, Evolution and Systematics, 2002, 5(1): 37–61. DOI:10.1078/1433-8319-0000022 |
| [9] | Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow TA, Cotner J B, Harrison F, Hobbie S E, Odell G M, Weider LW. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540–550. DOI:10.1046/j.1461-0248.2000.00185.x |
| [10] | 皮发剑, 舒利贤, 喻理飞, 严令斌, 周晨, 吴正花, 袁丛军. 黔中喀斯特10种优势树种根茎叶化学计量特征及其关联性. 生态环境学报, 2017, 26(4): 628–634. |
| [11] | 卢同平, 史正涛, 牛洁, 张文翔. 我国陆生生态化学计量学应用研究进展与展望. 土壤, 2016, 48(1): 29–35. |
| [12] | Michaels A F. The ratios of life. Science, 2003, 300(5621): 906–907. DOI:10.1126/science.1083140 |
| [13] | 张蕾蕾, 钟全林, 程栋梁, 费玲, 李静, 吴永宏, 张中瑞, 王卫军. 刨花楠不同相对生长速率下林木叶片碳氮磷的适应特征. 生态学报, 2016, 36(9): 2607–2613. |
| [14] | 王谢.柏木人工林土壤有机碳组分对人工更新林窗的早期响应机制[D].成都: 四川农业大学, 2015. |
| [15] | 罗艳, 贡璐, 朱美玲, 安申群. 塔里木河上游荒漠区4种灌木植物叶片与土壤生态化学计量特征. 生态学报, 2017(24): 8326–8335. |
| [16] | 曾凡鹏, 迟光宇, 陈欣, 史奕. 辽东山区不同林龄落叶松人工林土壤-根系C:N:P生态化学计量特征. 生态学杂志, 2016, 35(7): 1819–1825. |
| [17] | O'Brien E, Dowling D. The evolution of ecosystem processes:growth rate and elemental stoichiometry of a key herbivore in temperate and arctic habitats. Journal of Evolutionary Biology, 2000, 13(5): 845–853. DOI:10.1046/j.1420-9101.2000.00215.x |
| [18] | Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377–385. DOI:10.1111/j.1469-8137.2005.01530.x |
| [19] | Ågren G I. Stoichiometry and nutrition of plant growth in natural communities. Annual Review of Ecology, Evolution, and Systematics, 2008: 39153–170. |
| [20] | 黄建军, 王希华. 浙江天童32种常阔叶树叶片的营养及结构特征. 华东师范大学学报:自然科学版, 2003(1): 92–97. |
| [21] | 郑淑霞, 上官周平. 近一世纪黄土高原区植物气孔密度变化规律. 生态学报, 2004, 24(11): 2457–2464. DOI:10.3321/j.issn:1000-0933.2004.11.016 |
| [22] | 曾昭霞, 王克林, 刘孝利, 曾馥平, 宋同清, 彭晚霞, 张浩, 杜虎. 桂西北喀斯特区原生林与次生林鲜叶和凋落叶化学计量特征. 生态学报, 2016, 36(7): 1907–1914. |
| [23] | Aerts R. Nutrient resorption from senescing leaves of perennials:are there general patterns?. Journal of Ecology, 1996, 84(4): 597–608. DOI:10.2307/2261481 |
| [24] | He J S, Wang L, Flynn D F B, Wang X P, Ma W H, Fang J Y. Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes. Oecologia, 2008, 155(2): 301–310. DOI:10.1007/s00442-007-0912-y |
| [25] | Koerselman W, Meuleman A F M. The vegetation N:P ratio:a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441–1450. DOI:10.2307/2404783 |
| [26] | 李靖.不同林龄杉木和华北落叶松人工林氮磷养分特征与根构型研究[D].西安: 西北农林科技大学, 2013. |
| [27] | Elser J J, Dobberfuhl D R, Mackay N A, Schampel J H. Organism size, life history, and N:P stoichiometry:toward a unified view of cellular and ecosystem processes. BioScience, 1996, 46(9): 674–684. DOI:10.2307/1312897 |
| [28] | 崔宁洁, 刘小兵, 张丹桔, 张健, 刘洋, 邓长春, 纪托未, 陈亚梅. 不同林龄马尾松(Pinus massoniana)人工林碳氮磷分配格局及化学计量特征. 生态环境学报, 2014, 23(2): 188–195. DOI:10.3969/j.issn.1674-5906.2014.02.002 |
| [29] | Wardle D A, Walker L R, Bardgett R D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science, 2004, 305(5683): 509–513. DOI:10.1126/science.1098778 |
| [30] | Han W X, Tang L Y, Chen Y H, Fang J Y. Relationship between the relative limitation and resorption efficiency of Nitrogen vs Phosphorus in woody plants. PLoS One, 2013, 8(12): e83366. DOI:10.1371/journal.pone.0083366 |
| [31] | 安卓, 牛得草, 文海燕, 杨益, 张洪荣, 傅华. 氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C:N:P化学计量特征的影响. 植物生态学报, 2011, 35(8): 801–807. |
| [32] | 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937–3947. DOI:10.3321/j.issn:1000-0933.2008.08.054 |
| [33] | 李雪峰, 韩士杰, 胡艳玲, 赵玉涛. 长白山次生针阔混交林叶凋落物中有机物分解与碳、氮和磷释放的关系. 应用生态学报, 2008, 19(2): 245–251. |
| [34] | 黄昌勇. 土壤学. 北京: 中国农业出版社, 2000: 3–9. |
| [35] | Cleveland C C, Liptzin D. C:N:P stoichiometry in soil:is there a "Redfield ratio" for the microbial biomass?. Biogeochemistry, 2007, 85(3): 235–252. DOI:10.1007/s10533-007-9132-0 |
| [36] | Majdi H, Öhrvik J. Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in northern Sweden. Global Change Biology, 2004, 10(2): 182–188. DOI:10.1111/gcb.2004.10.issue-2 |
| [37] | McGroddy M E, Daufresne T, Hedin L O. Scaling of C:N:P stoichiometry in forests worldwide:implications of terrestrial Redfield-type ratios. Ecology, 2004, 85(9): 2390–2410. DOI:10.1890/03-0351 |
| [38] | Drenovsky R E, Richards J H. Critical N:P values:predicting nutrient deficiencies in desert shrublands. Plant and Soil, 2004, 259(1/2): 59–69. DOI:10.1023/B:PLSO.0000020945.09809.3d |
| [39] | Hedin L O. Physiology:plants on a different scale. Nature, 2006, 439(7079): 399–400. |
| [40] | 曾昭霞, 王克林, 刘孝利, 曾馥平, 宋同清, 彭晚霞, 张浩, 杜虎. 桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征. 植物生态学报, 2015, 39(7): 682–693. |