 2019, Vol. 39
2019, Vol. 39文章信息
- 陶宝先, 张保华, 董杰, 刘晨阳
- TAO Baoxian, ZHANG Baohua, DONG Jie, LIU Chenyang
- 有机碳质量对黄河三角洲芦苇凋落物分解及其温度敏感性的影响
- Effect of organic carbon quality on the litter decomposition and temperature sensitivity of Phragmites australi in the Yellow River Delta, China
- 生态学报. 2019, 39(15): 5564-5572
- Acta Ecologica Sinica. 2019, 39(15): 5564-5572
- http://dx.doi.org/10.5846/stxb201808261828
-
文章历史
- 收稿日期: 2018-08-26
- 网络出版日期: 2019-05-16





凋落物分解是陆地生物地球化学循环过程中的重要环节[1]。凋落物分解不仅为绝大多数陆地及水生生态系统的微生物提供养分, 在全球碳循环过程中也起着重要作用[2]。研究表明, 凋落物分解受制于气候及凋落物质量[3-6], 但在不同尺度范围内, 这些因素对凋落物分解的影响仍存在差异。在全球或区域尺度上, 影响凋落物分解的主要因素是气候条件;而局部范围内则是凋落物质量[7]。有研究发现, 凋落物生态化学计量比也是影响凋落物分解的重要因素[8]。在局部范围内, 凋落物质量及生态化学计量比对凋落物分解的影响仍不确定。早期研究认为, 生态化学计量比是控制凋落物分解的主要因素[8]。其后研究发现, 有机碳质量对凋落物分解的影响远大于生态化学计量比[9], 或生态化学计量比对凋落物分解无明显作用[10]。因此, 研究生态化学计量比及有机碳质量对凋落物分解的影响, 对于深入了解凋落物分解及其影响因素具有重要意义。
固态13C核磁共振技术作为一种非破坏性研究手段, 可以研究化合物的结构特征, 已被广泛用于凋落物[11]及土壤[12-13]有机碳结构及稳定性的研究。而且, 近期研究发现, 相对于凋落物生态化学计量比, 利用13C核磁共振技术评价凋落物有机碳质量能够更充分的解释凋落物分解[11]。因此, 利用13C核磁共振技术研究凋落物有机碳质量不仅能清晰地表明不同碳官能团(碳组分)的相对比例, 而且能深入反映凋落物有机碳质量的变化及有机碳的稳定性。
凋落物呼吸是土壤呼吸的一个重要组成部分, 占土壤呼吸的比例约为10%—40%[14-19]。研究凋落物呼吸温度敏感性的变化特征及其影响因素对准确理解地区的土壤碳循环具有重要意义[20]。有研究发现, 凋落物呼吸的温度敏感性受温度[21]、水分[22]及凋落物质量[20-21]等因素的影响。而且, 随土壤温度[22]及水分[20]增加凋落物呼吸温度敏感性呈增加趋势。早期研究发现, 有机碳质量能够影响有机碳分解温度敏感性, 但有机碳质量对土壤有机碳分解温度敏感性的影响仍不确定。土壤有机碳易分解组分分解的温度敏感性可能大于[23]、小于[24-25]或等于[26]土壤有机碳难分解组分分解的温度敏感性。这表明, 有机碳质量能够影响有机碳分解的温度敏感性。凋落物是土壤有机碳的主要来源之一, 凋落物有机碳质量与土壤有机碳质量有着密切的联系[27]。然而, 凋落物有机碳质量是否影响凋落物呼吸的温度敏感性?这方面的研究仍不充分。
滨海湿地仅占全球陆地表面积的0.22%—0.34%[28], 其土壤及植物有机碳贮量却高达13—17.2 Pg[29], 在全球碳循环过程中起着重要作用。黄河三角洲河口湿地是我国重要的河口湿地之一, 它由黄河携带大量泥沙填海淤积而成, 是我国暖温带保存最完整、最年轻的湿地生态系统。本研究以黄河三角洲典型植被——芦苇(Phragmites australis)为例, 通过凋落物袋法、室内模拟实验及固态13C核磁共振技术, 研究有机碳质量对凋落物分解及凋落物呼吸温度敏感性的影响, 探讨预测凋落物分解及其温度敏感性的指标。
1 材料与方法 1.1 研究区概况研究区设置于山东省东营市中国科学院黄河三角洲滨海湿地生态试验站(37°45′50″N, 118°59′24″E)。该区属暖温带半湿润大陆性季风气候, 四季分明, 雨热同期。年均气温12.9℃, 年降水量530—630 mm, 且主要集中在5—9月, 年蒸发量为1962 mm。湿地植被的建群种和优势种有芦苇(Phragmites australis)、盐地碱蓬(Suaeda salsa)和柽柳(Tamarix chinensis)等。该区土壤类型以潮土和盐土为主, 土壤质地主要为砂质黏壤土[30-31]。
