 2019, Vol. 39
2019, Vol. 39文章信息
- 邓巍, 王家亮, 刘李蕾, 房以好, 刘硕然, 杨晓燕, 肖文
- DENG Wei, WANG Jialiang, LIU Lilei, FANG Yihao, LIU Shuoran, YANG Xiaoyan, XIAO Wen
- 坡向尺度差异对大理苍山捕食线虫真菌分布格局的影响
- Slope's scale effect on the distribution patterns of nematode trapping fungi from Cangshan Mountain, Dali
- 生态学报. 2019, 39(17): 6452-6459
- Acta Ecologica Sinica. 2019, 39(17): 6452-6459
- http://dx.doi.org/10.5846/stxb201809282113
-
文章历史
- 收稿日期: 2018-09-28
- 网络出版日期: 2019-05-31






2. 阜阳市人民医院医院感染管理科, 阜阳 236000;
3. 大理大学农学与生物科学学院, 大理 671003;
4. 云南省高校洱海流域保护与可持续发展研究重点实验室, 大理 671003
2. Fu Yang People's Hospital Infection Management Section, Fuyang 236000, China;
3. School of Agriculture & Biology, Dali 671003, China;
4. The Key Laboratory of Yunnan State Education Department on Er'hai Lake Basin Protection and the Sustainable Development Research, Dali 671003, China
微生物是一类重要的生物类群, 是地球各生态系统物质循环的纽带, 并在土壤圈、生物圈、大气圈、水圈、岩石圈生物地球化学循环中扮演着不可或缺的角色[1-4]。长期以来, 人们认为微生物是呈随机分布的[5-6], 但随着研究的深入人们发现微生物类群的分布特征与其所处生境的空间异质性呈相关关系, 微生物存在空间分布并受到环境筛选作用的影响[7]。此前关于微生物空间分布格局是由当代环境条件(温度、降水、光照、土壤养分和pH)还是历史因素(地理阻隔、物理屏障、扩散限制等)决定的争论, 一直存在很大的分歧。如今的研究普遍开始认为环境条件和历史因素共同决定微生物的分布格局, 但两者的相对贡献率仍存在很多争议[8-11], 对微生物空间分布的驱动机制也仍未有清晰的定论。
坡向是十分重要的地理地形因子, 也是影响生物群落空间分布的重要因素, 坡向通过影响地面接收太阳辐射以及地面与盛行风的交角导致光、热、水、土壤矿物质等环境因子产生差异[12]。这些环境因子差异导致不同坡向间形成微型气候梯度, 生物群落结构的渐变也随之产生[13]。目前坡向对生物群落分布格局影响的研究主要集中于植物类群, 微生物类群的研究较少, 仅有少量学者对土壤细菌和丛枝根菌进行过描述性的报道, 已有研究大都基于单一尺度坡向开展[14-15]。
捕食线虫真菌是一类既可以营腐生生活又可以营养菌丝特化形成捕食器官(产黏性菌网的Arthrobotrys属, 产黏性球、黏性分枝的Dactylellina属, 产收缩环的Drechslerella属)来捕捉线虫的兼性菌。由于这类真菌在形态和功能上的特殊性以及在线虫生物防治中的潜在价值, 一直受到国内外真菌学研究者的重视, 并已经在菌种资源、分类、生态、分子与遗传、生理生化和生防菌剂开发与应用等方面开展了系统的研究[16]。研究资料显示捕食线虫真菌广泛分布于全球的土壤、水体和动物肠道甚至温泉等各类生境中[17]。由于捕食线虫真菌分离和纯化较为容易, 种类较少(共3属, 97种), 形态差异明显易于观察和鉴定[16], 因此捕食线虫真菌是研究微生物分布格局的理想对象。
鉴于微生物空间分布格局形成的机制尚不清晰, 坡向尺度差异对微生物分布格局的影响尚不明确, 本次研究以环境异质性较强、坡向尺度多样的大理苍山为研究区域, 以捕食线虫真菌为研究对象, 探究不同尺度坡向对捕食线虫真菌空间分布格局的影响。研究结果将有助于对微生物分布格局理论的完善, 促进对微生物多样性分布格局形成和维持机制的理解, 同时也有利于捕食线虫真菌资源的系统调查和种质资源的充分挖掘。
