 2019, Vol. 39
2019, Vol. 39文章信息
- 张勇, 崔海军, 张银烽, 杨苑君, 岳亮亮, 肖德荣
- ZHANG Yong, CUI Haijun, ZHANG Yinfeng, YANG Yuanjun, YUE Liangliang, XIAO Derong
- 柳杉种植对黔西南喀斯特山区金发藓沼泽植物群落和储水功能的影响
- Effects of artificial Cryptomeria fortunei forests on the plant community and water content of a Polytrichum commune bog in a karst region in southwestern Guizhou Province, China
- 生态学报. 2019, 39(17): 6304-6313
- Acta Ecologica Sinica. 2019, 39(17): 6304-6313
- http://dx.doi.org/10.5846/stxb201805040996
-
文章历史
- 收稿日期: 2018-05-04
- 网络出版日期: 2019-05-31






2. 云南农业大学水利学院, 昆明 650201
2. College of Water Resources and Hydraulic Engineering, Yunnan Agricultural University, Kunming 650201, China
由于强烈的水蚀作用和日趋增强的人为活动干扰, 云贵高原喀斯特地貌分布区面临严峻的石漠化问题[1-4], 三成以上石漠化区域的生态环境敏感性等级在高度敏感以上[5]、植物群落结构简单化、物种多样性显著下降[6]。此外, 近年来日趋频发的极端天气现象更使喀斯特山区生态系统的稳定性面临极大风险[7], 严重制约了当地的社会经济发展[8]。为遏制石漠化问题, 喀斯特山区开展了大量的生态治理措施[9-10]。其中, 植树造林是一项重要举措[11-12]。
根际土壤层蓄水是森林涵养水源的主要途径[13-15], 但乔木层林龄增加常导致土壤持水能力波动[16-17]。藓类植物覆盖可有效降低林下土壤水分蒸发, 对稳定表层土壤含水量有积极作用[18-19]。但有研究表明杉木林龄增加可导致林下草本层植物丰富度和生物量下降[20-21], 杉木林下藓类植物可能也会受到类似干扰。
藓类沼泽是一类沼泽湿地, 零星分布在我国东北山地和西南山地[22]。藓类植物具有良好的持水性能, 对维持区域水循环和水文功能有重要作用[23-24]。小规模成片分布在黔西南喀斯特山区的藓类沼泽对保障当地生活、农业生产用水有重要作用[25-26]。1998年前后黔西南喀斯特山区种植了大量的柳杉(Cryptomeria fortunei), 截至目前, 柳杉种植对该区域藓类沼泽植物群落和储水功能影响的研究尚少见报道。本研究拟在黔西南喀斯特山区典型金发藓沼泽分布区探讨人工柳杉林与林下金发藓沼泽植物群落以及沼泽储水能力的关系。本研究的研究假设为:1)人工柳杉林对林下金发藓植物群落有负面影响;2)人工柳杉林可降低林下金发藓沼泽的储水能力。
1 材料和方法 1.1 实验设计野外调查娘娘山位于贵州省六盘水市, 最高海拔2400 m, 为典型喀斯特高原山地类型, 该区域年均温15.2℃, 年均降雨量1413.6 mm, 雨热同期, 属典型亚热带季风气候[27]。本研究野外调查样地位于娘娘山顶台地, 地势较平坦(海拔介于2150—2250 m), 分布着大量的金发藓(Polytrichum commune)群落, 各调查样地的坡向、坡度均相似(图 1)。20世纪90年代末, 娘娘山顶开始大面积种植柳杉(C. fortunei), 调查区域的柳杉长势较均匀:经过近20年的生长, 柳杉的平均胸径为(9.5±2.7)cm, 平均最大冠幅为(230±40)cm。为厘清人工柳杉林对金发藓沼泽植物群落结构和储水功能的影响, 本研究分别于2017年7月(雨季)和2018年1月(干季)对娘娘山顶不同密度柳杉林下金发藓群落进行群落调查, 并采集表层土壤样品(0—15 cm)以测定土壤含水率。