 2019, Vol. 39
2019, Vol. 39文章信息
- 景春雷, 李可可, 何亚奇, 王娟, 张雷, 李东来, 万冬梅
- JING Chunlei, LI Keke, HE Yaqi, WANG Juan, ZHANG Lei, LI Donglai, WAN Dongmei
- 杂色山雀双亲差异性育雏策略
- Sex differences in parental investment in the varied tit, Sittiparus varius
- 生态学报. 2019, 39(15): 5725-5729
- Acta Ecologica Sinica. 2019, 39(15): 5725-5729
- http://dx.doi.org/10.5846/stxb201809061904
-
文章历史
- 收稿日期: 2018-09-06
- 修订日期: 2018-12-28

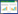




亲代抚育被定义为是指亲鸟对子代的保护、照顾、喂养等一切有利于子代存活的行为, 是一种复杂的社会行为。其主要好处是可提高后代的存活机会, 但亲代抚育需要花费大量时间和精力, 因此这种行为也给亲代带来了代价, 即降低了其生存和进一步繁殖的机会[1]。
由于鸟类育雏投入时身体能量消耗巨大, 因此双亲合理的育雏投入分配对其生存是非常重要, 雌雄育雏策略往往不同[2]。不同鸟种的双亲育雏策略存在差异, 例如对缝叶吸蜜鸟(Notiomystis cincta)的研究发现雌性比雄性有更高的递食率[3];但是在黑枕威森莺(Wilsonia citrina)的研究中却发现雄性比雌性的递食率高[4]。在此, 我们探究一种狭域分布的社会性单配制鸟类杂色山雀(Sittiparus varius)的双亲育雏投入策略。
影响双亲育雏投入的因素有很多, 包括双亲身体质量[5]、婚外父权[6]、气候因素[7]等。其中, 身体质量是影响育雏投入的关键因素[8], 高质量的个体比低质量的个体有更多的能量储备, 所以能够以更好的状态应对不利条件, 从而提供更多的育雏投入。传统生态学认为“身体质量”是多个表型性状的产物, 通常采用体征参数表示[9], 将鸟类的喙宽、喙长、翅长、跗跖长、尾长等体征参数作为身体大小的指标因子[10-11], 个体越大表示身体质量越高[9]。对于育雏投入与身体质量的研究结果不尽相同, 对大山雀(Parus major)的研究中发现亲鸟身体质量与其繁殖投入呈正相关关系[12], 雄性亲鸟饰羽斑块尺寸越大, 育雏投入越高[8]。但对石雀(Petronia petronia)的研究却发现拥有较高身体质量的雄鸟, 其配偶反而有更高的育雏投入[13]。大多数关于身体质量影响育雏投入的研究是从雄性亲本的角度出发, 很少有研究关注雌性亲本特征与亲代抚育的关系[14]。杂色山雀作为一种狭域分布的雀形目鸟类, 本实验旨在跨性别地探究杂色山雀双亲身体质量与育雏投入的关系。
1 材料与方法 1.1 研究地及物种概况辽宁仙人洞国家级自然保护区位于辽东半岛的东北部, 隶属于辽宁省大连庄河市仙人洞镇, 地理坐标122°53′24″—123°03′30″E, 39°54′00″—40°03′00″N, 海拔200—600 m。保护区属暖温带大陆性季风气候, 四季分明, 具有海洋性气候的特点, 雨热同季, 降水和光照集中在夏季。保护区内植物资源非常丰富, 为杂色山雀提供了丰富的食物和巢材资源[15]。杂色山雀(Sittiparus varius)隶属于雀形目山雀科山雀属, 是一种小体型的次生洞巢鸟类, 社会性单配制, 双亲抚育后代[16]。
1.2 野外工作2017年3月中旬开始, 利用鸣声招引方法, 雾网捕捉杂色山雀, 环志、翅下肱静脉取血(30 μL), 使用游标卡尺(King Guard DP—5, 150 mm/0.01 mm)测量喙长、喙宽、翅长、尾长等体征参数, 使用电子秤(Changxie, 100 g/0.01 g)测量体重。人工巢箱法招引杂色山雀繁殖, 记录雏鸟孵化时间, 将雏鸟出生日期定义为1日龄。在雏鸟6日龄时, 使用巢箱捕鸟器捕捉杂色山雀双亲, 对未曾环志的雌雄亲鸟进行取血环志与体征参数测量, 测量标准与网捕时一致。