1 研究方法 1.1 研究区域苍山地处99°54′30″—100°12′12″E和25°33′30″—25°59′48″N之间, 由十九座山峰自北而南组成。自北而南的主要走向将苍山划分为东、南、西、北4个主要坡向, 同时十九峰中每一座山峰也将其划分为南、北两个次级坡向。苍山位于国家级自然保护区属横断山脉云岭山系, 处于南亚热带和中亚热带的交汇处, 形成了明显的垂直地带性和东西差异性[18-19]。苍山东西坡气温有明显差异, 形成一山分四季, 十里不同天的气候特征, 其植被类型多样, 生物多样性极为丰富、独特, 并具有明显的植物垂直分布带谱[20], 使苍山成为南北生物交汇过渡带, 以及生物的分布和分化中心, 为生物多样性最为丰富的地区之一。
本研究在大理苍山东、西坡向上的次级南、北坡向展开(图 1)。

|
| 图 1 采样点地图 Fig. 1 Sampling point map |
土样采自大理苍山东、西坡及其次级南、北坡4个坡向的裸露区。以海拔2100 m为起始每升高100 m设置一个采样点直至3500 m, 共设置60个采样点。每个采样点对角线5点取样(5—30 cm深土层)充分混匀取约200 g装于塑料封口袋, 带回实验室低温密封保存。共采集300份土样, 其中东、西坡分别采集150份土样, 次级南、北坡各75份。
1.3 培养基的制备参照文献[21], 配制玉米琼脂培养基(Corn meal agar medium,CMA), 用于分离捕食线虫真菌;土豆培养基(Potato dextrose agar medium,PDA), 用于菌株测序时的富营养培养。
1.4 诱饵线虫的制备用贝尔曼氏法制备全齿复活线虫(Panagrellus redivivus)幼虫悬液(每毫升约含线虫5000条)备用[17]。
1.5 分离纯化诱饵平板法[21]:在CMA平板上撒入0.5—1 g土样, 加入约5000条诱饵线虫。每份样品设置3个平行, 再将平板放置于(25±2)℃的环境中进行培养。三周后使用体视显微镜开始镜检, 同时使用无菌牙签转接捕食线虫真菌单个分生孢子到60 mm CMA平板上, 置于(26.5±1)℃温箱中培养7 d, 如此重复3—5次, 直至捕食线虫真菌纯培养物的获得。
1.6 捕食线虫真菌形态观察和捕食器官的诱导用透明胶带黏取捕食线虫真菌孢子和孢子梗, 放置在滴有吕氏美蓝染色液的载玻片上, 加盖盖玻片, 制成临时装片, 显微镜(40×)下镜检, 确定分生孢子和菌丝形态及分生孢子梗的类型。
用无菌刀在CMA培养基上挖一个2 cm×2 cm的观察室, 将菌株接入, (25±2)℃培养5—7 d, 待菌丝长满观察室后加入诱饵线虫, 12—24 h后观察捕食器官产生情况。
1.7 捕食线虫真菌的鉴定通过菌株形态特征及捕食器官类型进行形态学鉴定[21]。同时用PDA培养基富营养培养, 提取真菌总DNA并进行PCR扩增[22], 将产物委托上海铂尚生物技术有限公司进行纯化和测序。通过ITS(Internal transcribed spacer region of the ribosomal RNA gene, 核糖体RNA上的非转录间隔区)和TUB(β-tubulin gene, 微管蛋白编码基因)基因序列同源性分析进行分子生物学鉴定[23]。结合形态学和分子生物学确定种属。
1.8 数据处理检出率(Occurrence frequency, OF)=某个种出现的土样数/总土样数×100%。
总检出率=检出菌的土样数/总的土样数×100%。
Shannon-Wiener指数(He′)=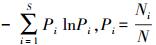
式中, S为物种数目;N为所有物种的个体数之和;Ni为第i个种个体数量。
非参数检验(Nonparametric tests):以200 m为梯度将研究区域分为4个海拔段, 对各坡向捕食线虫真菌物种数进行非参数检验。
群落相似性指数:Jaccard指数= 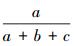
运用EXCEL和SPSS对原始数据进行整理分析。
2 实验结果 2.1 不同坡向捕食线虫真菌的物种组成与分布本次研究于苍山共分离和鉴定出3属12种57株捕食线虫真菌, 其中苍山东坡共分离到2属8种24株, 次级南、北坡分别分离到8种15株和5种9株;在西坡共分离到3属11种33株, 次级南、北坡分别分离到8种19株和8种14株(图 2)。

|
