 2019, Vol. 39
2019, Vol. 39文章信息
- 邹顺, 周国逸, 张倩媚, 孟泽, 褚国伟, 徐姗, 夏艳菊
- ZOU Shun, ZHOU Guoyi, ZHANG Qianmei, MENG Ze, CHU Guowei, XU Shan, XIA Yanju
- 1992-2015年鼎湖山季风常绿阔叶林群落种间关联动态
- Long-term (1992-2015) dynamics of interspecific associations among tree species in a monsoon evergreen broad-leaved forest in Dinghushan Biosphere Reserve
- 生态学报. 2019, 39(17): 6362-6371
- Acta Ecologica Sinica. 2019, 39(17): 6362-6371
- http://dx.doi.org/10.5846/stxb201804030753
-
文章历史
- 收稿日期: 2018-04-03
- 网络出版日期: 2019-05-31






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
群落是一个有机的组合体, 群落内的物种通过相互作用形成复杂的种间关系[1-2]。群落的种间关系能够反映群落的稳定性[3], 也是决定群落结构动态变化的主要内部因素之一[4-6]。研究发现, 随着群落由稳定性低的先锋群落演替为稳定性高的顶极群落, 群落物种间的正关联关系加强[4, 6-9], 在植被的恢复过程中也有相似的现象[10-12]。相反, 在生物入侵、生态系统退化时, 群落物种间的正关联关系减弱, 负关联关系加强[3, 13-16]。此外, 处于稳定环境条件下的群落物种间的正关联关系更强, 处于环境条件不断变化下的群落或经常遭受较强干扰的群落物种间的正关联关系弱, 负关联关系更强[17-19]。
近几十年来, 气候条件变化, 如地表平均温度升高, 区域性的降水减少, 以及伴随的极端气候事件、虫害、火灾等频率的增大, 改变了北方森林、温带森林和热带森林等全球不同类型森林群落的原有结构[20-21]。许多研究分别从森林生态系统的物种多样性、径级结构与年龄组成、林分密度、植被生产力与生物量等方面分析了群落结构的变化[22-26]。但种间关系作为森林群落最重要的结构和数量特征之一[27-28], 对其动态变化的报道较少。
季风常绿阔叶林是我国南亚热带的地带性植被。已有研究表明, 由于气温升高和降水变率增大, 季风常绿阔叶林土壤水分含量在持续下降[29-30]。季风常绿阔叶林生物量、林分密度、径级结构、物种组成等群落结构特征发生了显著变化[22, 31]。但这些变化对季风常绿阔叶林的种间关系产生了怎样的影响还不清楚, 利用固定样地长期监测季风常绿阔叶林种间关系动态变化的研究尚未见报道。因此, 本文利用1992—2015年, 鼎湖山季风常绿阔叶林1 hm2监测样地7次群落调查数据, 采用方差比率和Spearman秩相关分析的方法, 主要探讨3个科学问题: (1)鼎湖山季风常绿阔叶林群落整体关联性怎样变化? (2)群落各垂直层次内和层次之间的种间关系如何变化? (3)哪些物种是群落种间关系中的关键物种, 它们与其他物种的种间联结有何变化?本文旨在揭示季风常绿阔叶林群落种间关系的长期动态变化, 为季风常绿阔叶林的健康评估和科学管理提供理论依据。
1 材料和方法 1.1 研究地区与样地概况鼎湖山国家级自然保护区(23.16°—23.19° N, 112.51°—112.56° E)位于广东省肇庆市鼎湖区, 占地面积约1155 hm2, 地形主要为丘陵和低山, 最高峰鸡笼山海拔1000.3 m[32]。该地区气候属南亚热带季风气候, 年平均降水量为1714 mm, 年平均湿度为76%, 干湿季分明, 湿季(4—9月)降水量约占80%。年平均气温为22.5 ℃, 最冷月(1月)平均气温为13.8 ℃, 最热月(7月)平均气温为28.8 ℃。土壤类型为赤红壤, 土层厚度30—90 cm[22], 该地区土壤自然酸化严重, 水提pH约为3.7。
鼎湖山季风常绿阔叶林(以下简称季风林, MEBF)主要分布于保护区核心区三宝峰, 海拔100—490 m, 林龄在400年以上, 处于演替过程的近顶极阶段[32]。该群落终年常绿, 郁闭度约95%, 其中, 乔木层郁闭度约80%, 可分为2个亚层:第一亚层优势种为锥(Castanopsis chinensis)、木荷(Schima superba)、黄果厚壳桂(Cryptocarya concinna)、厚壳桂(Cryptocarya chinensis)、白颜树(Gironniera subaequalis)等, 第二亚层主要是鼎湖血桐(Macaranga sampsonii)、香楠(Aidia canthioides)等;灌木层盖度约50%, 优势种为柏拉木(Blastus cochinchinensis);草本层盖度约40%, 优势种为华山姜(Alpinia oblongifolia)等。