1.3 录像及数据收集将雏鸟4—6日龄定义为育雏前期, 10—12日龄定义为育雏后期, 分育雏前期和育雏后期两个阶段进行双亲育雏投入监测。杂色山雀育雏高峰期为育雏后期, 故本实验选择育雏后期用来研究体征对其育雏投入的影响。使用小蚁运动相机(YI, YDXJ01XY)进行育雏录像。制作相机支架, 将运动相机挂在人工巢箱口附近, 保证能录到亲鸟口中叼的食物及亲鸟彩色脚环, 整个录像过程都用移动电源(MI, PLM06ZM)进行供电。为了减少人为影响, 相机架设30分钟后正式开始实验记录, 实验的有效时长3 h, 录像时间为7:00—10:00, 为了减小运动相机对亲鸟的影响, 实验过程中关闭相机声音及光源。使用potplayer播放器对视频回看分析, 通过彩色脚环区分雌雄亲鸟, 分别记录雌雄亲鸟递食次数以及总递食次数, 计算递食率(次/小时)。
1.4 数据处理利用统计软件SPSS 21.0对数据统计分析。采用配对样本t检验(paired-samples T test)对育雏前期和育雏后期雌雄递食率分别进行统计分析。考虑到雏数的影响, 经过独立样本t检验(independent-samples T test)分析发现育雏前期雏数和育雏后期雏数并没有显著性差异, 然后采用独立样本t检验(independent-samples T test)分析双亲育雏前期和育雏后期各自递食率以及总递食率差异。显著性水平(P)设置为0.05。数据以平均值±标准差(Mean ± SD)形式表示。将雏数作为协变量, 使用多元线性回归模型(multivariable linear regression model)分析双亲体征参数对育雏后期雌雄递食率的影响[14]。
1.5 室内实验用DNA提取试剂盒(Cat.#DP304—03, TIANGN公司)提取DNA, 选择P2/P8引物对杂色山雀进行性别鉴定[17]。
2 研究结果 2.1 杂色山雀的育雏期投入共对17巢育雏前期和33巢育雏后期雌雄亲鸟递食率进行了统计。在育雏前期, 雌雄亲鸟递食率无显著差异(t=1.481, df=17, P=0.157);在育雏后期, 雌性亲鸟递食率显著高于雄性递食率(t=3.024, df=32, P=0.005)。从育雏前期到育雏后期, 雌性递食率显著性增加(t=2.419, df=48, P=0.019), 而雄性递食率无显著性变化(t=0.938, df=54, P=0.532), 总递食率显著性增加(t=2.328, df=54, P=0.024)(图 1)。

|
| 图 1 育雏前期和育雏后期雌雄鸟递食率及总递食率 Fig. 1 Feeding rates of Sittiparus varius parents according to sex and stage of nestling period |
采用多元线性回归模型分析杂色山雀亲鸟体征对递食率的影响, 结果显示, 雌性亲鸟递食率与雌性的喙宽呈极显著正相关关系(β=0.849, P=0.000), 雌性递食率与雄性各个体征参数均不相关;而雄性递食率与双亲体征参数均不相关(表 1)。
| 体征参数 Physical parameters |
递食率Feeding frequency | ||||
| 雌鸟Females | 雄鸟Males | ||||
| β | P | β | P | ||
| 雌性体重Body weight/g | 0.257 | 0.539 | 0.088 | 0.803 | |
| 雌性喙长Beak length/mm | -0.244 | 0.557 | -0.405 | 0.257 | |
| 雌性喙宽Beak width/mm | 0.849 | 0.000* | 0.162 | 0.315 | |
| 雌性翅长Wing length/mm | 0.008 | 0.876 | -0.016 | 0.717 | |
| 雌性尾长Tail length/mm | 0.08 | 0.656 | 0.067 | 0.661 | |
| 雄性体重Body weight/g | 0.002 | 0.72 | 0.482 | 0.355 | |
| 雄性喙长Beak length/mm | 0.481 | 0.998 | 0.404 | 0.218 | |