| 图 2 不同坡向捕食线虫真菌的物种组成与分布 Fig. 2 Species composition and distribution of nematode-trapping fungi from different slopes Drechslerella:小掘氏孢属;Dactylellina:亚隔指孢属;Arthrobotrys:节丛孢属;Dre.aphrobrochum:泡环单顶孢;Dac.parvicolle:细颈单顶孢;Dac.drechsleri:掘氏单顶孢;Dac. ellipsosporum:椭圆单顶孢;A. guizhouensis:贵州节丛孢;A. microscaphoide:小舟单顶孢;A. thaumasium:奇妙单顶孢;A. robusta:强力节丛孢;A. fusiformis:纺锤单顶孢;A. conoides:圆锥节丛孢;A. musiformis:弯孢节丛孢;A. oligospora少孢节丛孢 |
西坡捕食线虫真菌检出数大于东坡, 东、西坡的次级南坡捕食线虫真菌检出数大于次级北坡。产黏性菌网的Arthrobotrys属和产黏性球、黏性分枝的Dactylellina属在东、西坡及其次级南、北坡均有分布, 但Arthrobotrys属要分布于东坡, Dactylellina属主要分布于西坡, 而Drechslerella属仅在西坡有分布(图 2)。
东坡的次级南、北坡少孢节丛孢(Arthrobotrys oligospora)检出率高于其他种, 为东坡的优势种, 而椭圆单顶孢(Dactylellina ellipsosporum)在西坡的次级南、北坡的检出率最高, 为西坡的优势种。少孢节丛孢(A.oligospora)、弯孢节丛孢(Arthrobotrys musiformis)、圆锥节丛孢(Arthrobotrys conoides)和椭圆单顶孢(Dac.ellipsosporum)坡向分布范围最广, 其在东、西坡的次级南、北坡均有分布。纺锤单顶孢(Arthrobotrys fusiformis)的坡向分布范围次之, 其主要分布于东坡的次级南、北坡和西坡的次级北坡。掘氏单顶孢(Dactylellina drechsleri)和细颈单顶孢(Dactylellina parvicolle)主要分布于东、西坡的次级南坡。泡环单顶孢(Drechslerella aphrobrochum)只分布于西坡。贵州节丛孢(Arthrobotrys guizhouensis)、强力节丛孢(Arthrobotrys robusta)和小舟单顶孢(Arthrobotrys microscaphoide)要分布于西坡, 其中强力节丛孢(A.robusta)和小舟单顶孢(A. microscaphoide)主要分布于西坡的次级南坡, 贵州节丛孢(A.guizhouensis)主要分布于西坡的次级北坡。而奇妙单顶孢(Arthrobotrys thaumasium)主要分布于东坡的次级南坡(图 2)。
2.2 不同坡向捕食线虫真菌的检出率和多样性指数对不同坡向分布区的捕食线虫真菌多样性指数和检出率进行比较, 结果显示西坡捕食线虫真菌的检出率、Shannon-Wiener指数均大于东坡。而对次级南、北坡进行比较, 结果显示次级南坡捕食线虫真菌的检出率、Shannon-Wiener指数均大于次级北坡(图 3)。

|
| 图 3 各坡向捕食线虫真菌的检出率和多样性指数 Fig. 3 Detection rate and diversity index of nematode-trapping fungi in different slopes |
苍山东、西坡及其次级南、北坡间捕食线虫真菌物种组成均存在差异性(Z=-1.98, P=0.046; Z=-2.38, P=0.017)。计算不同坡向Jaccard指数, 比较群落相似程度。结果显示苍山次级南、北坡间的群落差异大于东、西坡间(0.5<0.58);西坡次级南、北坡间的群落差异最大(0.45);东坡次级南坡与西坡次级南坡间Jaccard指数为0.54;东坡次级北坡与次级南坡(0.63)、东坡次级北坡与西坡次级北坡(0.71)间的群落较为相似。