1.2 样地设置与调查鼎湖山季风林监测样地处于低山中坡, 坡度25—35°。样地投影面积为10000 m2, 采用全站仪将样地划分为25个20 m × 20 m的样方, 每个20 m × 20 m样方用插值法细分为4个10 m × 10 m样方, 10 m × 10 m样方共100个。1992—2015年共进行了7次群落调查, 调查时记录样地内所有胸径(DBH)大于等于1.0 cm个体的物种名、胸径、高度、坐标及生长状况等信息[33]。本文利用1992—2015年7次群落调查数据, 以10 m × 10 m样方为研究单元, 分析群落主要物种间的种间关系[34]。
1.3 重要值与群落主要物种采用重要值(Important value)((相对多度+相对显著度+相对频度) / 3)筛选群落中的主要物种[35], 其中显著度为生物量[36]。为了更加全面的识别群落中的主要物种, 只要某物种在7次群落调查中出现过至少1次物种重要值≥ 1.0, 则我们认为它是季风林中的主要物种。鼎湖山季风林有29种主要物种(表 1), 2015年其重要值、个体数和生物量之和分别占群落的90.4%、94.2%和97.0%。
| 物种 Species |
代码 Code |
1992 | 1994 | 1999 | 2004 | 2008 | 2010 | 2015 |
| 生活型Ⅰ Life form Ⅰ | ||||||||
| 锥Castanopsis chinensis | Cch | 9.3 | 9.2 | 8.5 | 9.7 | 9.0 | 8.6 | 7.9 |
| 白颜树Gironniera subaequalis | Gs | 3.2 | 3.3 | 3.8 | 4.4 | 4.8 | 5.1 | 5.9 |
| 肖蒲桃Syzygium acuminatissimum | Sa | 4.7 | 4.8 | 4.7 | 5.1 | 4.8 | 4.9 | 5.1 |
| 木荷Schima superba | Ss | 4.6 | 4.6 | 4.8 | 5.6 | 4.7 | 4.1 | 2.9 |
| 窄叶半枫荷Pterospermum lanceifolium | Pl | 1.5 | 1.6 | 1.7 | 2.1 | 2.3 | 2.3 | 2.8 |
| 厚壳桂Cryptocarya chinensis | Cr | 2.5 | 2.3 | 2.4 | 2.3 | 2.9 | 2.1 | 2.2 |
| 橄榄Canarium album | Ca | 1.0 | 1.1 | 1.4 | 1.7 | 1.8 | 1.8 | 2.0 |
| 白楸Mallotus paniculatus | Ma | 0.2 | 0.2 | 0.4 | 0.7 | 2.1 | 2.1 | 2.0 |
| 黄果厚壳桂Cryptocarya concinna | Cc | 10.2 | 9.9 | 8.4 | 1.0 | 1.5 | 1.6 | 1.8 |
| 臀果木Pygeum topengii | Pt | 1.7 | 1.7 | 1.5 | 1.5 | 1.7 | 1.7 | 1.7 |
| 肉实树Sarcosperma laurinum | Sl | 1.9 | 1.9 | 1.9 | 1.9 | 1.7 | 1.7 | 1.5 |
| 观光木Michelia odora | Mo | 0.9 | 0.9 | 1.0 | 1.1 | 1.1 | 1.2 | 1.2 |
| 华润楠Machilus chinensis | Mc | 0.9 | 0.9 | 1.0 | 1.2 | 1.1 | 1.1 | 1.2 |
| 鹅掌柴Schefflera heptaphylla | Sh | 0.9 | 0.9 | 0.8 | 1.1 | 1.1 | 1.2 | 1.2 |
| 黄杞Engelhardia roxburghiana | Er | 1.4 | 1.4 | 1.7 | 1.7 | 0.8 | 0.9 | 1.0 |
| 笔罗子Meliosma rigida | Mr | 1.2 | 1.2 | 1.1 | 1.0 | 0.9 | 1.0 | 0.6 |
| 广东金叶子Craibiodendron scleranthum | Cs | 1.0 | 1.1 | 0.9 | 0.9 | 0.7 | 0.8 | 0.6 |
| 生活型Ⅱ Life form Ⅱ | ||||||||
| 香楠Aidia canthioides | Ac | 2.6 | 2.7 | 3.3 | 5.1 | 8.4 | 9.5 | 11.0 |
| 鼎湖血桐Macaranga sampsonii | Ms | 1.4 | 1.4 | 2.8 | 4.6 | 7.7 | 7.7 | 8.3 |
| 柏拉木Blastus cochinchinensis | Bc | 5.5 | 5.4 | 5.0 | 5.1 | 5.4 | 6.1 | 8.1 |
| 云南银柴Aporosa yunnanensis | Ay | 15.1 | 15.3 | 15.1 | 13.6 | 8.5 | 8.1 | 7.2 |
| 褐叶柄果木Mischocarpus pentapetalus | Mi | 1.8 | 1.8 | 1.5 | 2.7 | 3.3 | 3.4 | 3.5 |