1.2 野外实验2015年10月底, 在研究区采集当年立枯芦苇植株上叶凋落物。于实验室内用去离子水洗净, 60℃烘干至恒重。取部分烘干凋落物剪至2 cm左右, 装入1 mm孔径的尼龙分解袋(长×宽=20 cm×20 cm), 每袋放入凋落物共10 g, 共设置18袋。2016年4月将上述凋落物袋平铺于研究区地表, 实验开始的第0、30、70、160、360、720天, 每次取3袋凋落物, 于实验室内用去离子水洗净, 60℃烘干至恒重, 称重, 计算凋落物的失重率。然后, 将凋落物样品粉碎、过0.1 mm筛, 测试有机碳、总氮、总磷含量及有机碳官能团相对含量。剩余样品装入自封袋保存、用于室内模拟实验。有机碳含量采用外加热-重铬酸钾氧化法测定, 总氮及总磷采用浓硫酸-H2O2消煮法制备待测液, 总氮含量采用靛酚蓝比色法, 总磷含量采用钼锑抗比色法测定[32]。
1.3 室内模拟实验2017年4月, 采集研究区芦苇湿地表层(0—20 cm)土壤样品, 实验室内手工挑出石块、动植物残体等杂物, 过2 mm筛, 于4℃冷藏备用。采用环刀法测试表层土壤最大持水率(WHC)[32]。取20 g(干重)新鲜土样于500 mL广口瓶, 并加入0.2 g野外实验收集、不同分解日期的凋落物样品(粉碎、过0.1 mm筛), 混匀;设置无凋落物的土样作为对照;用去离子水调节土壤含水率至100% WHC, 称重并记录, 作为后期补充水分的依据。将上述培养瓶于10℃、20℃、30℃恒温培养箱内培养;每个培养温度条件下各处理设置3个重复。
在培养实验的第1、3、8、15、25、40、60天, 抽取瓶内空气样品。采样时, 用大功率风扇对广口瓶逐个通风2 min, 用新鲜空气置换瓶内空气, 随即用带三通的胶塞密封瓶口, 并放入相应培养箱内培养5 h。取样时, 用带三通的注射器抽取瓶内空气50 mL, 采用气相色谱法(Aglient 7890A, USA)测试CO2浓度。含凋落物土样的呼吸速率与对照的差值即为凋落物呼吸速率。采样结束后, 打开胶塞上的三通, 使瓶内外空气流通。每隔3—5 d采用称重法补充瓶内水分, 使之保持恒定的土壤含水率。将相邻两次测得的呼吸速率均值乘以其时间间隔, 即得该时段CO2的累积释放量, 将各时段CO2的累积释放量相加, 即得凋落物总的CO2累积释放量。
凋落物呼吸的温度敏感性(Q10)采用如下方法计算[33]:
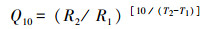
|
式中, R1和R2分别代表 10℃(T1)和30℃(T2)条件下凋落物呼吸速率。
1.4 凋落物有机碳分子结构测定凋落物有机碳分子结构测试采用核磁共振仪(AVANCE Ⅲ 400MH, 布鲁克公司)、固态13C-交叉极化魔角旋转技术测试, 转子直径4 mm, 13C谱共振频率为100.62 MHz, 旋转频率5000 Hz, 接触时间2 ms, 循环延迟时间3 s。核磁共振功能基团面积积分用MestReNova软件(v.12.0)分析, 共划分7个功能区[11-13]:烷基碳(0—45 ppm, alkyl C), 主要来自角质、蜡质等植物聚合物及微生物代谢物;甲氧基碳(45—60 ppm, methoxyl C), 是木质素的重要官能团;烷氧碳(60—90 ppm, O-alkyl C)、双烷氧碳区(90—110 ppm, di-O-alkyl C), 主要来自纤维素、半纤维素等碳水化合物;芳香碳(110—140 ppm, H- and C-substituted aromatic C;140—160 ppm, O-substituted aromatic C), 主要来自木质素等;羰基碳(160—190 ppm, carboxyl C), 主要来自脂肪酸、氨基酸、酰胺、酯等。其中, 烷基碳+芳香碳=疏水碳, 烷氧碳+羰基碳=亲水碳。
1.5 统计分析采用单因素方差分析(LSD法)统计不同分解日期凋落物失重率、元素含量及生态化学计量比、有机碳官能团相对含量、凋落物有机碳稳定性指标、凋落物CO2累积释放量及凋落物呼吸温度敏感性(Q10)等指标的差异, 显著性水平P=0.05;采用Pearson相关分析和逐步回归分析凋落物失重率及剩余率、凋落物CO2累积释放量及凋落物呼吸温度敏感性(Q10)与生态计量比、有机碳官能团相对含量、凋落物有机碳稳定性指标之间的关系。采用SPSS软件(v.17.0)进行数据统计, 采用Origin 8.0进行绘图。
2 结果与分析 2.1 凋落物分解及元素含量前70 d凋落物失重较少, 失重率仅为7.3%;第160—720天凋落物失重率显著增加(P < 0.05), 160 d时失重率达32.3%, 360 d时失重率达39.9%, 720 d时失重率为50.3%(图 1)。

|
| 图 1 凋落质量损失率 Fig. 1 Mass loss rate of litter |
