 2019, Vol. 39
2019, Vol. 39文章信息
- 焦亮, 刘雪蕊, 王圣杰, 董小刚
- JIAO Liang, LIU Xuerui, WANG Shengjie, DONG Xiaogang
- 敦煌阳关湿地芦苇克隆构件空间拓展策略及其对土壤环境因子的响应
- Spatial expansion strategy and soil environmental response of the clonal architectures for Phragmites australis in Yangguan wetland of Dunhuang
- 生态学报. 2019, 39(14): 5317-5325
- Acta Ecologica Sinica. 2019, 39(14): 5317-5325
- http://dx.doi.org/10.5846/stxb201809192043
-
文章历史
- 收稿日期: 2018-09-19
- 修订日期: 2019-03-05





植物对生长环境的响应是环境综合因素(气候、干扰、生物等)筛选效应的结果[1]。“克隆植物”(clonal plants)就是在自然生境条件下, 能通过营养繁殖产生与“母性”个体在基因上完全一致的新个体的植物[2]。因其具有强大的繁殖力和空间拓展性能强烈地影响植物群落的生长格局, 在许多生态系统中居于优势地位, 并对生态系统的恢复具有重要的意义[3-4]。环境异质性会促使克隆植物性状通过权衡机制发生变化, 进而调整克隆构件间的功能组合和资源配置, 形成相应的生态适应策略[5]。同时, 其克隆构件在异质环境下表现出不同的空间拓展策略来更有效的利用资源和空间。传统的研究认为“密集型”(Phalanx)和“游击型”(Guerilla)是克隆植物两种不同的克隆生长策略[6-7]。选择最优的生存策略能使克隆植物迅速占据生长空间, 进而提高种群的适合度[8]。“密集型”克隆植物表现为“入侵姿态”, 通过集群分布加强竞争能力, 主要表现为间隔子短、分枝角度小, 分株数多;而“游击型”克隆植物则采取“避让态度”, 通过克隆根系向外“觅食”扩张, 主要表现为间隔子长、分枝角度大, 分株数少[9]。
芦苇(Phragmites australis)属于多年生根茎型禾本科克隆植物, 具有极强的环境适应能力和重要的生态价值, 是内陆盐沼湿地分布面积最广的优势种和建群种之一, 被广泛应用于湿地生态系统的恢复与重建[10-11]。目前有关芦苇的研究多侧重于生理生化特征研究[12-13]、生物学特征对环境因子的响应[14-15]、克隆生长和有性繁殖的权衡[16-17]、种群生态系统功能及经济价值[18-19]等方面, 对于芦苇种群克隆构件空间拓展的策略研究关注较少, 而且土壤环境因子对克隆构件驱动力的研究也相对匮乏。干旱区内陆盐沼湿地是江河源区及绿洲重要的水源涵养地, 在维护生态安全、气候安全、淡水安全和生物多样性等方面发挥着不可替代的作用[20]。因此, 我们选择敦煌阳关国家级自然保护区湿地内不同环境梯度条件下的克隆植物芦苇开展研究, 分析芦苇种群对异质环境的生态适应性, 旨在解决以下问题:(1)对比不同环境梯度下芦苇各克隆构件的差异性, 探讨芦苇对异质性环境的空间拓展策略;(2)分析芦苇各克隆构件的协同进化关系及对主要土壤环境因子的响应, 明晰芦苇克隆生长及空间拓展的主要的驱动力。通过本研究有助于从克隆构件的角度明晰植物种群空间拓展与土壤环境因子关系, 进一步丰富克隆植物生态学研究。
1 研究地区与研究方法 1.1 研究区域和样地概况研究区敦煌阳关湿地隶属于敦煌阳关国家级自然保护区(93°53′ — 94°17′E, 39°39′ — 40°05′N), 总面积8.82 × 104 hm2, 海拔1150 — 1500 m, 四周被沙漠、戈壁包围(图 1)。研究区属典型的大陆性干旱气候, 昼夜温差大, 年均气温9.3 ℃, 全年降水稀少, 年均降水量39.9 mm, 年蒸发量2465 mm。保护区主要发育了风沙土、棕漠土、草甸土等, 主要植被类型为温带荒漠植被, 以芦苇、赖草(Leymus secalinus)、黑果枸杞(Lycium ruthenicum)、甘草(Glycyrrhiza uralensis)、疏叶骆驼刺(Alhagi sparsifolia)、罗布麻(Apocynum venetum)、盐角草(Salicornia europaea)等植物为主。