| 红枝蒲桃Syzygium rehderianum | Sr | 3.3 | 3.4 | 3.1 | 3.1 | 2.7 | 2.6 | 2.7 |
| 光叶红豆Ormosia glaberrima | Og | 2.5 | 2.5 | 2.6 | 2.4 | 2.1 | 1.9 | 2.1 |
| 黄叶树Xanthophyllum hainanense | Xh | 1.7 | 1.8 | 2.0 | 2.0 | 2.0 | 2.0 | 2.1 |
| 罗伞树Ardisia quinquegona | Aq | 2.5 | 2.4 | 2.3 | 1.8 | 1.5 | 1.5 | 1.6 |
| 黄毛榕Ficus esquiroliana | Fe | 0.2 | 0.2 | 0.4 | 1.1 | 1.2 | 1.2 | 1.5 |
| 鼎湖钓樟Lindera chunii | Lc | 1.6 | 1.4 | 1.1 | 1.2 | 0.8 | 0.7 | 0.5 |
| 九节Psychotria asiatica | Pa | 3.7 | 3.7 | 3.7 | 3.1 | 1.7 | 1.5 | 0.2 |
| 其他Others | 11.0 | 11.0 | 11.1 | 11.2 | 11.7 | 11.6 | 9.6 | |
| 合计Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |
基于生活型分类系统把群落物种划分为2类:生活型Ⅰ, 为中乔木-大乔木(Middle and big arbor)植物, 在季风林内成熟个体树高大于等于8 m, 占据冠层位置;生活型Ⅱ, 为小乔木-灌木(Small arbor and shrub)植物, 在季风林内成熟个体树高小于8 m[37]。
1.5 总体关联性检验采用Schluter[38]提出的方差比率法(VR)检验群落多物种间的总体关联性。
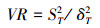
|
式中, 总样方物种数方差
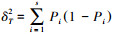
在独立零假设下, VR期望值为1, 表示群落物种间整体无关联, VR>1表示整体表现正联结, VR < 1表示整体表现负联结。采用统计量ω(ω=VR×N)检验VR偏离1的显著性, 当χ0.952(N)<ω<χ0.052(N)时, 认为整体关联不显著[27, 38]。
根据简单线性模型拟合, 得到VR值随时间的变化趋势, 以反映群落总体关联性的变化趋势。
1.6 物种种间关联分析基于个体数的Spearman秩相关分析不但能够得到种对间的联结方式(正或负联结)及显著性, 其计算公式如下:
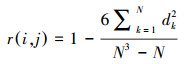
|
式中, r(i, j)为Spearman秩相关系数, N为总样方数;dk=(xik -xjk), 而xik和xjk分别为种i和种j在样方k中的秩[27]。
进一步地, 通过简单线性模型拟合得到各类型种对数随时间的变化趋势, 以反映群落种间关系的变化趋势。
2 结果 2.1 群落整体方差比率法及其变化由表 2可知, 从1992年到2015年, 鼎湖山季风林群落VR皆显著(P<0.05)大于1, 群落总体呈显著正联结关系;然而, 1992年群落VR为2.18, 到2015年减小为1.40, VR总体呈显著下降趋势(R2=0.93, P<0.001), 群落正联结关系减弱。
| 年 Year |
方差比率 Variance ratio |
检验统计量 Statistic ω |
χ2临界值 χ2 threshold value |
结果 Result |
| 1992 | 2.18 | 217.84 | [77.93, 124.34] | 显著正联结 |
| 1994 | 2.20 | 220.32 | [77.93, 124.34] | 显著正联结 |
| 1999 | 2.07 | 207.41 | [77.93, 124.34] | 显著正联结 |
| 2004 | 1.98 | 197.50 | [77.93, 124.34] | 显著正联结 |
| 2008 | 1.68 | 167.84 | [77.93, 124.34] | 显著正联结 |
| 2010 | 1.66 | 165.84 | [77.93, 124.34] | 显著正联结 |
| 2015 | 1.40 | 139.95 | [77.93, 124.34] | 显著正联结 |
| 方差比率变化趋势The trend of variance ratio 显著下降Significantly decreased(R2=0.93, P<0.001) | ||||
由表 3可知, 1992—2015年, 鼎湖山季风林29个主要物种显著正联结种对数由1992年的68对减少为2015年的50对, 总体呈显著下降趋势(R2=0.82, P=0.02), 显著负联结种对数由1992年的20对增加为2015年的25对, 总体呈显著上升趋势(R2=0.69, P=0.01)。
| 年Year | 正相关Positive correlation | 负相关Negative correlation | 不相关 No correlation |