| 雄性喙宽Beak width/mm | 0.277 | 0.186 | -0.063 | 0.656 | |
| 雄性翅长Wing length/mm | 0.028 | 0.087 | 0.154 | 0.697 | |
| 雄性尾长Tail length/mm | -0.029 | 0.516 | 0.015 | 0.455 | |
| β为回归系数; *表示P < 0.01, 差异极显著 | |||||
本研究结果显示, 育雏前期雌雄亲鸟递食率差异不显著;育雏后期雄性杂色山雀递食率仅略有增加, 基本与前期保持一致, 且其育雏投入与其自身体征参数、配偶体征参数等均无相关性;而雌性杂色山雀后期递食率增加明显, 远大于同期雄性配偶, 也大于其自身的前期递食率。双亲递食率差异可能是双亲投入分工不同造成的。研究发现, 雌雄双亲在抚育子代时, 常存在两性分工, 不同鸟种的双亲投入分工存在差异。对于大多数鸟类来说, 雌鸟参与孵卵、暖雏、递食等, 而雄鸟负责递食和巢防卫等[18];但是在对红隼(Falco tinnunculus)的研究发现, 在育雏期雄性主要任务是捕食, 而雌性主要进行巢防卫[19]。在育雏期, 杂色山雀雌鸟参与暖雏、递食, 而雄鸟负责递食和巢防卫。在育雏前期阶段, 杂色山雀雌性递食率低于育雏后期, 这是因为雌性会将很大一部分精力用来暖雏。而在育雏后期, 雌性没有了暖雏任务, 会将更大的精力增加育雏投入来满足日益增加的雏鸟需求;杂色山雀雄鸟递食投入少, 可能是因为育雏阶段雄鸟主要负责巢防卫, 其在育雏前后期任务分工不变, 导致其始终保持相同的递食率。
较低的递食率对于雄性来说, 可使其保留更强的竞争力来获取其他效益, 比如获得更多婚外交配的机会等, 这或许会导致雄性成为更有竞争力的性别。亲代投入假说认为, 性选择会促使具有竞争力性别的个体拥有更大的体型[20]。有研究已经证明杂色山雀雄性比雌性拥有更大的体型, 雄鸟的翅长和尾长都显著长于雌鸟[21]。在对主红雀(Cardinalis cardinalis)的研究中发现, 雄性会在繁殖投入以及寻求婚外交配之间进行能量分配[22]。杂色山雀虽为社会性单配制鸟类, 但其婚外父权发生率较高, 发生婚外父权巢的比例可达40%—45%, 婚外子代的比例为14.4%—14.7%[23], 因此我们推测雄性较低的育雏投入也许跟婚外配也有关, 当然这些推测还有待于进一步的实验验证。
身体质量是影响育雏投入的关键因素[8], 亲本提供育雏投入时会权衡自身和配偶的身体质量[24]。目前关于身体质量与育雏投入的关系主要有三种假说:优秀父母假说、差异分配假说及繁殖补偿假说[25]。差异分配假说是指与高质量配偶配对的个体提供更多的亲代抚育;繁殖补偿是指与低质量配偶配对的个体提供更多的亲代抚育, 以补偿配偶较少的亲代抚育。优秀父母假说则是说亲代抚育与亲代身体质量有直接关系, 有更好身体质量的双亲能够提供更多的育雏投入[14]。蓝山雀(Cyanistes caeruleus)的研究支持差异分配假说[26], 而斑胸草雀(Taeniopygia guttata)的研究支持繁殖补偿假说[27]。在繁殖季检测了33巢杂色山雀亲代递食率与其体征参数以及与配偶体征参数之间的关系, 结果显示雌鸟的递食率与雌鸟喙宽极显著正相关。鸟类喙宽、喙长、翅长、跗跖等体征参数可以作为鸟类身体大小的指标因子[28], 体征参数可以反映鸟类的身体质量, 体征参数越好, 身体质量越高[14]。而身体质量高的个体觅食能力强、递食率高, 会给雏鸟提供更多的营养物质来促进其生长[29]。研究中发现, 杂色山雀递食时的食物运量特点与大山雀完全不同, 大山雀每次回巢时嘴里只叼一个食物, 而杂色山雀亲鸟每次回巢递食, 嘴里至少会有2—4条虫子, 有时候是满满一嘴食物, 我们推测其喙越宽大容纳食物的空间越大, 因此喙宽越大可能其递食能力越强, 因此该研究结果初步支持优秀父母假说。但是身体质量是一个综合性指标, 除了用体征参数之外, 有的研究用羽毛鲜亮程度[30], 亲本体内皮质酮水平指标[27]以及鸟类鸣声质量[31]等衡量身体质量, 单一的身体质量指标造成结果的可比性差, 因此未来将进一步结合生理状况指标和生化指标研究亲本身体质量对其育雏投入的影响。
致谢: 感谢仙人洞自然保护区管理局对野外工作支持。| [1] |
尚玉昌. 动物行为学. 北京: 北京大学出版社, 2005.