3 分析与讨论 3.1 苍山不同坡向捕食线虫真菌分布差异研究表明, 坡向会在不同程度上影响土壤的温度、含水量等理化性质[24], 其中土壤含水量在不同坡向上的变化最为明显[25-26], 西坡的土壤含水量通常高于东坡[27], 这种不同坡向上微小气候的不同会导致土壤微生物生物量存在坡向差异[28]。本研究也发现, 苍山东、西坡捕食线虫真菌的物种丰富度和多样性均是西坡大于东坡, 东、西坡之间捕食线虫真菌的分布存在差异。苍山东坡属于亚热带气候类型, 而西坡属于中亚热带气候类型。西坡立体气候跨度比东坡大, 气候干湿季分明, 雨量充沛, 立体气候明显, 年日照时数略大于东坡, 月平均气温均高于东坡[19]。这种水、热条件的差异是导致东、西坡上捕食线虫真菌物种丰富度和物种多样性不同的主要原因。同时, 土壤微生物作为土壤有机质和养分循环的动力, 能够对周围的环境变化迅速做出反应[29], 因此, 东、西坡向上水、热条件的差异, 势必会导致不同坡向上土壤中微生物的种群结构发生改变。这种改变一方面会影响到其他生物种群如线虫的群落结构, 进而影响到捕食线虫真菌的分布[30];另一方面则会影响到土壤pH、营养元素(N、P、K)等, 从而影响到土壤中的捕食线虫真菌的种群结构[17]。
3.2 尺度效应与捕食线虫真菌分布格局自然界的所有格局和规律都具有尺度依赖性[31]。Hanson等通过比较一系列微生物生物地理分布格局发现, 空间尺度是导致微生物分布格局差异的主要诱因[32]。但是, 在研究生物分布格局和生物多样性时, 选取不同的空间尺度所得到的结果存在差异, 有的甚至截然相反[33-35]。本研究结果表明, 苍山捕食线虫真菌分布格局在较大尺度(东、西坡)和较小尺度(次级南、北坡)均存在差异;小尺度坡向间捕食线虫真菌群落差异性更大。曹鹏等认为造成不同尺度微生物分布格局差异的原因是群落构建过程不同, 在小尺度下扩散过程更容易发生[36]。这种扩散限制的改变导致小尺度捕食线虫真菌群落构建过程发生改变, 次级南、北坡间因此产生了更大的群落差异。因此小尺度坡向间地理屏障对捕食线虫真菌分布格局的影响大于大尺度坡向间环境异质性的作用。在研究捕食线虫真菌分布格局以及生物多样性形成和维持机制时, 更应该关注诸如次级坡向这样的小空间尺度。
3.3 坡向因子与群落构建生态群落构建即生物多样性形成和维持机制长期以来是生态学研究的核心。基于环境筛选(确定性过程)的生态位理论和基于扩散限制(随机性过程)的中性理论是解释群落构建机制最为经典的理论, 同时也是最具有争议性的理论。传统的群落构建理论认为, 像微生物这样扩散力强、种群数量庞大、具有生长迅速和休眠特征的种群群落构建机制主要受到环境筛选作用的影响[37]。但越来越多学者认为这两种看似矛盾的理论对于群落构建的解释并不排斥, 生物多样性形成和维持以及生物地理分布格局是环境筛选和扩散限制共同作用的结果[38-39]。有研究认为坡向是影响群落构建过程强有力的环境筛选因素[40], 苍山不同坡向上的捕食线虫真菌群落结构明显不同, 这再一次印证了环境筛选对于群落构建的作用。产收缩环的Drechslerella属捕食线虫真菌仅在苍山西坡有分布, 这也说明东、西坡间存在一定的扩散限制, 扩散限制同样影响到了群落构建的过程。苍山不同坡向上捕食线虫真菌的分布格局表明环境筛选和扩散限制均会影响到捕食线虫真菌的群落构建过程, 基于环境异质性较强的区域开展不同坡向上微生物群落构建研究将有助于更加准确的理解土壤微生物的生物多样性形成和维持以及分布格局, 也有助于探索环境筛选和扩散限制在微生物群落构建过程中的相对贡献。
4 结论坡向是影响苍山捕食线虫真菌空间分布格局的驱动因子之一。环境筛选和扩散限制均会影响到捕食线虫真菌的群落构建过程, 从而导致次级坡向间捕食线虫真菌空间分布格局发生改变。因此在微生物分布格局及资源调查的研究中, 需要考虑小尺度坡向对群落结构造成的影响。
| [1] |
Amann R I, Ludwig W, Schleifer K H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiological Reviews, 1995, 59(1): 143-169. |
| [2] |
Whitman W B, Coleman D C, Wiebe W J. Prokaryotes:the unseen majority. Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(12): 6578-6583. DOI:10.1073/pnas.95.12.6578 |
| [3] |
Balser T C. Linking Soil Microbial Communities and Ecosystem Functioning[D]. Berkeley: University of California, 2000.