总计 Sum. |
|||
| 显著 Significant (P<0.05) |
不显著 Not significant (P ≥ 0.05) |
显著 Significant (P<0.05) |
不显著 Not significant (P ≥ 0.05) |
||||
| 1992 | 68 | 163 | 20 | 153 | 2 | 406 | |
| 1994 | 73 | 156 | 18 | 159 | 0 | 406 | |
| 1999 | 62 | 159 | 19 | 166 | 0 | 406 | |
| 2004 | 55 | 167 | 24 | 159 | 1 | 406 | |
| 2008 | 49 | 157 | 27 | 173 | 0 | 406 | |
| 2010 | 54 | 163 | 26 | 163 | 0 | 406 | |
| 2015 | 50 | 173 | 25 | 157 | 1 | 406 | |
| 趋势Trends | SD | NS | SI | NS | NS | - | |
| SD:显著下降(P<0.05); NS:不显著(P>0.05); SI:显著上升(P<0.05) | |||||||
由表 4可知, 1992—2015年, 生活型Ⅰ物种之间显著正联结种对数总体变化不显著(P ≥ 0.05), 但1992年(9对)、1994年(12对)显著正联结种对数要明显高于1999—2015年对数(4.4对);生活型Ⅱ物种之间显著正联结种对数由1992年的28对减少为2015年的20对, 总体呈显著下降趋势(R2=0.59, P=0.02), 而显著负联结种对数总体变化不显著(P ≥ 0.05), 但1992—1999年显著负联结种对数(5.7对)要明显低于1999—2015年对数(11.2对);生活型Ⅰ与生活型Ⅱ物种组成的种对中, 显著正联结种对数由1992年的31对减少为2015年的25对, 总体呈显著下降趋势(R2=0.52, P=0.04)。
| 年Year | 正相关Positive correlation | 负相关Negative correlation | 不相关 No correlation |
总计 Sum. |
|||
| 显著 Significant (P<0.05) |
不显著 Not significant (P ≥ 0.05) |
显著 Significant (P<0.05) |
不显著 Not significant (P ≥ 0.05) |
||||
| 种对类型1:生活型Ⅰ-生活型Ⅰ Type 1 species pairs: life form Ⅰ-life form Ⅰ | |||||||
| 1992 | 9 | 60 | 3 | 63 | 1 | 136 | |
| 1994 | 12 | 56 | 3 | 65 | 0 | 136 | |
| 1999 | 4 | 58 | 3 | 71 | 0 | 136 | |
| 2004 | 4 | 57 | 1 | 74 | 0 | 136 | |
| 2008 | 4 | 57 | 2 | 73 | 0 | 136 | |
| 2010 | 5 | 61 | 3 | 67 | 0 | 136 | |
| 2015 | 5 | 67 | 2 | 61 | 1 | 136 | |
| 趋势Trends | NS | NS | NS | NS | NS | - | |
| 种对类型2:生活型Ⅱ-生活型Ⅱ Type 2 species pairs: life form Ⅱ-life form Ⅱ | |||||||
| 1992 | 28 | 17 | 6 | 15 | 0 | 66 | |
| 1994 | 28 | 19 | 6 | 13 | 0 | 66 | |
| 1999 | 30 | 19 | 5 | 12 | 0 | 66 | |
| 2004 | 29 | 16 | 10 | 11 | 0 | 66 | |
| 2008 | 23 | 16 | 13 | 14 | 0 | 66 | |
| 2010 | 24 | 17 | 12 | 13 | 0 | 66 | |
| 2015 | 20 | 19 | 10 | 17 | 0 | 66 | |
| 趋势Trends | SD | NS | NS | NS | NS | - | |
| 种对类型3:生活型Ⅰ-生活型Ⅱ Type 3 species pairs: life form Ⅰ-life form Ⅱ | |||||||
| 1992 | 31 | 86 | 11 | 75 | 1 | 204 | |
| 1994 | 33 | 81 | 9 | 81 | 0 | 204 | |
| 1999 | 28 | 82 | 11 | 83 | 0 | 204 | |
| 2004 | 22 | 94 | 13 | 74 | 1 | 204 | |
| 2008 | 22 | 84 | 12 | 86 | 0 | 204 | |
| 2010 | 25 | 85 | 11 | 83 | 0 | 204 | |
| 2015 | 25 | 87 | 13 | 79 | 0 | 204 | |