|
| 图 1 研究区地理位置和样地分布示意图 Fig. 1 Location of studying area and distribution of sampling sites |
2017年9月中旬, 在敦煌阳关湿地渥洼池保护站, 对植物和土壤进行了综合野外调查的基础上, 沿湿地到荒漠的方向由内向外平行设置了3条采样线, 每条采样线基于离水库500 m、1500 m、2500 m的距离和芦苇种群的密度与盖度设定了近水区(平均密度88%, 平均盖度90%)、过渡区(平均密度50%, 平均盖度55%)、荒漠区(平均密度20%, 平均盖度23%)3个不同的采样梯度, 每一个梯度上随机设置3个5 m×5 m的采样样方, 并记录每个采样样方的海拔、经纬度、地理地貌等小环境因素。
基于克隆植物群落特征, 以芦苇分株作为取样单位, 在每个样方随机选取3株芦苇, 按照克隆构件采集方法采用“全挖法”进行“跟踪挖掘”[21-22], 对选定植株地上及地下部分进行全部收获, 带回实验室使用卷尺测量植株高度、根茎节间长(地下横走根茎的两节之间长度)、间隔子长(克隆分株间的距离);使用游标卡尺测量茎粗、使用量角器测量分枝角度(分枝与根状茎之间夹角);通过计数法直接记录初级根茎条数(地下横走的根茎连接的根茎)、分株数(同一基株内, 形体和生理独立的, 或具备潜在形体和生理独立性的最小个体单位)和芽数(亲代藉由细胞分裂产生子代, 但是子代并不立即脱离母体, 而与母体相连, 继续接受母体提供的养分, 直到个体可独立生活才脱离母体, 是一种特殊的无性生殖方式)。随后用剪刀将植株材料分为叶、地上茎、地下茎、水平根茎、根等克隆构件, 并将所有样品统一编号后, 立即放入冷藏箱(0—4℃)中保存。
土壤取样点与植物取样点一一对应, 在每个样方内, 利用环刀法按照每10 cm取土的方法取0—100 cm的土壤, 混合0—30 cm、30—60 cm、60—100 cm的土壤为表层土、中层土、深层土, 重复采样3次。将采集的土壤样去杂、风干、研磨过筛后, 采用烘干称重法测定土壤含水量, 采用环刀法测量土壤容重, 采用PHS-SD型pH仪(同德, 北京同德创业科技有限公司)测定土壤pH值, 采用电导率法测定土壤含盐量。
1.3 数据处理不同环境条件间的克隆构件的差异性主要使用的是方差分析(One-way ANOVA)和LSD(Least significant difference)显著性多重比较, 利用SPSS20.0软件中的Kolmogorov-Smirnov对数据进行检验, 均符合正态分布(P < 0.05)。在正态分布的基础上利用单因素方差分析了不同环境梯度下克隆构件、土壤环境因子的差异显著性, 并采用LSD方法进行显著性多重比较, 显著性水平设置为P < 0.05。
克隆构件间及其与土壤环境的相关性主要使用的是Pearson相关分析法, 其中Pearson相关性系数R的计算如公式(1)所示。
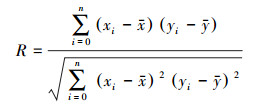
|
式中, n为样本数, xi和yi分别为两变量的变量值。
冗余分析Redundancy analysis(RDA)被广泛用于生物与环境因子的关系研究中, 是一种多变量直接梯度分析方法, 同时又是多元线性回归的扩展, 采用2个变量集的线性关系模型, 得到数值矩阵并对特征值进行分解, 能将表征芦苇各克隆构件指标及土壤环境因子之间的关系反映在坐标轴上, 并能够精确揭示物种与环境因子之间的关系。在进行冗余分析之前, 必须对因变量(芦苇各构件和各层土壤环境因子)进行DCA分析, 结果表明排序轴最大的梯度长度(Lengths of gradient, LGA)均小于3, 表明该数据均具有较好的线性反应, 即芦苇各克隆构件指标对环境梯度的响应是线性的, 对此利用线性响应模型分析(RDA)比较适宜。因此本文利用CANOCO中直接梯度分析方法RDA分析芦苇克隆构件对各层土壤环境因子(表、中、深层土壤含水量、容重、pH、盐分)的响应关系, 进一步揭示芦苇空间拓展的主要驱动力。实验数据用SPSS 20.0和CANOCO 5.0进行统计分析, Origin 9.0进行绘图。