相对于实验初始(第0天), 凋落物分解过程中, 有机碳含量基本无明显变化, 仅第160天的有机碳含量有明显增加(P < 0.05)。前70 d凋落物总氮含量无显著变化;第160天后, 凋落物总氮含量较实验初期显著增加(P < 0.05), 增幅达57.33%(160 d)、55.30%(360 d)、73.52%(720 d)。第30天, 凋落物总磷含量较实验初始(第0天)下降了70.67%;其后总磷含量显著增加, 增幅达61.54%(70 d)、187.50%(160 d)、215.38%(360 d)、206.25%(720 d;图 2)。前70 d凋落物C/N无显著变化, 第160—720天显著降低(P < 0.05)。第30天凋落物C/P明显增加, 其后C/P显著降低(P < 0.05)。第30天凋落物N/P明显增加(P < 0.05), 其后N/P无显著变化(图 3)。

|
| 图 2 凋落物元素含量 Fig. 2 Concentrations of organic carbon, nitrogen and phosphorus of litter |

|
| 图 3 凋落物生态化学计量比 Fig. 3 C/N, C/P, and N/P ratios of litter |
前70 d凋落物烷基碳相对含量无明显变化, 其后则明显增加(P < 0.05);甲氧基碳相对含量仅在第720天显著增加(P < 0.05);烷氧碳及双烷氧碳相对含量在实验初期均无显著变化, 其后则明显下降(P < 0.05);芳香碳相对含量在第30天无明显变化, 其后则显著增加(P < 0.05);羰基碳相对含量随分解时间延长逐渐增加(P < 0.05;图 4)。疏水碳/亲水碳、芳香碳/烷氧碳在第30天无明显变化, 其后则显著增加(P < 0.05);烷基碳/烷氧碳在前70 d无显著变化, 其后则逐渐增加(P < 0.05;图 5)。

|
| 图 4 凋落物有机碳分子结构 Fig. 4 Molecular structure of organic carbon of litter |

|
| 图 5 凋落物有机碳稳定性指数 Fig. 5 Index of organic carbon stability of litter |
10℃培养条件下, 野外分解30 d的凋落物CO2累积释放量与第0天的无明显差异;野外分解70 d及分解时间更长的凋落物CO2累积释放量随凋落物分解时间的延长逐渐下降, 降幅达21.00%(第70天)、32.60%(第160天)、37.26%(第360天)、54.45%(第720天)。20℃、30℃培养条件下, 野外分解30及70 d的凋落物CO2累积释放量与分解0天的无明显差异, 野外分解160天及分解时间更长的凋落物CO2累积释放量随凋落物分解时间的延长逐渐下降。20℃培养条件下, 相对于分解0天的凋落物, CO2累积释放量降幅达20.66%(第160天)、27.77%(第360天)、49.73%(第720天);30℃培养条件下, 相对于分解0天的凋落物, CO2累积释放量降幅达13.02%(第160天)、28.94%(第360天)、34.06%(第720天;图 6)。分解0—720天的凋落物呼吸温度敏感性(Q10)处于1.52—1.83之间。随着凋落物野外分解时间延长, 凋落物呼吸温度敏感性Q10值逐渐增加(P < 0.05), 第720天的凋落物呼吸温度敏感性最大(图 7)。

|
| 图 6 凋落物CO2累积释放量 Fig. 6 Cumulative CO2 emissions of litter |

|
| 图 7 凋落物呼吸的温度敏感性 Fig. 7 Temperature sensitivity of litter respiration |
由表 1可知, 有机碳组分相对含量及有机碳稳定性指标与凋落物分解及其CO2释放均呈显著相关关系(P < 0.05), 而且有机碳组分相对含量及有机碳稳定性指标与凋落物呼吸温度敏感性(Q10)也存在显著相关关系(P < 0.05)。表明凋落物分解与有机碳质量及生态化学计量比之间存在密切关系。由于很多指标与凋落物分解及其温度敏感性存在共线性关系, 单因素相关分析结果不能准确解释有机碳质量及生态化学计量比对凋落物分解及其温度敏感性的关系。因此, 采用逐步回归的方法, 将各有机碳质量及生态化学计量比指标逐步纳入回归模型, 回归模型如下:
| 项目 Items |
烷基碳 Alkyl C |
甲氧基碳 Methoxyl C |
烷氧碳 O-Alkyl C |
双烷氧碳 di-O-Alkyc C |
芳香碳 H- and C-substituted aromatic C |
芳香碳 O-substituted aromatic C |
羰基碳 Carbonyl C |