雨季, 从娘娘山顶东部到西北部设置调查样线, 沿调查样线, 根据柳杉林密度设置9个20 m×20 m的调查样地;干季, 参照雨季调查样地分布, 设置6个20 m×20 m的调查样地;在雨季和干季的调查中, 均在山顶西北部没有柳杉种植的区域设置3个10 m×10 m的对照样地, 雨季的对照样地记作CKR, 干季的对照样地记作CKD(图 1)。在每个调查样地内随机设置3—5个0.5 m×0.5 m的调查样方, 在每个样方内, 1)记录草本层(包括金发藓)的物种数量、盖度和高度。调查样地内出现的植物种在表 1中列出, 这些物种基本都属于本土物种;2)齐地剪取金发藓的地上部分并密封带回实验室;3)用口径5 cm的土钻收集0—15 cm的土壤样品并密封带回实验室。

|
| 图 1 雨季和干季调查样地分布 Fig. 1 Sampling sites in a rainy season and a dry season CKR:雨季对照样地Control Check in a Rainy Season;R:雨季Rainy Season;CKD:干季对照样地Control Check in a Dry Season;D:干季Dry Season |
| 中文名Chinese name | 拉丁名Latin name |
| 金发藓 | Polytrichum commune |
| 泥炭藓 | Sphagnum palustre |
| 大灰藓 | Hypnum plumaeforme |
| 委陵菜 | Potentilla chinensis |
| 香青 | Anaphalis sinica |
| 天胡荽 | Hydrocotyle sibthorpioides |
| 过路黄 | Lysimachia christinae |
| 鞭打绣球 | Hemiphragma heterophyllum |
| 小二仙草 | Haloragis micrantha |
| 薄叶新耳草 | Neanotis hirsuta |
| 蛛丝毛蓝耳草 | Cyanotis arachnoidea |
| 石松 | Lycopodium japonicum |
| 鹅观草 | Roegneria kamoji |
| 野青茅 | Deyeuxia arundinacea |
| 李氏禾 | Leersia hexandra |
| 早熟禾 | Poa annua |
室内实验分别称量野外收集的金发藓植物样品和土壤样品的鲜重/湿重, 然后烘干至恒重并称量干重。
1.2 数据处理物种综合优势度(Summed dominance ratio, SDR)通过分种盖度和高度计算调查样方内各物种在群落中的优势度, 计算方式如下[28]:
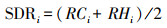
|
式中, RCi= Ci× 100/∑Ci;RHi=Hi× 100/∑Hi。
式中, RCi为物种i的相对盖度;∑Ci为群落中所有物种盖度之和;RHi为物种i的相对高度;∑Hi为群落中所有物种高度之和。某个样方内, 所有物种的SDR之和为1, 某物种的SDR越大, 表明该物种在群落中越占优。优势种的SDR变化可指示群落结构变化。本研究用金发藓的SDR作为反应群落结构变化的一个指数。
含水率及储水量称量单位面积金发藓的鲜重(记作Wp, 单位为g/m2);称量单位体积表层土壤的鲜重, 即土壤湿密度(记作ρs, 单位为g/cm3)。用金发藓植物鲜重和干重计算金发藓含水率(记作Rp);用表层土壤的湿重和干重计算土壤含水率(记作Rs)。用金发藓鲜重和金发藓含水率计算单位面积的金发藓含水量(记作Cp), 计算公式为:Cp= Wp×Rp。用土壤湿密度和土壤含水率计算单位体积(1 m2×0.15 m)的土壤含水量(记作Cs), 计算公式为:Cs=ρs×Rs。金发藓沼泽生态系统单位面积的储水量记作Csystem, 其计算方式为:Csystem= Cp+Cs。