|
| [2] |
Verbeek N A M. Daily and annual time budget of the yellow-billed magpie. Auk, 1972, 89(3): 567-582. |
| [3] |
Low M, Makan T, Castro I. Food availability and offspring demand influence sex-specific patterns and repeatability of parental provisioning. Behavioral Ecology, 2012, 23(1): 25-34. |
| [4] |
Ogden L J E, Stutchbury B J M. Fledgling care and male parental effort in the Hooded Warbler (Wilsonia citrina). Canadian Journal of Zoology, 1997, 75(4): 576-581. DOI:10.1139/z97-071 |
| [5] |
Sheldon B C. Differential allocation:tests, mechanisms and implications. Trends in Ecology and Evolution, 2000, 15(10): 397-402. DOI:10.1016/S0169-5347(00)01953-4 |
| [6] |
Du B, Guan M M, Ren Q M, Chen G L. Cuckolded male ground tits increase parental care for the brood. Animal Behaviour, 2015, 110: 61-67. DOI:10.1016/j.anbehav.2015.09.023 |
| [7] |
Sicurella B, Caffi M, Caprioli M, Rubolini D, Saino N, Ambrosini R. Weather conditions, brood size and hatching order affect Common Swift Apus apus nestlings' survival and growth. Bird Study, 2015, 62(1): 64-77. DOI:10.1080/00063657.2014.989193 |
| [8] |
Pagani-Núñez E, Senar J C. Are colorful males of great tits Parus major better parents? Parental investment is a matter of quality. Acta Oecologica, 2014, 55: 23-28. DOI:10.1016/j.actao.2013.11.001 |
| [9] |
Horváthová T, Nakagawa S, Uller T. Strategic female reproductive investment in response to male attractiveness in birds. Proceedings:Biological Sciences, 2012, 279(1726): 163-170. DOI:10.1098/rspb.2011.0663 |
| [10] |
Labocha M K, Hayes J P. Morphometric indices of body condition in birds:a review. Journal of Ornithology, 2011, 153(1): 1-22. |
| [11] |
Blums P, Nichols J D, Hines J E, Lindberg M S, Mednis A. Individual quality, survival variation and patterns of phenotypic selection on body condition and timing of nesting in birds. Oecologia, 2005, 143(3): 365-376. DOI:10.1007/s00442-004-1794-x |
| [12] |
Van Noordwijk A J, de Jong G. Acquisition and allocation of resources:their influence on variation in life history tactics. The American Naturalist, 1986, 128(1): 137-142. DOI:10.1086/284547 |
| [13] |
Matessi G, Carmagnani C, Griggio M, Pilastro A. Male rock sparrows differentially allocate nest defence but not food provisioning to offspring. Behaviour, 2009, 146(2): 209-223. DOI:10.1163/156853909X410748 |
| [14] |
Laubach Z M, Wei P, Lombardo M, Murdock C, Foufopoulos J. Determinants of parental care in mountain white-crowned sparrows (Zonotrichia leucophrys oriantha). The Auk, 2015, 132(4): 893-902. DOI:10.1642/AUK-15-9.1 |
| [15] |
王鹏, 邱英杰. 辽宁仙人洞国家级自然保护区科学考察集. 北京: 中国林业出版社, 2002: 1-9.