C/N | C/P | N/P | HB/HI | 烷基碳/烷氧碳 Alkyl C/O-Alkyl C |
芳香碳/烷氧碳 Aromatic C/O-Alkyl C |
| Q10 | 0.38 | 0.41 | -0.52** | -0.49* | 0.53* | 0.50* | 0.56* | -0.32 | -0.22 | -0.19 | 0.47* | 0.43 | 0.52* |
| CP10 | -0.84** | -0.71** | 0.91** | 0.91** | -0.74** | -0.71** | -0.88** | 0.78** | 0.58* | 0.50* | -0.90** | -0.89** | -0.87** |
| CP20 | -0.86** | -0.81** | 0.86** | 0.89** | -0.61** | -0.56* | -0.78** | 0.82** | 0.56* | 0.47* | -0.86** | -0.89** | -0.79** |
| CP30 | -0.87** | -0.75** | 0.85** | 0.88** | -0.58* | -0.55 | -0.75** | 0.85** | 0.63** | 0.55* | -0.86** | -0.89** | -0.78** |
| MLR | 0.94** | 0.74** | -0.97** | -0.97** | 0.76** | 0.71** | 0.89** | -0.88** | -0.57* | -0.48* | 0.97** | 0.97** | 0.91** |
| MLR | -0.94** | -0.74** | 0.97** | 0.97** | -0.76** | -0.71** | -0.89** | 0.88** | 0.57* | 0.48* | -0.97** | -0.97** | -0.91** |
| CP10:10度条件下凋落物CO2累积释放量,cumulative production of CO2 at 10 degree; CP20:20度条件下凋落物CO2累积释放量,cumulative production of CO2 at 20 degree; CP30:30度条件下凋落物CO2累积释放量,cumulative production of CO2 at 30 degree; MLR:凋落物失重率, mass loss rate of litter; RRL:凋落物剩余率, rate of residual litter; HB/HI:疏水碳/亲水碳, hydrophobic C/hydrophilic C; *在0.05水平(双侧)上显著相关;**在0.01水平(双侧)上显著相关 | |||||||||||||
凋落物失重率(%)=249.403 X1-0.393 X2+29.916, P < 0.05, r2=0.97, F=184.048;
10度CO2累积释放量=29.272 X3-448.574, P < 0.001, r2=0.836, F=87.821;
20度CO2累积释放量=297.307 X3-1478.786, P < 0.01, r2=0.793, F=61.366;
30度CO2累积释放量=247.693 X3-921.646, P < 0.05, r2=0.787, F=63.917;
凋落物呼吸温度敏感性(Q10)=0.026 X4+1.553, P < 0.001, r2=0.276, F=7.465。
式中, X1为烷基碳/烷氧碳;X2为C/N;X3为烷氧碳;X4为羰基碳。
3 讨论凋落物有机碳质量是影响凋落物分解的内在因素[34]。凋落物分解初期, 大量水溶性有机质快速淋溶释放, 为微生物的生长繁殖提供养分与能量[35], 这可能进一步促进微生物对凋落物的降解。凋落物分解后期, 随着缩合单宁、木质素、角质等难溶的和不规则的高分子有机物的富集, 凋落物分解的抵抗性增加[36]。本研究结果显示, 随着凋落物分解时间延长, 凋落物易分解有机碳组分(烷氧碳、双烷氧碳)相对含量随之下降, 而难分解有机碳组分(烷基碳、芳香碳)相对含量逐渐增加(图 4)。这与谌贤等[37]关于川西高山森林凋落物分解的研究结果相似。疏水碳/亲水碳、芳香碳/烷氧碳、烷基碳/烷氧碳是反映有机碳质量的重要指标, 其值越大, 有机碳越稳定[11-13]。本研究也发现, 随着凋落物分解, 疏水碳/亲水碳、芳香碳/烷氧碳、烷基碳/烷氧碳也随之增大(图 5);凋落物CO2累积释放量也逐渐降低(图 6);同时, 分解第160—720天凋落物失重率(18%)也低于前160天的失重率(32.3%)。表明:随着凋落物分解, 易分解碳组分在逐渐消耗, 而难分解碳组分在累积, 有机碳稳定性逐渐增加, CO2释放量及失重率也在逐渐下降。