统计分析雨季可观测到金发藓沼泽植物群落中最多的植物种数, 干季除藓类植物外大部分物种的地上部分均枯萎, 无法完整记录植物群落的组成情况, 因此采用雨季的调查数据分析柳杉林密度和金发藓沼泽植物群落盖度、平均高度、植物物种丰富度和金发藓SDR的回归关系。在本次调查中柳杉林密度介于800—6700株/hm2, 根据实地调查的情况, 将柳杉林密度分为低密度(密度≤3000株/hm2)和高密度(>3000株/hm2)两组进行统计分析, 按以上标准将柳杉林调查样地分为雨季低密度柳杉林(记作LSR1)、雨季高密度柳杉林(LSR2)、干季低密度柳杉林(LSD1)和干季高密度柳杉林(LSD2)。通过单因素方差分析(one-way ANOVA)分析不同密度柳杉林对金发藓沼泽植物群落盖度、平均高度、物种数、金发藓SDR和金发藓沼泽储水能力的影响。计算不同密度柳杉林下金发藓植物含水率、表层土壤含水率、单位面积金发藓含水量、单位面积土壤含水量和单位面积生态系统含水量在季节间的变异系数(Coefficient of variation), 以分析不同密度柳杉林下金发藓沼泽储水能力的稳定性。回归分析和方差分析均在IBM SPSS Statistics 19.0中完成。
2 结果 2.1 柳杉对金发藓沼泽植物群落的影响对植物群落盖度和高度的影响柳杉密度低于3000株/hm2时(即LSR1), 林下的金发藓群落盖度与对照(CKR)没有显著差异;柳杉密度高于3000株/hm2时(即LSR2), 林下的金发藓群落盖度显著降低, 其仅有对照样地的45.6%左右。与群落盖度情况相似, 低密度柳杉林下金发藓群落平均高度与对照没有显著差异, 但当柳杉密度较高时, 金发藓群落平均高度显著低于对照(图 2)。

|
| 图 2 柳杉种植对金发藓群落盖度和平均高度的影响 Fig. 2 Effects of Cryptomeria fortunei forest on the cover and average height of Pogonatumin flexum communities LSR1:柳杉密度≤3000株/hm2的雨季样地Sampling sites with a low density of Cryptomeria fortunei forest (density ≤3000 individuals per hm2) in a rainy season;LSR2:柳杉密度>3000株/hm2的雨季样地Sampling sites with a high density of Cryptomeria fortunei forest (density > 3000 individuals per hm2) in a rainy season |
对雨季调查的9块柳杉样地进行进一步分析, 发现随柳杉密度增加, 金发藓群落盖度和平均高度均显著降低, 这两个指标与柳杉密度之间存在明显的线性回归关系(图 3)。

|
| 图 3 柳杉密度与金发藓群落盖度和平均高度的关系 Fig. 3 Relationships between the density of the Cryptomeria fortunei forest and the cover and average height of Pogonatumin flexum communities |
对群落物种数和金发藓优势度的影响与对照相比, 无论是低密度柳杉林还是高密度柳杉林均显著降低了金发藓群落中的植物物种数。不同密度柳杉林下草本层群落中, 金发藓在群落中的优势度(即金发藓SDR)没有显著变化, 即柳杉种植对金发藓在草本层群落中的优势地位没有显著影响(图 4)。

|
| 图 4 柳杉种植对金发藓群落物种数和金发藓SDR的影响 Fig. 4 Effects of the Cryptomeria fortunei forest on the specie richness of Pogonatumin flexum communities and the SDR of P. commune SDR:综合优势度Summed Dominance Ratio |
在柳杉分布区, 随柳杉密度增加林下金发藓群落的植物种数显著降低, 两者存在明显的回归关系。与物种数的变化趋势不同, 随柳杉密度增加, 金发藓SDR呈现先增加后降低的特征, 即柳杉密度和金发藓SDR之间是显著的二次曲线关系。由拟合方程可推知, 当柳杉密度约为2400株/hm2时, 金发藓在群落中的优势度最大(图 5)。