| 趋势Trends | SD | NS | NS | NS | NS | - | |
1992年, 鼎湖山季风林群落29个主要物种中有25个物种构成了68个显著正联结种对, 其中与九节、罗伞树、红枝蒲桃、光叶红豆、香楠、肉实树、鼎湖钓樟、黄叶树、黄果厚壳桂、广东金叶子和柏拉木形成的显著正联结种对数较多, 分别有11对、11对、11对、10对、10对、10对、8对、8对、8对、7对和7对;而2015年, 29个主要物种中有26个物种构成了50个显著正联结种对, 其中与香楠、红枝蒲桃、光叶红豆、黄叶树、罗伞树和鼎湖钓樟形成的显著正联结的种对数较多, 分别有12对、10对、7对、6对、6对和6对(图 1)。

1992年, 群落29个主要物种中有16个物种构成了20个显著正联结种对, 其中与鼎湖血桐、窄叶半枫荷形成的显著负联结种对数较多, 分别有8对和7对;而2015年, 29个主要物种中有21个物种构成了25个显著负联结的种对, 其中与鼎湖血桐、窄叶半枫荷形成的显著负联结的种对数较多, 分别有11对和5对(图 2)。

研究发现, 与草地群落不同[39], 随着森林的演替, 森林群落种间关系正关联逐渐增强[4, 6-9], 这可能和森林复杂的空间结构有关[40]。当针阔混交林演替为中生性常绿阔叶林时, 群落形成复杂的种间关系, 但主要表现为群落种间正关联关系加强和负关联关系减弱, 而群落结构趋于稳定, 达到多物种稳定共存[4]。我们的研究发现, 1992—2015年间群落VR和显著正负联结种对数之比皆显著大于1, 表明鼎湖山季风林作为近演替顶极的森林群落, 群落整体呈显著正关联, 群落结构相对稳定, 与之前的研究结果一致[34, 41-42]。
但与此同时, 研究还发现从1992到2015年, 群落方差比率和显著正联结种对数显著下降, 显著负联结种对数显著上升, 表明1992—2015年间该群落虽然整体仍呈显著正关联, 但种间正关联关系在逐渐减弱, 负关联关系在逐渐增强;针对群落各垂直层次还发现, 1992—2015年间群落生活型Ⅰ树种之间、生活型Ⅱ树种之间以及生活型Ⅰ树种与生活型Ⅱ树种之间的显著正联结种对数都明显减少, 表明群落各垂直层次内和垂直层次之间种间的正关联关系减弱, 而且生活型Ⅱ树种之间的负关联关系增强。该研究结果与该森林群落演替进程中种间关系的变化趋势相反[4], 表明群落原有的稳定结构遭到了破坏[17]。已有研究指出, 过去30年间, 由于气温升高和降水变率增大降低了华南地区森林土壤含水量[29-30], 由于不同径级、生活型和物种个体对逐渐干热环境的适应性不同, 鼎湖山季风林群落大径级个体和不抗旱植物个体数量减少, 小径级个体和抗旱或速生植物个体数量增加[22, 31];由于降水变率增加, 使害虫樟翠尺蛾(Thalassodes quadraia)的种群爆发频率和危害加大[43-44], 对作为季风林建群种的黄果厚壳桂种群造成了极大破坏[44];气候条件变化和自然干扰加大还可能加速了群落演替过程中残留的大个体的锥和木荷的死亡[45]。这些变化会严重地改变季风林的群落结构和种间关系, 因此1992—2015年间鼎湖山季风林群落种间关系趋势性变化可能和气候条件变化、虫害、演替以及它们之间的相互作用的影响有关。
在1992—2015年, 香楠、红枝蒲桃、光叶红豆、黄叶树、罗伞树和鼎湖钓樟与其他物种形成的显著正联结种对数较多, 很少与其他物种形成显著负联结, 暗示着这些物种能够和大多数物种稳定共存, 在季风林中有重要地位和作用, 可以作为生态恢复、林业种植的备选物种[11-12, 46]。而鼎湖血桐和窄叶半枫荷与许多物种形成显著负联结种对, 表明这两个物种与其他物种空间分布分离, 在群落中占据特殊生境或与其他物种具有很强的竞争关系。鼎湖血桐、白楸、和黄毛榕为比较典型的喜阳植物[35, 47-48], 多生于林窗、林缘等光照充足的地方[45, 49], 彼此之间形成显著正联结, 而多与其他物种形成显著负联结, 这可能是由于过去大量冠层个体死亡, 这3个物种快速占据了林窗[49], 在郁闭的季风林中, 这3个物种种群的大小可以用来衡量群落受到干扰的程度。
群落的正关联性越强, 尤其是冠层树种与下层树种的正关联性越强, 群落物种间的互补性越强, 群落能够更加充分的利用光等资源[39], 相反, 群落的负关联性越强, 物种间的互补性越弱, 不利于资源的充分利用。此外, 群落的种间关联特征与物种入侵有重要关系, 物种入侵会降低群落的正关联性, 增大负关联性[3, 13, 15], 而正关联性强的群落物种间的联结关系紧密, 群落结构稳定, 不容易遭受物种入侵[50]。鼎湖山季风林群落整体, 以及各垂直层次内、垂直层次之间种间的正关联关系减弱, 可能引起物种间的互补性不足, 对资源的利用效率下降, 进而降低群落生产力;而物种间正联结关系的减弱, 使鼎湖山季风林群落结构愈加松散, 群落稳定性下降, 遭受生物入侵的风险增大。