有机碳质量是影响凋落物分解的重要因素, 其影响凋落物分解的重要性甚至高于生态化学计量比[9, 11]。本研究发现, 凋落物CO2累积释放量分别与有机碳易分解组分相对含量呈显著正相关关系(P < 0.001), 与难分解有机碳组分相对含量以及疏水碳/亲水碳、芳香碳/烷氧碳、烷基碳/烷氧碳均呈显著负相关关系(P < 0.05;表 1)。然而, 凋落物失重率与疏水碳/亲水碳、芳香碳/烷氧碳、烷基碳/烷氧碳均呈显著负相关关系(P < 0.01);且凋落物剩余率与易分解碳组分相对含量呈显著正相关, 而与难分解碳组分相对含量呈显著负相关关系(P < 0.01;表 1)。表明:凋落物易分解碳组分含量越低, 凋落物有机碳越稳定, 其分解速率越小。据此推测, 有机碳质量是预测凋落物分解及其CO2释放的敏感性指标。
凋落物生态化学计量比与有机碳质量均对凋落物的分解有显著影响, 然而两因素对凋落物分解的影响仍存争议。研究表明, 生态化学计量比也是控制凋落物分解的主要因素[8];其中, C/N是预测凋落物分解的敏感指标[38]。也有研究发现, 相对于生态化学计量比, 有机碳质量能更好的解释凋落物的分解[9, 11]。本研究发现, 生态化学计量比(C/N、C/P、N/P)与凋落物失重率及CO2累积释放量呈显著相关关系(P < 0.05;表 1), 这与Parton等[38]的结论相似。意味着生态化学计量比也能显著影响凋落物分解及CO2释放。逐步回归结果显示, 烷基碳、烷氧碳相对含量及C/N能很好的解释凋落物失重率, 而烷氧碳及双烷氧碳相对含量可以较好的解释CO2累积释放量。据此推断, 有机碳质量与生态化学计量比均是预测凋落物分解及CO2释放的敏感性指标, 且有机碳质量较生态化学计量比更为敏感。
有机碳质量能够影响有机碳分解的温度敏感性, 但影响结果仍不确定[23-26]。本研究发现, 凋落物分解时间越长, 有机碳越稳定(图 4、5), 凋落物呼吸的温度敏感性Q10值越大(图 7)。这暗示有机碳难分解组分CO2释放的温度敏感性可能大于有机碳易分解组分。这与Vanhala等[24]与Conant等[25]关于土壤有机碳组分矿化温度敏感性的结论相同。其原因可能是:根据化学动力学理论(Chemical kinetic theory), 难分解有机物具有较高的活化能(activation energy), 故难分解有机质矿化具有较高的温度敏感性[39-40]。而且, Q10值与凋落物有机碳难分解组分(芳香碳, 110—140 ppm)及有机碳稳定性指数(疏水碳/亲水碳)均呈显著正相关关系(P < 0.05), 而与凋落物有机碳易分解组分(烷氧碳、双烷氧碳)相对含量呈显负相关关系(P < 0.05), 这进一步解释了Q10值的上述变化趋势。由此可知, 随着凋落物分解, 有机碳的稳定性逐渐增加, CO2释放速率虽然降低, 但其对温度的响应程度则进一步增加, 这可能进一步增加其对全球变暖的积极响应。此外, Q10值与生态化学计量比无显著相关性(表 1)。而逐步回归的结果显示, 羰基碳相对含量可以用来解释Q10的变异, 但仅能解释27.6%的变异。表明仍有其他显著影响Q10的因素未在本研究考虑范围内, 这需要在后续实验中进一步探索。据此推断:相对于生态化学计量比, 有机碳质量可以解释凋落物呼吸温度敏感性的变化, 是预测凋落物呼吸温度敏感性相对敏感的指标。综上所述, 烷基碳、烷氧碳、双烷氧碳的相对含量是预测凋落物分解的敏感性指标, 而羰基碳相对含量是预测凋落物呼吸温度敏感性的相对敏感的指标。
4 结论(1) 随着凋落物分解, 烷氧碳、双烷氧碳相对含量逐渐降低, 而芳香碳的相对含量显著增加, 疏水碳/亲水碳、芳香碳/烷氧碳比值逐渐增大, 凋落物有机碳更加稳定, 凋落物呼吸速率及失重率呈下降趋势。烷基碳、烷氧碳相对含量及C/N能很好的解释凋落物失重率, 而烷氧碳及双烷氧碳相对含量可以较好的解释CO2累积释放量。表明有机碳质量及生态化学计量比均是预测凋落物分解及CO2释放的敏感性指标, 且有机碳质量较生态化学计量比更为敏感。
(2) 随着凋落物分解, 有机碳稳定性逐渐增强, 凋落物呼吸温度敏感性显著增加。而且, 羰基碳相对含量可以用来解释Q10的变异。总之, 相对于生态化学计量比, 烷基碳、烷氧碳、双烷氧碳、羰基碳相对含量是预测凋落物分解及其温度敏感性的敏感性指标。
致谢: 本研究承蒙中国科学院黄河三角洲滨海湿地生态试验站支持, 特此致谢。| [1] |
Sinsabaugh R L, Carreiro M M, Repert D A. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss. Biogeochemistry, 2002, 60(1): 1-24. DOI:10.1023/A:1016541114786 |