2 结果分析 2.1 研究区土壤环境因子特征分析敦煌阳关湿地近水区-过渡区-荒漠区平均土壤含水量(0—100 cm)分别为(13.94±0.255)%、(10.42±0.542)%、(4.45±0.712)%, 呈现出逐渐减少的趋势;土壤容重、土壤含盐量呈现逐渐增加的趋势, 尤其是在荒漠区最为明显, 具体表现为荒漠区(容重:(1.31±0.006) g/cm3和盐分:(2.54±0.011)%)>过渡区(容重:(1.24±0.002) g/cm3和盐分:(2.35±0.028)%)>近水区(容重:(1.22±0.007) g/cm3和盐分:(2.12±0.060)%);近水区土壤pH值为7.92±0.029, 过渡区土壤pH值为8.17±0.008, 荒漠区土壤pH值为8.06±0.023, 土壤基本上呈弱碱性, 随环境梯度变化表现出出先降低再增加的趋势(表 1)。
| 0—100 cm平均 0—100 cm on average | 近水区 Close-water zone | 过渡区 Transition zone | 荒漠区 Desert zone |
| 土壤含水量Soil water content/% | 13.94±0.255a | 10.42±0.542b | 4.45±0.712c |
| 土壤容重Soil bulk density (g/cm3) | 1.22±0.007a | 1.24±0.002b | 1.31±0.006c |
| 土壤pH Soil pH | 7.92±0.029a | 8.17±0.008b | 8.06±0.023c |
| 土壤含盐量Soil salinity/% | 2.12±0.060a | 2.35±0.028b | 2.54±0.011c |
| 同行不同小写字母表示环境梯度间差异显著(P < 0.05; n = 9) | |||
从近水区到荒漠区, 随着环境梯度胁迫的增强, 芦苇全株高度、茎粗、分株数呈明显的下降趋势, 而芽数、初级根茎条数、根茎节间长、间隔子长、分枝角度呈明显的上升趋势(图 2)。其中芦苇空间拓展重要克隆构件表现为分株数近水区(4株±0.20株)>过渡区(3株±0.29株)>荒漠区(2株±0.05株);根茎节间长表现为荒漠区(10.22 cm±0.29 cm)>过渡区(6.56 cm±0.19 cm)>近水区(4.68 cm±0.28 cm);间隔子长表现为荒漠区(70.6 cm±2.34 cm)>过渡区(65.35 cm±2.67 cm)>近水区(30.2 cm±3.15 cm);而分枝角度表现为荒漠区(76.12°±2.33°)>过渡区(62.015°±2.47°)>近水区(38.75°±1.98°)。从统计学角度分析, 植株高度、分株数、初级根茎条数、根茎节间长、间隔子长、分枝角度等克隆构件在三个环境梯度下达到了显著差异(P < 0.05)。

|
| 图 2 不同环境条件下芦苇克隆构件特征(数值为平均值±标准误差) Fig. 2 Features of clonal architectures for Phragmites australis under different environmental conditions (mean±SE) 不同小写字母表示环境梯度间差异显著(P < 0.05; n = 9);1:近水区, 2:过渡区, 3:荒漠区 |
图 2表现出芦苇各克隆构件之间存在协同进化的关系, 主要表现为生长克隆构件植株高度和茎粗达到了显著正相关(R=0.716, P < 0.01), 而空间拓展克隆构件间也达到了显著相关(P < 0.01)。具体表现为除芽数外, 分株数与初级根茎条数、根茎节间长、间隔子长、分枝角度呈现显著负相关(P < 0.01);除芽数与初级根茎条数和分枝角度不相关外, 初级根茎条数、根茎节间长、间隔子长、分枝角度4个指标之间达到了显著正相关(P < 0.01)。
2.4 芦苇克隆构件对环境因子的响应芦苇的克隆构件主要受到0—100 cm平均土壤水分、pH值、盐分的影响(图 3)。土壤水分与植株高度、茎粗、分株数呈显著的正相关(P < 0.01), 而与芽数、初级根茎条数、根茎节间长、间隔子长、分枝角度呈现显著负相关(P < 0.01);土壤pH值与植株高度、分株数表现为显著负相关(P < 0.05), 与初级根茎条数、间隔子长、分枝角度呈现显著正相关(P < 0.01);土壤盐分与植株高度、茎粗、分株数呈显著的负相关(P < 0.01), 与初级根茎条数、根茎节间长、间隔子长、分枝角度呈现显著正相关(P < 0.01)。