|
| [4] |
Wardle D A. Communities and Ecosystems:Linking the Aboveground and Belowground Components. Princeton: Princeton University Press, 2002.
|
| [5] |
MacArthur R H, Wilson E O. The theory of Island Biogeography. Princeton: Princeton University Press, 1967.
|
| [6] |
Noguez A M, Arita H T, Escalante A E, Forney L J, García-Oliva F, Souza V. Microbial macroecology:highly structured prokaryotic soil assemblages in a tropical deciduous forest. Global Ecology and Biogeography, 2005, 14(3): 241-248. DOI:10.1111/j.1466-822X.2005.00156.x |
| [7] |
Bell T, Ager D, Song J I, Newman J A, Thompson I P, Lilley A K, Van Der Gast C J. Larger islands house more bacterial taxa. Science, 2005, 308(5730): 1884-1884. DOI:10.1126/science.1111318 |
| [8] |
Fierer N, Jackson R B. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(3): 626-631. DOI:10.1073/pnas.0507535103 |
| [9] |
Ge Y, He J Z, Zhu Y G, Zhu Y G, Zhang J B, Xu Z H, Zhang L M, Zheng Y M. Differences in soil bacterial diversity:driven by contemporary disturbances or historical contingencies?. The ISME Journal, 2008, 2(3): 254-264. DOI:10.1038/ismej.2008.2 |
| [10] |
Garcia-Pichel F, Loza V, Marusenko Y, Mateo P, Potrafka R M. Temperature drives the continental-scale distribution of key microbes in topsoil communities. Science, 2013, 340(6140): 1574-1577. DOI:10.1126/science.1236404 |
| [11] |
Ranjard L, Dequiedt S, Chemidlin Prévost-Bouré N, Thioulouse J, Saby N P, Lelievre M, Maron P A, Morin F E, Bispo A, Jolivet C, Arrouays D, Lemanceau P. Turnover of soil bacterial diversity driven by wide-scale environmental heterogeneity. Nature Communications, 2013, 4: 1434. DOI:10.1038/ncomms2431 |
| [12] |
Auslander M, Nevo E, Inbar M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. Journal of Arid Environments, 2003, 55(3): 405-416. DOI:10.1016/S0140-1963(02)00281-1 |
| [13] |
Gong X, Brueck H, Giese K M, Zhang L, Sattelmacher B, Lin S. Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia, China. Journal of Arid Environments, 2008, 72(4): 483-493. DOI:10.1016/j.jaridenv.2007.07.001 |
| [14] |
Hazard C, Gosling P, Van Der Gast C J, Mitchell D T, Doohan F M, Bending G D. The role of local environment and geographical distance in determining community composition of arbuscular mycorrhizal fungi at the landscape scale. The ISME Journal, 2013, 7(3): 498-508. DOI:10.1038/ismej.2012.127 |
| [15] |
Chu H Y, Xiang X J, Yang J, Adams J M, Zhang K P, Li Y T, Shi Y. Effects of Slope Aspects on Soil Bacterial and Arbuscular Fungal Communities in a Boreal Forest in China. Pedosphere, 2016, 26(2): 226-234. DOI:10.1016/S1002-0160(15)60037-6 |
| [16] |
Zhang K Q, Hyde K D. Nematode-Trapping Fungi. Dordrecht, Netherlands: Springer, 2014.
|
| [17] |
Liu J, Wang J N, Gao G H, Bartlam M G, Wang Y Y. Distribution and diversity of fungi in freshwater sediments on a river catchment scale. Frontiers in Microbiology, 2015, 6: 329. |
| [18] |
大理白族自治州苍山保护管理局. 苍山志. 昆明: 云南民族出版社, 2008.
|
| [19] |
杨金涛. 大理苍山气候与旅游资源的分析研究. 民营科技, 2010, 7: 148-148. |
| [20] |
冯建孟, 李继红, 王浩波, 董晓东. 苍山西坡漾濞地区种子植物区系成分的垂直分布格局. 楚雄师范学院学报, 2010, 25(3): 48-52, 58-58. DOI:10.3969/j.issn.1671-7406.2010.03.008 |
| [21] |
李天飞, 张克勤, 刘杏忠. 食线虫菌物分类学. 北京: 中国科学技术出版社, 2000.
|
| [22] |
张颖.捕食线虫丝孢菌代表种的群体遗传学研究[D].昆明: 云南大学, 2010.