| [2] |
Gessner M O, Swan C M, Dang C K, McKie B G, Bardgett R D, Wall D H, Hättenschwiler S. Diversity meets decomposition. Trends in Ecology & Evolution, 2010, 25(6): 372-380. |
| [3] |
Coûteaux M M, Bottner P, Berg B. Litter decomposition, climate and liter quality. Trends in Ecology & Evolution, 1995, 10(2): 63-66. |
| [4] |
Moorhead D L, Currie W S, Rastetter E B, Parton W J, Harmon M E. Climate and litter quality controls on decomposition:an analysis of modeling approaches. Global Biogeochemical Cycle, 1999, 13(2): 575-589. DOI:10.1029/1998GB900014 |
| [5] |
Gholz H L, Wedin D A, Smitherman S M, Harmon M E, Parton W J. Long-term dynamics of pine and hardwood litter in contrasting environments:toward a global model of decomposition. Global Change Biology, 2000, 6(7): 751-765. DOI:10.1046/j.1365-2486.2000.00349.x |
| [6] |
Cornwell W K, Cornelissen J H C, Amatangelo K, Dorrepaal E, Eviner V T, Godoy O, Hobbie S E, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested H M, Santiago L S, Wardle D A, Wright I J, Aerts R, Allison S D, van Bodegom P, Brovkin V, Chatain A, Callaghan T V, Díaz S, Garnier E, Gurvich D E, Kazakou E, Klein J A, Read J, Reich P B, Soudzilovskaia N A, Vaieretti M V, Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecology Letter, 2008, 11(10): 1065-1071. DOI:10.1111/j.1461-0248.2008.01219.x |
| [7] |
Aerts R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems:a triangular relationship. Oikos, 1997, 79(3): 439-449. DOI:10.2307/3546886 |
| [8] |
Melillo J M, Aber J D, Muratore J F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology, 1982, 63(3): 621-626. DOI:10.2307/1936780 |
| [9] |
Hättenschwiler S, Jørgensen H B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. Journal of Ecology, 2010, 98(4): 754-763. DOI:10.1111/j.1365-2745.2010.01671.x |
| [10] |
Aerts R, van Bodegom P M, Cornelissen J H C. Litter stoichiometric traits of plant species of high-latitude ecosystems show high responsiveness to global change without causing strong variation in litter decomposition. New Phytologist, 2012, 196(1): 181-188. DOI:10.1111/j.1469-8137.2012.04256.x |