|
| 图 3 芦苇空间拓展克隆构件指标与土壤环境因子(0—100 cm)的关系 Fig. 3 Relationship between the clonal architectures indexes of spatial expansion and soil environmental factors (0—100 cm) for Phragmites australis |
| 植株高度 Plant height | 茎粗 Stem diameter | 分株数 Ramet number | 芽数 Bud number | 初级根茎条数 P-R-N | 根茎节间长 R-I-L | 间隔子长 Spacer length | 分枝角度 Branch angle | |
| 植株高度Plant height | 1 | 0.716** | 0.427** | 0.05 | -0.552** | -0.755** | -0.451** | -0.627* |
| 茎粗Stem diameter | 1 | 0.276 | -0.18 | -0.334* | -0.552** | -0.241 | -0.265 | |
| 分株数Ramet number | 1 | -0.068 | -0.768** | -0.542** | -0.427** | -0.537** | ||
| 芽数Bud number | 1 | 0.145 | 0.345* | 0.386** | 0.204 | |||
| 初级根茎条数P-R-N | 1 | 0.794** | 0.705** | 0.804** | ||||
| 根茎节间长R-I-L | 1 | 0.667** | 0.790** | |||||
| 间隔子长Spacer length | 1 | 0.745** | ||||||
| 分枝角度Branch angle | 1 | |||||||
| *表示在0.05水平(双侧)上显著相关;**表示在0.01水平(双侧)上显著相关; P-R-N:Primary rhizome number初级根茎条数; R-I-L:Rhizome internode length根茎节间长 | ||||||||
冗余分析显示植株高度、茎粗、分株数与每一层的水分都达到显著的正相关(P < 0.01), 芽数、初级根茎条数、根茎节间长、间隔子长、分枝角度与每一层的水分都达到显著的负相关(P < 0.01)(图 4);植株高度、茎粗、分株数与深层土壤pH值都达到显著的负相关(P < 0.05), 初级根茎条数、根茎节间长、间隔子长、分枝角度与深层土壤pH值达到显著的正相关(P < 0.01)(图 4);植株高度、茎粗、分株数与表、深层土壤盐分显著负相关(P < 0.05), 初级根茎条数、根茎节间长、间隔子长、分枝角度与表、深层土壤盐分显著正相关(P < 0.01)(图 4)。这说明各层土壤水分、深层土壤pH值和盐分是芦苇空间拓展最重要的环境因子。

|
| 图 4 芦苇空间拓展克隆构件指标与水分、容重、pH、盐分土壤环境因子(表层:0—30 cm, 中层:30—60 cm, 深层:60—100 cm)的冗余分析(RDA) Fig. 4 Redundancy analysis (RDA) between the spatial expansion clonal architectures indicators and soil environmental factors (surface layer: 0—30 cm, middle layer: 30—60 cm, deep layer: 60—100 cm) of Water, Bulk Density, pH, Salt for Phragmites australis water1:表层水分, water2:中层水分, water3:深层水分, BD1:表层容重, BD:中层容重, BD3:深层容重, pH1:表层pH, pH2:中层pH, pH3:深层pH, salt1:表层盐分, salt2:中层盐分, salt3:深层盐分;A:植株高度, B:茎粗, C:分株数, D:芽数, E:初级根茎条数, F:根茎节间长, G:间隔子长, H:分枝角度 |