4 结论对鼎湖山季风常绿阔叶林种间关系动态研究表明, 从1992到2015年, 群落整体仍呈显著正关联, 但种间正关联关系在逐渐减弱, 负关联关系在逐渐增强;群落各垂直层次内和垂直层次之间种间的正关联关系在减弱, 而且生活型Ⅱ树种之间的负关联关系在增强;香楠、红枝蒲桃、光叶红豆、黄叶树、罗伞树和鼎湖钓樟能够和大多数物种长时间稳定共存, 在季风林中有重要地位和作用, 而喜阳速生的鼎湖血桐种群快速增长, 与许多物种形成显著负联结, 其种群大小可能反映了群落受到的干扰程度。鼎湖山季风林群落种间关系趋势性变化可能和气候条件变化、虫害、演替以及这些因素的相互作用的影响有关。群落正关联关系减弱, 可能造成群落资源利用效率下降, 降低群落生产力;群落物种结构愈加松散, 群落稳定性下降, 增大遭受生物入侵的风险。
| [1] |
叶万辉. 物种多样性与植物群落的维持机制. 生物多样性, 2000, 8(1): 17-24. DOI:10.3321/j.issn:1005-0094.2000.01.003 |
| [2] |
刘翔宇, 何东, 田文斌, 宋彦君, 尹芳, 许洺山, 程浚洋, 阎恩荣. 浙江普陀山岛森林木本植物的种间关联格局. 植物生态学报, 2017, 41(12): 1219-1227. |
| [3] |
郭连金. 苏门白酒草对乡土植物群落种间联结性及稳定性的影响. 亚热带植物科学, 2011, 40(2): 18-23. DOI:10.3969/j.issn.1009-7791.2011.02.006 |
| [4] |
周先叶, 王伯荪, 李鸣光, 昝启杰. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析. 植物生态学报, 2000, 24(3): 332-339. DOI:10.3321/j.issn:1005-264X.2000.03.015 |
| [5] |
韩路, 王海珍, 王家强, 郝文芳. 塔里木荒漠绿洲过渡带植物种间关联性分析. 生态环境学报, 2015, 24(6): 932-937. |
| [6] |
Gu L, Gong Z W, Li W Z. Niches and interspecific associations of dominant populations in three changed stages of natural secondary forests on Loess Plateau, P.R. China. Scientific Reports, 2017, 7: 6604. DOI:10.1038/s41598-017-06689-9 |
| [7] |
康冰, 刘世荣, 温远光, 张跃进, 姜在民, 常建国. 广西大青山南亚热带次生林演替过程的种群动态. 植物生态学报, 2006, 30(6): 931-940. DOI:10.3321/j.issn:1005-264X.2006.06.007 |
| [8] |
张岗岗, 王得祥, 张明霞, 刘文桢, 郭小龙. 秦岭南坡松栎林群落演替过程中种间联结性和相关性研究. 西北植物学报, 2015, 35(8): 1657-1668. |
| [9] |
张悦, 郭利平, 易雪梅, 曹伟, 王远遐, 吴培莉, 姬兰柱. 长白山北坡3个森林群落主要树种种间联结性. 生态学报, 2015, 35(1): 106-115. |
| [10] |
王秀梅, 刘志民, 刘博, 闫守刚. 流动沙丘区和固定沙丘区丘间低地植物种间关联关系. 生态学杂志, 2010, 29(1): 16-21. |
| [11] |
Yang Q W, Liu S J, Hu C H, Lin Y B, Zhang B, Luo M L, Peng H L. Ecological species groups and interspecific association of vegetation in natural recovery process at Xiejiadian landslide after 2008 Wenchuan earthquake. Journal of Mountain Science, 2016, 13(9): 1609-1620. DOI:10.1007/s11629-016-3807-8 |
| [12] |
Liu L T, Wang X D, Wen Q, Jia Q Q, Liu Q J. Interspecific associations of plant populations in rare earth mining wasteland in southern China. International Biodeterioration & Biodegradation, 2017, 118: 82-88. |
| [13] |
郭连金, 王涛. 空心莲子草入侵对乡土植物群落种间联结性及稳定性的影响. 中国生态农业学报, 2009, 17(5): 851-856. |
| [14] |
刘珏宏, 高慧, 张丽红, 陈丽萍, 赵念席, 高玉葆. 内蒙古锡林郭勒草原大针茅-克氏针茅群落的种间关联特征分析. 植物生态学报, 2010, 34(9): 1016-1024. DOI:10.3773/j.issn.1005-264x.2010.09.002 |
| [15] |
钟军弟, 李先琨, 吕仕洪, 曾丹娟, 陈燕, 成夏岚. 桂西南喀斯特区域群落主要环境因子及稳定性对飞机草入侵的影响. 生态环境学报, 2011, 20(12): 1819-1826. DOI:10.3969/j.issn.1674-5906.2011.12.007 |