|
| 图 5 柳杉密度与金发藓群落植物种数和金发藓SDR的关系 Fig. 5 Relationships between the density of Cryptomeria fortunei forests and the specie richness of Pogonatumin flexum communities and the SDR of P. commune |
在没有柳杉种植的区域, 雨季和干季金发藓植物含水率均在52%左右, 两者没有显著差异。与对照相比, 雨季和干季不同密度柳杉林下金发藓植物含水率平均各增加约24%, 呈显著增加的特征(图 6)。

|
| 图 6 柳杉对金发藓植物含水率和表层土壤含水率的影响 Fig. 6 Effects of the Cryptomeria fortunei forest on water content of the Pogonatumin flexum and surface soils LSD1:柳杉密度≤3000株/hm2的干季样地Sampling sites with a low density of Cryptomeria fortunei forest (density ≤3000 individuals per hm2) in a dry season;LSD2:柳杉密度>3000株/hm2的干季样地Sampling sites with a high density of Cryptomeria fortunei forest (density > 3000 individuals per hm2) in a dry season |
雨季, 对照样地表层土壤含水率近80%, 与其相比, 柳杉种植区表层土壤含水率显著降低, 其中高密度柳杉林下表层土壤含水率下降最明显, 降低了近1/3。干季, 对照样地的表层土壤含水率约55%, 柳杉种植区表层土壤含水率与对照样地没有显著差异(图 6)。
在不同的调查区域, 表层土壤含水率表现出明显的季节差异性。在对照样地中, 干季表层土壤含水率显著低于雨季。在柳杉种植区, 干季柳杉林(包括低密度和高密度)林下表层土壤含水率显著低于雨季低密度柳杉林下表层土壤含水率, 但与雨季高密度柳杉林下表层土壤含水率没有显著差异(图 6)。
随柳杉密度增加, 金发藓植物含水率和表层土壤含水率在干季的变化趋势与在雨季的变化趋势相反。雨季, 随柳杉密度增加, 金发藓植物含水率和表层土壤含水率均显著降低, 它们之间存在显著的线性变化关系。干季, 随柳杉密度增加, 金发藓植物含水率和表层土壤含水率均呈线性增加趋势(图 7, 图 8)。

|
| 图 7 柳杉密度与金发藓含水率在雨季和干季的关系 Fig. 7 Relationships between the density of the Cryptomeria fortunei forest and water contents of Pogonatumin flexum in a rainy season and a dry season |

|
| 图 8 柳杉密度与表层土壤含水率在雨季和干季的关系 Fig. 8 Relationships between the density of the Cryptomeria fortunei forest and surface soil water contents in a rainy season and a dry season |
表层土壤含水量柳杉种植未对表层土壤的湿密度产生显著影响(表 2)。在雨季, 对照样地(柳杉密度为0)表层土壤含水量与低密度柳杉林下表层土壤含水量无显著差异, 但显著高于高密度柳杉林下表层土壤含水量(表 2)。在干季, 与对照相比, 柳杉种植显著增加了表层土壤含水量(表 2)。
| 样地* Plots |
表层土壤含水量 Water content of surface soils |
金发藓含水量 Water content of P. flexum |
系统储水量 Water content of the system/ (kg/m2) |
|||
| 湿密度 Natural density/ (g/cm3) |