空间拓展不仅能够反映植物对异质性生境的适应能力, 而且体现了植物与环境权衡维度的生活史对策[23]。在长期的适应进化过程中, 克隆植物形成了有效利用异质性资源以及抵御各种局部胁迫的生态适应对策[24], 如不同生长构型之间的权衡关系[25]。本研究通过分析芦苇各克隆构件空间拓展策略及其对土壤环境因子的响应, 发现芦苇最佳生活史主要体现在二个方面, 一方面是异质环境中空间拓展的最优策略, 另一方面是各克隆构件间的协同进化关系。
研究发现从近水区向荒漠区演变过程中芦苇空间拓展主要构件初级根茎条数、根茎节间长、间隔子长和分枝角度逐渐增大, 而分株数逐渐减少(图 2), 这显示芦苇种群随着环境胁迫的增加, 空间拓展策略由“密集型”向“游击型”转变。研究结果表明当芦苇处于资源条件好的斑块中表现为“入侵姿态”, 克隆构型趋于“密集型”, 从而利于整个植株对资源的获取;而在资源条件相对较差的斑块中表现为“避让态度”, 克隆构型趋于“游击型”, 利于植株能逃离资源水平差的斑块, 向条件更好的斑块转移[26]。而且塔克拉玛干沙漠南缘疏叶骆驼刺[27]、毛乌素沙地沙棘(Hippophae rhamnoides)[28]、四川盆地西缘聚花过路黄(Lysimachia congestiflora)[29]的研究结果也证明了随着环境胁迫的增强植物克隆生长格局由“密集型”向“游击型”过渡的结论。同时, 研究结果显示随着环境梯度的恶化, 芦苇将更多的资源投入到根茎节间、间隔子等地下生物量以实现生存生长投入(图 2)。Rmstrong模拟了多年生根茎克隆植物生物量分配格局, 发现2/3的最佳资源分配于根茎的生长[30]。另外, 对兰州秦王川湿地芦苇[10]、四川西北沙生苔草(Carex praeclara)和赖草[31]研究也得到了相同的结论。这说明当植物生长所需要的资源受到严重胁迫时, 克隆植物可通过控制生物量投资进行耗益权衡, 达到对资源获取和利用效率的最大化, 实现空间拓展的最优策略, 并通过不同功能过程(生长、繁殖等)间的权衡形成最佳生活史对策[32]。
空间拓展是克隆植株各构件共同作用的结果, 构件间有一定的生理整合[33], 表现出协同进化关系。对敦煌阳关湿地研究发现, 芦苇克隆构件在不同环境梯度制约下差异显著, 具体表现为初级根茎条数、根茎节间长、间隔子长、分枝角度呈现“同增同涨”的协同进化关系(P < 0.01), 而与分枝数呈现“此消彼长”的权衡关系(P < 0.01)。腾格里沙漠克隆植物羊草(Leymus chinensis)和四川绵阳克隆植物蛇莓(Duchesnea indica)克隆构件之间的关系表现正好证实了研究结果[34-35]。在不同的环境条件下, 植物总是权衡各功能间(生长、繁殖和维持)的资源分配, 使其各功能分配达到最佳状态[36]。
3.2 芦苇克隆构件空间拓展的驱动力研究土壤环境因子是决定植物分布格局的主导因子[37], 其中土壤水分、容重、pH值、盐分是影响植物生长的最主要驱动力。研究发现敦煌阳关国家级自然保护区湿地芦苇克隆构件空间拓展的直接驱动力是土壤水分、pH值和盐分(图 3)。
干旱荒漠区年降水量稀少, 水分成为湿地生态系统的基本生态因子和主要控制因素[38]。研究结果显示, 土壤水分与芦苇的生长指标(植株高度、茎粗)显著正相关, 与空间拓展指标(初级根茎条数、根茎节间长、间隔子长和分枝角度)显著负相关(P < 0.01)。每一层的土壤水分对植株高度、茎粗等生长指标起促进作用, 对初级根茎条数、根茎节间长、间隔子长和分枝角度等空间拓展指标起抑制作用(图 4)。由于水分胁迫, 土壤溶液水势降低, 植物失水大于吸水, 破坏植物体内的水分代谢, 导致植物生理干旱缺水, 限制植物生长指标, 但促进空间拓展指标, 使其逃离胁迫环境。这说明当植物所处区域环境条件较好时更注重地上生物量的投资, 减少地下根茎的投入, 克隆植物各构件间的变异格局以及性状间的“投资-收益”权衡则要靠资源优化配置和补偿机制来调控[39]。因此, 土壤水分是植物空间拓展重要的驱动因子。