|
| [23] |
袁长婷.核糖体RNA基因间隔区ITS及IGS在真菌分子生物学鉴定和分型中应用[D].上海: 第二军医大学, 2001.
|
| [24] |
Pen-Mouratov S, Berg N, Genzer N, Ukabi S, Shargil D, Steinberger Y. Do slope orientation and sampling location determine soil biota composition?. Frontiers of Biology in China, 2009, 4(3): 364-375. DOI:10.1007/s11515-009-0023-8 |
| [25] |
徐长林. 坡向对青藏高原东北缘高寒草甸植被构成和养分特征的影响. 草业学报, 2016, 25(4): 26-35. |
| [26] |
杨婷.坡向对土壤微生物特征与生态化学计量特征的影响研究[D].杨凌: 西北农林科技大学, 2016.
|
| [27] |
肖智勇, 李根前, 代光辉, 李孙玲, 李甜江. 黄土高原不同坡向中国沙棘种群生物量投资与分配. 东北林业大学学报, 2011, 39(5): 44-46, 57-57. DOI:10.3969/j.issn.1000-5382.2011.05.014 |
| [28] |
Sidari M, Ronzello G, Vecchio G, Muscolo A. Influence of slope aspects on soil chemical and biochemical properties in a Pinus laricio forest ecosystem of Aspromonte (Southern Italy). European Journal of Soil Biology, 2008, 44(4): 364-372. DOI:10.1016/j.ejsobi.2008.05.001 |
| [29] |
Avidano L, Gamalero E, Cossa G P, Carraro E. Characterization of soil health in an Italian polluted site by using microorganisms as bioindicators. Applied Soil Ecology, 2005, 30(1): 21-33. DOI:10.1016/j.apsoil.2005.01.003 |
| [30] |
Wang X, Li G H, Zou C G, Ji X L, Liu T, Zhao P J, Liang L M, Xu J P, An Z Q, Zheng X, Qin Y K, Tian M Q, Xu Y Y, Ma Y C, Yu Z F, Huang X W, Liu S Q, Niu X M, Yang J K, Huang Y, Zhang K Q. Bacteria can mobilize nematode-trapping fungi to kill nematodes. Nature Communications, 2014, 5: 5776. DOI:10.1038/ncomms6776 |
| [31] |
吕一河, 傅伯杰. 生态学中的尺度及尺度转换方法. 生态学报, 2001, 21(12): 2096-2105. DOI:10.3321/j.issn:1000-0933.2001.12.019 |
| [32] |
Dolan J R. Microbial biogeography?. Journal of Biogeography, 2006, 33(2): 199-200. DOI:10.1111/j.1365-2699.2005.01406.x |
| [33] |
Hawkins B A, Porter E E, Diniz-Filho J A F. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology, 2003, 84(6): 1608-1623. DOI:10.1890/0012-9658(2003)084[1608:PAHAPO]2.0.CO;2 |
| [34] |
Qian H, Ricklefs R E. A latitudinal gradient in large-scale beta diversity for vascular plants in North America. Ecology Letters, 2007, 10(8): 737-744. DOI:10.1111/j.1461-0248.2007.01066.x |
| [35] |
Villalobos F, Lira-Noriega A, Soberón J, Arita H T. Range-diversity plots for conservation assessments:Using richness and rarity in priority setting. Biological Conservation, 2013, 158: 313-320. DOI:10.1016/j.biocon.2012.10.002 |
| [36] |
曹鹏, 贺纪正. 微生物生态学理论框架. 生态学报, 2015, 35(22): 7263-7273. |
| [37] |
马转转, 乔沙沙, 曹苗文, 周永娜, 刘晋仙, 贾彤, 李毳, 柴宝峰. 环境选择和扩散限制驱动温带森林土壤细菌群落的构建. 应用生态学报, 2018, 29(4): 1179-1189. |
| [38] |
Hubbell S P. Neutral theory and the evolution of ecological equivalence. Ecology, 2006, 87(6): 1387-1398. DOI:10.1890/0012-9658(2006)87[1387:NTATEO]2.0.CO;2 |
| [39] |
Adler P B, Hillerislambers J, Levine J M. A niche for neutrality. Ecology Letters, 2007, 10(2): 95-104. DOI:10.1111/j.1461-0248.2006.00996.x |
| [40] |
李新娥.亚高寒草甸阳坡-阴坡梯度上植物功能性状及群落构建机制研究[D].兰州: 兰州大学, 2011.
|