含水量 Water content/ (kg/(0.15 m×1 m2)) |
鲜重 Fresh weight/ (g/m2) |
含水量 Water content/ kg/m2) |
|||
| CKR | 1.12±0.11a | 134.2±13.6a | 1255.8±41.4b | 0.66±0.02b | 134.8±13.6a | |
| LSR1 | 1.00±0.12a | 101.3±14.9ab | 1987.2±132.7a | 1.44±0.12a | 102.7±14.8ab | |
| LSR2 | 0.95±0.14a | 80.2±9.0b | 1274.9±68.8b | 0.88±0.06b | 81.1±9.0b | |
| CKD | 0.92±0.12a | 66.0±6.3b | 1293.0±148.0a | 0.65±0.18a | 66.6±6.4b | |
| LSD1 | 1.28±0.12a | 91.5±4.9a | 1686.1±181.2a | 1.12±0.15a | 88.1±2.5a | |
| LSD2 | 1.33±0.06a | 108.1±3.8a | 1346.5±126.1a | 1.00±0.04a | 106.8±5.3a | |
| * CKR/CKD:Control plots in a rainy/dry season;LSR1/LSD1:柳杉密度≤3000株/hm2的雨季/干季样地Sampling sites with a low density of Cryptomeria fortunei forest (density ≤3000 individuals per hm2) in a rainy/dry season;LSR2/LSD2:柳杉密度>3000株/hm2的雨季/干季样地Sampling sites with a high density of Cryptomeria fortunei forest (density > 3000 individuals per hm2) in a rainy/dry season | ||||||
金发藓含水量在雨季, 低密度柳杉林下单位面积金发藓鲜重和金发藓含水量显著高于对照样地和高密度柳杉林;在干季, 柳杉种植区域单位面积的金发藓鲜重和含水量与对照区域没有显著差异(表 2)。
金发藓沼泽储水量单位面积表层土壤含水量大概是单位面积金发藓含水量的100倍, 是整个金发藓沼泽储水量的主要贡献者(表 2)。在雨季, 对照样地的沼泽储水量与低密度柳杉林无显著差异, 但显著高于高密度柳杉林;在干季, 柳杉种植区域的金发藓沼泽储水量显著高于对照样地(表 2)。
与对照相比, 柳杉分布区的表层土壤含水率、金发藓含水率、表层土壤含水量、金发藓含水量和系统储水量的变异系数均降低(表 3), 说明柳杉种植有利于维持金发藓沼泽储水功能的稳定性。特别地, 对维持表层土壤含水量和系统储水量稳定性而言, 低密度柳杉林比高密度柳杉林效果好(表 3)。
| 柳杉密度 Density of the C. fortunei |
变异系数The coefficient of variation | ||||
| 金发藓含水率 WA of P. flexum |
表层土壤含水率 WA of surface soils |
表层土壤含水量 WC of surface soils |
金发藓含水量 WC of P. flexum |
系统储水量 WC of the system |
|
| 对照(密度=0) Control (density=0) | 0.016 | 0.262 | 0.408 | 0.308 | 0.405 |
| 低密度(0 < 密度≤3000株/hm2) Low density (0 < density ≤3000 individuals per hm2) |
0.017 | 0.229 | 0.194 | 0.214 | 0.191 |
| 高密度(密度>3000株/hm2) High density (>3000 individuals per hm2) |
0.0003 | 0.003 | 0.239 | 0.135 | 0.237 |