中国北方的植物主要受到盐碱化土壤的作用[10]。研究结果表明, 土壤的盐碱化对芦苇的生长指标起抑制作用, 尤其是深层盐分与芦苇植株高度、茎粗之间呈现显著的负相关(图 4)。盐碱化土壤中的盐碱含量过高, 造成土壤坚硬而密实, 团粒结构破坏, 土壤孔隙度减少, 有毒物质不断积累, 同时土壤中过量的Na+、Cl-和Ca2+渗入植物细胞内, 打破植物体力的离子平衡, 破坏叶绿素和蛋白质合成, 从而抑制植物生长繁殖[40]。松嫩平原南部和塔里木盆地芦苇克隆生长构件(株高、茎粗)对盐碱化土壤响应的研究结果也给予了证实[41-42]。同时研究还发现盐分对主要的克隆空间拓展构件(初级根茎条数、根茎节间长、间隔子长、分枝角度)起促进作用, 随着环境梯度土壤盐碱化程度的增加, 可以刺激克隆植物初级根茎条数、根茎节间长、间隔子长、分枝角度等空间拓展各构件的增加, 使植株能通过长距离扩散以期尽快逃离不利生境斑块。
随环境退化程度的加深, 研究区克隆植物芦苇的空间拓展策略从“密集型”转向“游击型”, 反映了植物应对胁迫生境的适应性机制, 以及降低投资风险实现生存、繁衍的更新策略。在内陆盐沼湿地生境斑块间可利用资源水平(土壤水分、pH值、盐分)是芦苇克隆构件空间拓展的主要驱动力, 其中各层土壤水分, 深层土壤pH值、盐分是影响更重要的土壤环境因子。在不同的环境退化梯度中, 克隆植物通过构建适宜的空间拓展策略和协同进化关系, 一方面进行有效的资源获取和利用, 另一方面通过不断调整克隆构件, 使植物种群能长时间的、更大规模的进行繁衍、拓展。随着未来内陆盐沼湿地的干旱程度的逐渐加剧, 克隆植物空间拓展也会随之改变, 因此, 对于不同环境梯度条件下的克隆植物应根据响应特征采取不同的管理策略。
致谢: 感谢敦煌阳关国家级自然保护区管理局为本研究提供的支持。| [1] |
刘羽霞, 许嘉巍, 靳英华, 张英洁, 牛莉平, 王嫒林, 贺红士. 长白山苔原草本植物入侵与土壤环境的关系. 生态学报, 2018, 38(04): 1235-1244. |
| [2] |
葛俊, 邢福. 克隆植物对种间竞争的适应策略. 植物生态学报, 2012, 36(06): 587-596. |
| [3] |
Cornelissen J H C, Song YB, Yu F H, Dong M. Plant trait sand ecosystem effects of clonality:a new research agenda. Annals of Botany, 2014, 114(2): 369-376. DOI:10.1093/aob/mcu113 |
| [4] |
Yu F H, Wang N, He W M, Dong M. Effects of clonal integration on species composition and biomass of sand dune communities. Journal of Arid Environments, 2010, 74(6): 632-637. DOI:10.1016/j.jaridenv.2009.12.008 |
| [5] |
Blonder B, Violle C, Enquist B J. Assessing the causes and scales of the leaf economics spectrum using venation networks in Populus tremuloides. Journal of Ecology, 2013, 101(4): 981-989. DOI:10.1111/1365-2745.12102 |
| [6] |
董鸣, 于飞海. 克隆植物生态学术语和概念. 植物生态学报, 2007, 31(04): 689-694. DOI:10.3321/j.issn:1005-264X.2007.04.020 |
| [7] |
Schmid B, Harper J L. Clonal growth in grassland perennials:1.Density and pattern-dependent competition between plants with different growth forms. The Journal of Ecology, 1985, 73(3): 793-808. DOI:10.2307/2260147 |
| [8] |
周资行, 焦健, 李毅, 李永兵, 张世虎. 民勤沙拐枣克隆种群构件结构及与环境因子的灰色关联分析. 林业科学, 2012, 48(05): 141-149. |