| [16] |
龚容, 高琼, 王亚林. 围封对温带半干旱典型草原群落种间关联的影响. 植物生态学报, 2016, 40(6): 554-563. |
| [17] |
贺立静, 张璐, 苏志尧. 南岭国家级自然保护区不同保护条件下优势种群的种间联结性分析. 华南农业大学学报, 2011, 32(1): 73-77. DOI:10.3969/j.issn.1001-411X.2011.01.016 |
| [18] |
简敏菲, 刘琪璟, 朱笃, 游海. 九连山常绿阔叶林乔木优势种群的种间关联性分析. 植物生态学报, 2009, 33(4): 672-680. DOI:10.3773/j.issn.1005-264x.2009.04.005 |
| [19] |
简尊吉, 李堂中, 郭泉水, 秦爱丽, 马凡强, 裴顺祥, 肖文发. 三峡水库峡谷地貌区消落带主要植物种间关联性研究. 西北植物学报, 2016, 36(10): 2086-2093. DOI:10.7606/j.issn.1000-4025.2016.10.2086 |
| [20] |
Allen C D, Macalady A K, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears D D, Hogg E H, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J H, Allard G, Running S W, Semerci A, Cobb N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 2010, 259(4): 660-684. DOI:10.1016/j.foreco.2009.09.001 |
| [21] |
Allen C D, Breshears D D, McDowell N G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere, 2015, 6(8): 1-55. |
| [22] |
Zhou G Y, Peng C H, Li Y L, Liu S Z, Zhang Q M, Tang X L, Liu J X, Yan J H, Zhang D Q, Chu G W. A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China. Global Change Biology, 2013, 19(4): 1197-1210. DOI:10.1111/gcb.12128 |
| [23] |
Zhou G Y, Houlton B Z, Wang W T, Huang W J, Xiao Y, Zhang Q M, Liu S Z, Cao M, Wang X H, Wang S L, Zhang Y P, Yan J H, Liu J X, Tang X L, Zhang D Q. Substantial reorganization of China′s tropical and subtropical forests: based on the permanent plots. Global Change Biology, 2014, 20(1): 240-250. DOI:10.1111/gcb.12385 |
| [24] |
Anderegg W R L, Hicke J A, Fisher R A, Allen C D, Aukema J, Bentz B, Hood S, Lichstein J W, Macalady A K, McDowell N, Pan Y D, Raffa K, Sala A, Shaw J D, Stephenson N L, Tague C, Zeppel M. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytologist, 2015, 208(3): 674-683. DOI:10.1111/nph.13477 |
| [25] |
Bennett A C, McDowell N G, Allen C D, Anderson-Teixeira K J. Larger trees suffer most during drought in forests worldwide. Nature Plants, 2015, 1(10): 15139. DOI:10.1038/nplants.2015.139 |
| [26] |
Anderegg W R L, Klein T, Bartlett M, Sack L, Pellegrini A F A, Choat B, Jansen S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(18): 5024-5029. DOI:10.1073/pnas.1525678113 |
| [27] |
张金屯. 数量生态学(第二版). 北京: 科学出版社, 2011.