黔西南的藓类沼泽主要分布在海拔较高的喀斯特山区, 其生境具有湿冷的特征。柳杉具有良好的抗冻性能, 因此能在藓类沼泽的湿冷环境中生长[29]。本研究发现, 人工种植柳杉后娘娘山顶金发藓植物群落的盖度和平均高度均随柳杉密度增加而显著下降, 该结果与长白山地区天然森林冠层郁蔽度和林下藓类植物群落特征的研究结果相同[30-31]。这说明, 在我国藓类沼泽集中分布的两个区域, 森林(无论是天然林还是人工林)对林下藓类植物群落外貌结构的负面影响是类似的。
由于生态系统的脆弱性, 黔西南喀斯特山区是贵州省生物多样性较低的区域[32-33], 而生物多样性是维持生态系统稳定的重要机制之一[34]。因此保护和恢复生物多样性是黔西南卡斯特山区进行生态治理时重点关注的问题[35]。本研究发现, 种植柳杉后金发藓植物群落的物种多样性显著下降, 这种变化无疑会对金发藓群落的稳定性产生负面影响。且有研究发现柳杉林易受柳杉云毛虫(Hoenimnema roesleri)危害[36]。据此推测, 柳杉林下藓类植物群落由于物种数下降会变得不耐环境变化, 一旦柳杉林遭受病虫害, 在明显的干、湿季交替影响下, 娘娘山顶喀斯特荒漠化的进程会加剧。综上, 本研究的第一个研究假设得到验证, 即:人工柳杉林对林下金发藓植物群落有负面影响。
藓类沼泽的储水能力包括藓类植物的储水能力和沼泽土壤/泥炭层的储水能力。本研究发现, 土壤含水量是藓类沼泽储水量的主要贡献者, 该结果与太行山区的研究结论一致[15]。无论雨季和干季, 本研究中人工柳杉林林下金发藓植物含水率均显著高于对照。因此, 当柳杉林密度较低时(此时, 金发藓植物群落的盖度变化不明显), 林下金发藓植物储水量可得到有效提升。该结果与贵州独山湿地保护区开展的相关研究结果一致[26]。柳杉林下金发藓植物储水能力增加可能由柳杉林的遮荫作用导致。研究表明森林遮荫一方面可以降低阳光对林下植物的直接照射, 降低林下植物的蒸腾作用, 另一方面可以保持土壤潮湿、维持较稳定的林下空气潮湿度, 为林下植物提供水源补给[37]。柳杉林的遮荫作用可通过本研究中表层土壤含水率的变化得到部分印证:雨季, 没有柳杉种植的对照区表层土壤含水率最高, 但到干季这个区域的表层土壤含水率显著降低;但是在柳杉种植区, 雨季和干季的表层土壤含水率均维持在较稳定的水平。此外, 本研究中柳杉种植区的金发藓含水率、表层土壤含水率、金发藓含水量、表层土壤含水量和系统储水量的稳定性均高于对照, 这说明柳杉种植对维持金发藓沼泽储水功能的稳定性有积极作用。另据本研究中柳杉密度与金发藓含水率、表层土壤含水率的关系可推知:干季柳杉林的遮荫作用是维持金发藓沼泽储水功能稳定的重要因素。
需要注意的是, 当柳杉密度很高时, 由于林下金发藓植物群落盖度很低, 金发藓植物的储水量几乎可以忽略不计。此时, 柳杉凋落物成为地表储水的主要物质基础。但研究表明, 贵州境内柳杉枯(凋)落物的持水性能较差[38]。综上, 人工柳杉林对林下金发藓沼泽储水功能的影响是非线性的, 呈现明显的季节差异, 因此, 本研究的第二个假设只适用于雨季柳杉林密度较高时, 即:雨季高密度的人工柳杉林可降低林下金发藓沼泽的储水能力。
本研究认为有必要对贵州娘娘山顶的人工柳杉林密度进行控制。经过合理间伐后, 柳杉林下金发藓沼泽的储水功能可以得到提升;同时, 间伐后经过一段时间的恢复, 可以消减柳杉种植对金发藓植物群落结构造成的负面影响。但是, 柳杉林的间伐方式和间伐强度还需要开展进一步的研究论证。
致谢: 感谢六盘水娘娘山国家湿地公园管理处王凯、郭应和李龙江在野外调查中给予的帮助。| [1] |
王世杰, 张信宝, 白晓永. 中国南方喀斯特地貌分区纲要. 山地学报, 2015, 33(6): 641-648. |
| [2] |
万军. 贵州省喀斯特地区土地退化与生态重建研究进展. 地球科学进展, 2003, 18(3): 447-453. DOI:10.3321/j.issn:1001-8166.2003.03.019 |
| [3] |
毛雨景, 赵志芳, 吴文春, 王锋德. 云南省水蚀荒漠化遥感调查及成因分析. 国土资源遥感, 2013, 25(1): 123-129. |
| [4] |