| [9] |
李尝君, 吕光辉, 贡璐, 张海峰, 刘晓星. 艾比湖湿地自然保护区克隆植物群落空间格局及其对水盐胁迫的响应. 干旱区研究, 2013, 30(01): 122-128. |
| [10] |
王继伟, 赵成章, 赵连春, 王小鹏, 李群. 内陆盐沼芦苇根系形态及生物量分配对土壤盐分因子的响应. 生态学报, 2018, 38(13): 4843-4851. |
| [11] |
Eller F, Skalova H, Caplan JS, et al. Cosmopolitan species as models for ecophysiological responses to global change:The common reed Phragmites australis. Frontiers in Plant Science, 2017, 8: 1833. DOI:10.3389/fpls.2017.01833 |
| [12] |
聂恺宏, 邹旭, 吉生丽, 姜准, 刘春红, 高海银, 李根前. 中国沙棘克隆生长对灌水强度的响应规律及其激素调控机制. 生态学报, 2018, 38(14): 4942-4952. |
| [13] |
Yang Z F, Xie T, Liu Q. Physiological responses of phragmites australis to the combined effects of water and salinity stress. Ecohydrology, 2014, 7(2): 420-426. DOI:10.1002/eco.v7.2 |
| [14] |
鱼腾飞, 冯起, 刘蔚, 司建华, 席海洋, 陈丽娟. 黑河下游土壤水盐对生态输水的响应及其与植被生长的关系. 生态学报, 2012, 32(22): 7009-7017. |
| [15] |
玛丽娅·奴尔兰, 刘卫国, 霍举颂, 李宏侠, 张雨, 刘建国, 徐悦. 旱生芦苇对地下水位变化的生态响应及适应机制. 生态学报, 2018, 38(20): 1-10. |
| [16] |
徐苏男, 刘一晨, 刘艳虹, 陈忠林, 李悦, 张利红. 养分异质条件下结缕草克隆分株生长及光合作用的生理整合. 应用生态学报, 2018, 29(03): 811-817. |
| [17] |
Van Drunen W E, van Kleunen M, Dorken M E. Consequences of clonality for sexual fitness:Clonal expansion enhances fitness under spatially restricted dispersal. Proceedings of the National Academy of Sciences, 2015, 112(29): 8929-8936. DOI:10.1073/pnas.1501720112 |
| [18] |
向泽宇, 张莉, 刘雪娟. 蚁丘干扰下高寒草甸植物群落特征和生态位. 生态学杂志, 2016, 35(02): 321-329. |
| [19] |
Wang Z Q, Wang L N, Wu D Q, Zhang Z G. The Actuality and Trend in the Study of Phragmites australis (Cav.) Trin. exSteudel. in China. Journal of Shandong Forestry Science and Technology, 2006, 6(36): 85-87. |
| [20] |
张剑, 王利平, 谢建平, 赵庭伟, 曹建军. 敦煌阳关湿地土壤有机碳分布特征及其影响因素. 生态学杂志, 2017, 36(09): 2455-2464. |
| [21] |
Harper J L. Population Biology of Plant. London and New York: Academic Press, 1977: 36-41.
|
| [22] |
董鸣, 于飞海, 等. 克隆植物生态学. 北京: 科学出版社, 2011: 5-7.