|
| [16] |
万冬梅, 丁锋, 王爽, 金春日, 霍亚鹏. 杂色山雀的研究现状. 四川动物, 2008, 27(1): 157-160. |
| [17] |
霍雅鹏, 曹军, 张宇婷, 万冬梅. 杂色山雀性别分子鉴定方法的研究. 辽宁林业科技, 2009(1): 3-7, 13-13. DOI:10.3969/j.issn.1001-1714.2009.01.002 |
| [18] |
Deeming C. Avian incubation:behaviour, environment and evolution. Oxford: Oxford University Press, 2002.
|
| [19] |
孙寒梅.红隼繁殖期的双亲投入与繁殖成功的关系[D].长春: 东北师范大学, 2006. http://cdmd.cnki.com.cn/article/cdmd-10200-2007013456.htm
|
| [20] |
Ramírez F, Hobson K A, Wangensteen O S, Genovart M, Viscor G, Sanpera C, Jover L. A physiological marker for quantifying differential reproductive investment between the sexes in Yellow-legged gulls (Larus michahellis). Journal of Experimental Marine Biology and Ecology, 2010, 396(1): 48-52. DOI:10.1016/j.jembe.2010.09.012 |
| [21] |
赵济川.杂色山雀育雏投入的影响因素研究[D].沈阳: 辽宁大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10140-1017097920.htm
|
| [22] |
Linville S U, Breitwisch R, Schilling A J. Plumage brightness as an indicator of parental care in northern cardinals. Animal Behaviour, 1998, 55(1): 119-127. DOI:10.1006/anbe.1997.0595 |
| [23] |
马锐强, 常鹏, 万冬梅, 鞠静, 张雷, 李东来. 杂色山雀的婚外父权与配偶间的遗传相容性无关. 生态学报, 2015, 35(15): 5018-5025. |
| [24] |
Kokko H, Jennions M D. Parental investment, sexual selection and sex ratios. Journal of Evolutionary Biology, 2008, 21(4): 919-948. |
| [25] |
Maguire S E, Safran R J. Morphological and genetic predictors of parental care in the North American barn swallow Hirundo rustica erythrogaster. Journal of Avian Biology, 2010, 41(1): 74-82. DOI:10.1111/j.1600-048X.2009.04806.x |
| [26] |
Amininasab S M, Birker M, Kingma S A, Hildenbrandt H, Komdeur J. The effect of male incubation feeding on female nest attendance and reproductive performance in a socially monogamous bird. Journal of Ornithology, 2017, 158(3): 687-696. DOI:10.1007/s10336-016-1427-2 |
| [27] |
Bolund E, Schielzeth H, Forstmeier W. Compensatory investment in zebra finches:females lay larger eggs when paired to sexually unattractive males. Proceedings of the Royal Society B:Biological Sciences, 2009, 276(1657): 707-715. DOI:10.1098/rspb.2008.1251 |
| [28] |
Cooch E, Lank D B, Dzubin A, Rockwell R F, Cooke F. Body size variation in lesser snow geese:environmental plasticity in gosling growth rates. Ecology, 1991, 72(2): 503-512. DOI:10.2307/2937191 |
| [29] |
Saino N, Calza S, Moller A P. Immunocompetence of nestling barn swallows in relation to brood size and parental effort. Journal of Animal Ecology, 1997, 66(6): 827-836. DOI:10.2307/5998 |
| [30] |
Wells S J, Safran R J, Dale J. Piecing together female extra-pair mate choice:females really do prefer more ornamented males. Molecular Ecology, 2016, 25(15): 3521-3524. DOI:10.1111/mec.13720 |
| [31] |
Zollinger S A, Brumm H. Why birds sing loud songs and why they sometimes don't. Animal Behaviour, 2015, 105: 289-295. DOI:10.1016/j.anbehav.2015.03.030 |