|
| [28] |
徐满厚, 刘敏, 翟大彤, 刘彤. 植物种间联结研究内容与方法评述. 生态学报, 2016, 36(24): 8224-8233. |
| [29] |
Zhou G Y, Wei X H, Wu Y P, Liu S G, Huang Y H, Yan J H, Zhang D Q, Zhang Q M, Liu J X, Meng Z, Wang C L, Chu G W, Liu S Z, Tang X L, Liu X D. Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China. Global Change Biology, 2011, 17(12): 3736-3746. DOI:10.1111/j.1365-2486.2011.02499.x |
| [30] |
刘效东, 周国逸, 陈修治, 张德强, 张倩媚. 南亚热带森林演替过程中小气候的改变及对气候变化的响应. 生态学报, 2014, 34(10): 2755-2764. |
| [31] |
Li R H, Zhu S D, Chen H Y H, John R, Zhou G Y, Zhang D Q, Zhang Q M, Ye Q. Are functional traits a good predictor of global change impacts on tree species abundance dynamics in a subtropical forest?. Ecology Letters, 2015, 18(11): 1181-1189. DOI:10.1111/ele.12497 |
| [32] |
熊鑫, 张慧玲, 吴建平, 褚国伟, 周国逸, 张德强. 鼎湖山森林演替序列植物-土壤碳氮同位素特征. 植物生态学报, 2016, 40(6): 533-542. |
| [33] |
张咏梅, 周国逸, 温达志, 张德强, 张倩媚. 南亚热带季风常绿阔叶林锥栗-荷木-黄果厚壳桂群落发展趋势探讨. 植物生态学报, 2003, 27(2): 256-262. DOI:10.3321/j.issn:1005-264X.2003.02.017 |
| [34] |
王伯荪, 彭少麟. 鼎湖山森林群落分析——Ⅱ.物种联结性. 中山大学学报, 1983(4): 29-37. |
| [35] |
叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274-286. DOI:10.3773/j.issn.1005-264x.2008.02.005 |
| [36] |
温达志, 魏平, 孔国辉, 张倩媚, 黄忠良. 鼎湖山锥栗+黄果厚壳桂+荷木群落生物量及其特征. 生态学报, 1997, 17(5): 497-504. DOI:10.3321/j.issn:1000-0933.1997.05.008 |
| [37] |
宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001.
|
| [38] |
Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998-1005. DOI:10.2307/1938071 |
| [39] |
娄彦景, 赵魁义. 三江平原毛苔草群落近30年演替过程中的种间联结性分析. 生态学杂志, 2008, 27(4): 509-513. |
| [40] |
周刘丽, 张晴晴, 赵延涛, 许洺山, 程浚洋, 朱丹妮, 宋彦君, 黄海侠, 史青茹, 阎恩荣. 浙江天童枫香树群落不同垂直层次物种间的联结性与相关性. 植物生态学报, 2015, 39(12): 1136-1145. DOI:10.17521/cjpe.2015.0110 |
| [41] |
彭李菁. 鼎湖山气候顶极群落种间联结变化. 生态学报, 2006, 26(11): 3732-3739. DOI:10.3321/j.issn:1000-0933.2006.11.028 |
| [42] |
张倩媚, 陈北光, 周国逸. 鼎湖山主要林型优势树种种间联结性的计算方法研究. 华南农业大学学报, 2006, 27(1): 79-83. DOI:10.3969/j.issn.1001-411X.2006.01.021 |
| [43] |
黄忠良. 樟翠尺蛾种群动态与植物群落结构及气候因子的关系. 生态学杂志, 2000, 19(3): 24-27, 31-31. DOI:10.3321/j.issn:1000-4890.2000.03.006 |
| [44] |
陈志成, 万贤崇. 虫害叶损失造成的树木非结构性碳减少与树木生长、死亡的关系研究进展. 植物生态学报, 2016, 40(9): 958-968. |
| [45] |
张池, 黄忠良, 史军辉, 李林, 魏识广, 李炯. 鼎湖山季风常绿阔叶林木本植物个体死亡动态. 生态学报, 2006, 26(8): 2457-2462. DOI:10.3321/j.issn:1000-0933.2006.08.006 |
| [46] |
Zhao H Y, Kang X G, Guo Z Q, Yang H, Xu M. Species interactions in spruce-fir mixed stands and implications for enrichment planting in the Changbai Mountains, China. Mountain Research and Development, 2012, 32(2): 187-196. DOI:10.1659/MRD-JOURNAL-D-11-00125.1 |
| [47] |
Zhu S D, Song J J, Li R H, Ye Q. Plant hydraulics and photosynthesis of 34 woody species from different successional stages of subtropical forests. Plant, Cell & Environment, 2013, 36(4): 879-891. |
| [48] |
肖炜, 王峥峰, 曹洪麟, 叶万辉, 黄国敏. 鼎湖血桐种群生物学及遗传多样性初步研究. 广西植物, 2010, 30(1): 70-74. DOI:10.3969/j.issn.1000-3142.2010.01.012 |
| [49] |
彭闪江, 黄忠良, 周国逸, 周小勇, 张池, 贺握权. 鼎湖山林窗形成特征及其对幼树组成和多样性的影响. 热带亚热带植物学报, 2003, 11(3): 229-235. DOI:10.3969/j.issn.1005-3395.2003.03.006 |
| [50] |
Tilman D. The ecological consequences of changes in biodiversity: a search for general principles. Ecology, 1999, 80(5): 1455-1474. |