张文源, 王百田. 贵州喀斯特石漠化分类分级探讨. 南京林业大学学报:自然科学版, 2015, 39(2): 148-154. |
| [5] |
凡非得, 罗俊, 王克林, 陈洪松, 张伟. 桂西北喀斯特地区生态系统服务功能重要性评价与空间分析. 生态学杂志, 2011, 30(4): 804-809. |
| [6] |
盛茂银, 熊康宁, 崔高仰, 刘洋. 贵州喀斯特石漠化地区植物多样性与土壤理化性质. 生态学报, 2015, 35(2): 434-448. |
| [7] |
郭兵, 姜琳, 罗巍, 杨光, 戈大专. 极端气候胁迫下西南喀斯特山区生态系统脆弱性遥感评价. 生态学报, 2017, 37(21): 7219-7231. |
| [8] |
凡非得, 王克林, 宣勇, 岳跃民. 西南喀斯特区域生态环境敏感性评价及其空间分布. 长江流域资源与环境, 2011, 20(11): 1394-1399. |
| [9] |
Wang S J, Liu Q M, Zhang D F. Karst rocky desertification in southwestern China:geomorphology, landuse, impact and rehabilitation. Land Degradation & Development, 2004, 15(2): 115-121. DOI:10.1002/ldr.592 |
| [10] |
熊康宁, 朱大运, 彭韬, 喻理飞, 薛建辉, 李坡. 喀斯特高原石漠化综合治理生态产业技术与示范研究. 生态学报, 2016, 36(22): 7109-7113. |
| [11] |
郭柯, 刘长成, 董鸣. 我国西南喀斯特植物生态适应性与石漠化治理. 植物生态学报, 2011, 35(10): 991-999. |
| [12] |
张军以, 戴明宏, 王腊春, 苏维词, 曹立国. 西南喀斯特石漠化治理植物选择与生态适应性. 地球与环境, 2015, 43(3): 269-278. |
| [13] |
喻阳华, 李光容, 严令斌, 黄宗胜, 喻理飞. 赤水河上游主要树种根际土壤调控水分功能研究. 水土保持研究, 2015, 22(5): 284-288. |
| [14] |
石小亮, 张颖, 单永娟, 段维娜. 云南省高原典型森林植被涵养水源功能研究. 长江流域资源与环境, 2015, 24(8): 1366-1372. DOI:10.11870/cjlyzyyhj201508015 |
| [15] |
马维玲, 石培礼, 宗宁, 赵广帅, 柴曦, 耿守保. 太行山区主要森林生态系统水源涵养能力. 中国生态农业学报, 2017, 25(4): 478-489. |
| [16] |
崔宁洁, 张丹桔, 刘洋, 张健, 欧江, 张捷, 邓超, 纪托未. 不同林龄马尾松人工林林下植物多样性与土壤理化性质. 生态学杂志, 2014, 33(10): 2610-2617. |
| [17] |
张淑兰, 张海军, 张武, 王长宝, 邓际华, 宋英博, 张强. 小兴安岭南麓典型森林类型的土壤水文功能研究. 水土保持研究, 2015, 22(1): 140-145. |
| [18] |
张娇, 刘江华, 吴孟鸿, 普学才, 潘雄. 昆明山地不同人工林类型冬季土壤水分特征研究. 西南林业大学学报, 2016, 36(1): 38-43. |
| [19] |
王顺利, 刘贤德, 金铭, 张学龙, 赵维俊, 王荣新, 马剑. 青海云杉林下苔藓层对土壤蒸发的影响. 干旱区资源与环境, 2017, 31(4): 131-135. |
| [20] |
林凯敏, 俞新妥, 黄宝龙, 何智英. 杉木人工林林下植物物种多样性的动态特征. 应用与环境生物学报, 2001, 7(1): 13-19. DOI:10.3321/j.issn:1006-687X.2001.01.003 |
| [21] |
杨超, 田大伦, 胡曰利, 闫文德, 方晰, 梁小翠. 连栽杉木林林下植被生物量动态格局. 生态学报, 2011, 31(10): 2737-2747. |
| [22] |
郎惠卿. 中国湿地植被. 北京: 科学出版社, 2000.