| [11] |
Bonanomi G, Incerti G, Giannino F, Mingo A, Lanzotti V, Mazzoleni S. Litter quality assessed by solid state 13C NMR spectroscopy predicts decay rate better than C/N and Lignin/N ratios. Soil Biology and Biochemistry, 2013, 56: 40-48. DOI:10.1016/j.soilbio.2012.03.003 |
| [12] |
张勇, 胡海波, 黄玉洁, 李土生, 周子贵, 郭晓平, 高海力. 不同植被恢复模式对土壤有机碳分子结构及其稳定性的影响. 环境科学研究, 2015, 28(12): 1870-1878. |
| [13] |
郭素春, 郁红艳, 朱雪竹, 高彦征, 丁维新. 长期施肥对潮土团聚体有机碳分子结构的影响. 土壤学报, 2013, 50(5): 922-930. |
| [14] |
Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B, 1992, 44(2): 81-99. DOI:10.3402/tellusb.v44i2.15428 |
| [15] |
Kim H, Hirano T, Koike T, Urano S. Contribution of litter CO2 production to total soil respiration in two different deciduous forests. Phyton, 2005, 45(4): 385-388. |
| [16] |
Liang N, Hirano T, Zheng Z M, Tang J, Fujinuma Y. Soil CO2 efflux of a larch forest in northern Japan. Biogeosciences, 2010, 7(11): 3447-3457. DOI:10.5194/bg-7-3447-2010 |
| [17] |
Rey A, Pegoraro E, Tedeschi V, De Parri I, Jarvis P G, Valentini R. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Global Change Biology, 2002, 8(9): 851-866. DOI:10.1046/j.1365-2486.2002.00521.x |
| [18] |
Sulzman E W, Brant J B, Bowden R D, Lajtha K. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest. Biogeochemistry, 2005, 73(1): 231-256. DOI:10.1007/s10533-004-7314-6 |
| [19] |
Zhang Y J, Gan Z T, Li R J, Wang R, Li N N, Zhao M, Du L L, Guo S L, Jiang J S, Wang Z Q. Litter production rates and soil moisture influences interannual variability in litter respiration in the semi-arid Loess Plateau, China. Journal of Arid Environments, 2016, 125: 43-51. DOI:10.1016/j.jaridenv.2015.09.016 |
| [20] |
张彦军. 凋落物呼吸温度敏感性的变化特征及其影响因素. 环境科学, 2017, 38(8): 3497-3506. |
| [21] |
范少辉, 唐晓鹿, 漆良华, 刘广路, 官凤英, 杜满义. 不同经营措施对毛竹林土壤呼吸温度敏感性的影响. 四川农业大学学报, 2012, 30(3): 300-307. DOI:10.3969/j.issn.1000-2650.2012.03.007 |
| [22] |
苏爱玲.气候变化对高寒草甸枯落物分解特性的影响及温度敏感性研究[D].西宁: 中国科学院研究生院, 2010.