|
| [23] |
Pinno B D, Wilson S D. Nitrogen translocation between clonal mother and daughter trees at a grassland-forest boundary. Plant Ecology, 2014, 215(3): 347-354. DOI:10.1007/s11258-014-0305-3 |
| [24] |
郭京衡, 曾凡江, 李尝君, 张波. 塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略. 植物生态学报, 2014, 38(01): 36-44. |
| [25] |
Ye X H, Yu F H, Dong M. A trade-off between guerrilla and phalanx growth forms in Leymus secalinusunder different nutrient supplies. Annals of Botany, 2006, 98(1): 187-191. |
| [26] |
de Kroon H, Knops J. Habitat exploitation through morphological plasticity in two chalk grassland perennials. Oikos, 1990, 59(1): 39-49. DOI:10.2307/3545120 |
| [27] |
张利刚, 曾凡江, 袁娜, 刘波, 罗维成, 宋聪, 彭守兰. 不同水分条件下疏叶骆驼刺(Alhagi sparifolia)生长及根系分株构型特征. 中国沙漠, 2013, 33(03): 717-723. |
| [28] |
贺斌, 赵粉侠, 李根前, 马先锋, 徐德兵, 李甜江. 毛乌素沙地中国沙棘克隆生长对土壤水分含量的响应. 南京林业大学学报:自然科学版, 2012, 36(04): 46-50. |
| [29] |
陈劲松, 董鸣, 于丹, 刘庆. 不同光照条件下聚花过路黄的克隆构型和分株种群特征. 应用生态学报, 2004, 15(08): 1383-1388. DOI:10.3321/j.issn:1001-9332.2004.08.017 |
| [30] |
Armstrong R A. A quantitative theory of reproductive effort in rhizomatous perennial plants. Ecology, 1982, 63(3): 679-686. DOI:10.2307/1936788 |
| [31] |
李莹, 曾晓琳, 游明鸿, 刘金平, 蔡捡. 5种川西北沙化地草本植物生态适应策略的差异性. 草业科学, 2016, 33(5): 843-850. |
| [32] |
张玉芬, 张大勇. 克隆植物的无性与有性繁殖对策. 植物生态学报, 2006, 30(1): 174-183. DOI:10.3321/j.issn:1005-264X.2006.01.024 |
| [33] |
陈玉橱, 董鸣. 毛乌素沙地根茎灌木羊柴的基株特征和不同生境中的分株种群特征. 植物生态学报, 2000, 24(1): 40-45. DOI:10.3321/j.issn:1005-264X.2000.01.008 |
| [34] |
廖明隽, 王其兵, 宋明华, 董鸣. 内蒙古锡林河流域不同生境中羊草的克隆构型和分株种群特征. 植物生态学报, 2002, 26(01): 33-38. DOI:10.3321/j.issn:1005-264X.2002.01.006 |
| [35] |
罗学刚, 董鸣. 匍匐茎草本蛇莓克隆构型对土壤养分的可塑性反应. 生态学报, 2001, 21(12): 1957-1963. DOI:10.3321/j.issn:1000-0933.2001.12.001 |
| [36] |
周天阳, 高景, 贺俊东, 薛晶月, 孙建, 王金牛, 徐波, 谢雨, 吴彦. 高山草地环山样带异质坡向上3种植物的株高、叶片性状与生物量分配. 应用与环境生物学报, 2018, 24(03): 425-433. |
| [37] |
Boulangeat I, Georges D, Dentant C, Bonet R, Van E S J, Abdulhak S, Thuiller W. Anticipating the spatio-temporal response of plant diversity and vegetation structure to climate and land use change in a protected area. Ecography, 2014, 37(12): 1230-1239. DOI:10.1111/ecog.00694 |
| [38] |
李红丽, 智颖飙, 雷光春, 赵磊, 安树青, 邓自发, 周长芳, 徐贵明. 不同水位梯度下克隆植物大米草的生长繁殖特性和生物量分配格局. 生态学报, 2009, 29(07): 3525-3531. DOI:10.3321/j.issn:1000-0933.2009.07.010 |
| [39] |
Fortunel C, Fine P V A, Baraloto C. Leaf, stem and root tissue strategies across 758 Neotropical tree species. Functional Ecology, 2012, 26(5): 1153-1161. DOI:10.1111/j.1365-2435.2012.02020.x |
| [40] |
Shi L X, Guo J X. Changes in photosynthetic and growth characteristics of Leymus chinensis community along the retrogression on the Songnen grassland in northeastern China. Photosynthetica, 2006, 44(4): 542-547. DOI:10.1007/s11099-006-0068-8 |
| [41] |
邱天, 鞠淼, 徐嘉咛, 杨允菲. 芦苇生长与物质生产对盐碱胁迫的可塑性响应. 东北师大学报:自然科学版, 2013, 45(01): 108-112. |
| [42] |
贡璐, 朱美玲, 塔西甫拉提·特依拜, 张雪妮, 韩丽, 解丽娜. 塔里木盆地南缘旱生芦苇生态特征与水盐因子关系. 生态学报, 2014, 34(10): 2509-2518. |