|
| [23] |
刘艳, 赵正武. 基于最大熵模型模拟气候变化下中国两个沼泽藓类属的潜在分布. 应用与环境生物学报, 2017, 23(5): 792-799. |
| [24] |
麻俊虎, 李大华, 彭涛, 李飞, 文浩斌, 莫秀模, 周建忠. 独山泥炭藓沼泽湿地五种泥炭藓植物持水特性研究. 分子植物育种, 2017, 15(3): 1170-1176. |
| [25] |
周传艳, 陈训, 刘晓玲, 赵卫权, 李坤, 唐金刚. 基于土地利用的喀斯特地区生态系统服务功能价值评估——以贵州省为例. 应用与环境生物学报, 2011, 17(2): 174-179. |
| [26] |
杨成华, 潘平晓, 莫秀模, 钱爽霏. 贵州省独山湿地保护区两个主要沼泽植被类型地被层持水量研究. 中央民族大学学报:自然科学版, 2014, 23(1): 22-26. |
| [27] |
崔海军, 张勇, 张银峰, 田昆, 肖德荣, 王凯, 郭应, 李龙江. 贵州娘娘山湿地藓类沼泽植物群落特征及优势种种间关系. 生态学杂志, 2018, 37(9): 2619-2626. |
| [28] |
李博, 杨持. 生态学. 北京: 高等教育出版社, 2000.
|
| [29] |
聂朝俊, 张克丽, 邓朝国, 杨怡之. 贵州几个树种的苗木抗冻性研究. 贵州林业科技, 2013, 41(1): 1-4. |
| [30] |
郭水良, 曹同. 长白山主要生态系统地面藓类植物分布格局研究. 应用生态学报, 1999, 10(3): 270-274. DOI:10.3321/j.issn:1001-9332.1999.03.004 |
| [31] |
郭水良, 曹同. 长白山主要生态系统地面藓类植物的生态位研究. 生态学报, 2001, 21(2): 231-236. DOI:10.3321/j.issn:1000-0933.2001.02.009 |
| [32] |
苏维词, 朱文孝. 贵州喀斯特山区生态环境脆弱性分析. 山地学报, 2000, 18(5): 429-434. DOI:10.3969/j.issn.1008-2786.2000.05.005 |
| [33] |
王瑞, 安裕伦, 王培彬, 马良瑞. 贵州省生物多样性热点地区研究. 水土保持研究, 2014, 21(6): 152-157. |
| [34] |
Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature, 2006, 441(7093): 629-632. DOI:10.1038/nature04742 |
| [35] |
陈浒, 熊康宁, 刘忠斌, 邱杰, 陈永毕, 肖时珍, 杨洪. 基于生物多样性保护的河头喀斯特土地整理探索. 地球与环境, 2011, 39(4): 450-455. |
| [36] |
丁治国, 蒲恒浒, 王松, 邹军, 王才军. 贵州柳杉云毛虫发生特点与防治. 贵州林业科技, 2015, 43(2): 35-37, 42-42. |
| [37] |
黄奕龙, 傅伯杰, 陈利顶. 生态水文过程研究进展. 生态学报, 2003, 23(3): 580-587. DOI:10.3321/j.issn:1000-0933.2003.03.022 |
| [38] |
潘明亮, 丁访军, 谭伟, 戴全厚, 朱军. 六盘水地区4种典型森林类型凋落物的水文特性. 贵州农业科学, 2011, 39(5): 183-186, 190-190. DOI:10.3969/j.issn.1001-3601.2011.05.053 |