|
| [23] |
Fissore C, Giardina C P, Swanston C W, King G M, Kolka R K. Variable temperature sensitivity of soil organic carbon in North American forests. Global Change Biology, 2009, 15(9): 2295-2310. DOI:10.1111/j.1365-2486.2009.01903.x |
| [24] |
Vanhala P, Karhu K, Tuomi M, Sonninen E, Jungner H, Fritze H, Liski J. Old soil carbon is more temperature sensitive than the young in an agricultural field. Soil Biology and Biochemistry, 2007, 39(11): 2967-2970. DOI:10.1016/j.soilbio.2007.05.022 |
| [25] |
Conant R T, Drijber R A, Haddix M L, Parton W J, Paul E A, Plante A F, Six J, Steinweg J M. Sensitivity of organic matter decomposition to warming varies with its quality. Global Change Biology, 2008, 14(4): 868-877. DOI:10.1111/j.1365-2486.2008.01541.x |
| [26] |
Conen F, Leifeld J, Seth B, Alewell C. Warming mineralizes young and old soil carbon equally. Biogeosciences, 2006, 3: 515-519. DOI:10.5194/bg-3-515-2006 |
| [27] |
潘萍, 赵芳, 欧阳勋志, 臧颢, 宁金魁, 国瑞. 马尾松林两种林下植被土壤碳氮特征及其与凋落物质量的关系. 生态学报, 2018, 38(11): 3988-3997. |
| [28] |
Fennessy M S. Wetland ecosystems and global change//Freedman B, ed. Global Environmental Change. Dordrecht: Springer, 2014: 255-261.
|
| [29] |
Hiraishi T, Krug T, Tanabe K, Srivastava N, Baasansuren J, Fukuda M, Troxler T G. 2013 Supplement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories: Wetlands. Switzerland: IPCC, 2014.
|
| [30] |
朱敏, 张振华, 于君宝, 吴立新, 韩广轩, 杨利琼, 邢庆会, 谢宝华, 毛培利, 王光美. 氮沉降对黄河三角洲芦苇湿地土壤呼吸的影响. 植物生态学报, 2013, 37(6): 517-529. |
| [31] |
Han G X, Yang L Q, Yu J B, Wang G M, Mao P L, Gao Y J. Environmental controls on net ecosystem CO2 exchange over a reed (Phragmites australis) wetland in the Yellow River delta, China. Estuaries and Coasts, 2013, 36(2): 401-413. DOI:10.1007/s12237-012-9572-1 |
| [32] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999.
|
| [33] |
Fissore C, Giardina C P, Kolka R K, Trettin C C. Soil organic carbon quality in forested mineral wetlands at different mean annual temperature. Soil Biology and Biochemistry, 2009, 41(3): 458-466. DOI:10.1016/j.soilbio.2008.11.004 |
| [34] |
Berg B, Johansson M B, Meentemeyer V. Litter decomposition in a transect of Norway spruce forests:substrate quality and climate control. Canadian Journal of Forest Research, 2000, 30(7): 1136-1147. DOI:10.1139/x00-044 |
| [35] |
Almendros G, Dorado J, Gonzâlez-Vila F J, Blanco M J, Lankes U. 13C NMR assessment of decomposition patterns during composting of forest and shrub biomass. Soil Biology and Biochemistry, 2000, 32(6): 793-804. DOI:10.1016/S0038-0717(99)00202-3 |
| [36] |
Preston C M, Nault J R, Trofymow J A, Smyth C, CIDET Working Group. Chemical changes during 6 years of decomposition of 11 litters in some Canadian Forest sites. Part 1. Elemental composition, tannins, phenolics, and proximate fractions. Ecosystems, 2009, 12(7): 1053-1077. DOI:10.1007/s10021-009-9266-0 |
| [37] |
谌贤, 刘洋, 唐实玉, 杨帅, 陈亚梅, 杨林, 郑海峰, 李洪杰. 川西亚高山森林凋落物不同分解阶段基质质量特征. 西北植物学报, 2017, 37(3): 586-594. |
| [38] |
Parton W, Silver W L, Burke I C, Grassens L, Harmon M E, Currie W S, King J Y, Adair E C, Brandt L A, Hart S C, Fasth B. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science, 2007, 315(5810): 361-364. DOI:10.1126/science.1134853 |
| [39] |
Bosatta E, Ågren G I. Soil organic matter quality interpreted thermodynamically. Soil Biology and Biochemistry, 1999, 31(13): 1889-1891. DOI:10.1016/S0038-0717(99)00105-4 |
| [40] |
Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165-173. DOI:10.1038/nature04514 |