 2019, Vol. 39
2019, Vol. 39文章信息
- 陈春晓, 谢秀华, 王宇鹏, 李俊朋, 信健, 朱婷婷, 刘静, 陈敏
- CHEN Chunxiao, XIE Xiuhua, WANG Yupeng, LI Junpeng, XIN Jian, ZHU Tingting, LIU Jing, CHEN Min
- 盐分和干旱对沙枣幼苗生理特性的影响
- Effects of salt and drought on the physiological characteristics of Elaeagnus angustifolia L. seedlings
- 生态学报. 2019, 39(12): 4540-4550
- Acta Ecologica Sinica. 2019, 39(12): 4540-4550
- http://dx.doi.org/10.5846/stxb201807041463
-
文章历史
- 收稿日期: 2018-07-04
- 网络出版日期: 2019-04-01





2. 山东省东营市农业农村局, 东营 257091;
3. 山东省潍坊北海中学, 潍坊 261200
2. Dongying City Agricultural and Rural Bureau, Dongying 257091, China;
3. Weifang Beihai Middle School, Shandong Province, Weifang 261200, China
非生物胁迫(例如:干旱、盐碱、极端温度、氧化胁迫)是限制植物生长的主要因素, 非生物胁迫通过影响植物生长、发育、营养代谢及一系列分子变化, 进而影响植物形态、结构和产量[1]。盐和干旱是两种主要的非生物胁迫因子而且二者常常同时发生, 尤其在干旱、半干旱地区, 干旱和盐胁迫共同作用严重危害着植物的生长及产量。黄河三角洲是黄河数百年来冲积而成的一块年轻的陆地, 由于成陆较晚, 土地整体质量不高, 现有的533333 hm2边际性土地多为盐碱地, 其特点是土壤含盐量高、有机质含量低、营养元素不均衡, 地下水位高, 春秋季易旱、夏季易涝, 不适合于普通农作物生长[2]。生长在黄河三角洲地区的植物在春、秋季常常受到盐和干旱共同的影响。
正是由于黄河三角洲地区特殊的土壤情况和气候特征, 黄河三角洲地区的植物种类少、植被单一, 严重制约了当地的经济发展和生态改良。所以引种一些适合当地生长的经济和生态植物迫在眉睫。沙枣(Elaeagnus angustifolia L.)是胡颓子科Elaeagnaceae)胡颓子属(Elaeagnus Lnn.)的落叶小乔木, 主要分布于我国西北五省, 在北京、天津和山东等地也开展了引种栽培[3-4]。沙枣具有一定的耐盐性, 可以在黄河三角洲地区生长良好[2, 5]。但是对于沙枣适应黄河三角洲地区春、秋季的盐旱共同胁迫的机理是还不清楚。目前单一胁迫因子(盐胁迫或干旱胁迫)对植物生长的影响研究较多, 但对于盐分和干旱共胁迫的研究相对较少。在已有的研究中发现:许多植物在盐分或者干旱胁迫时, 其生物量是降低的, 而在盐分和干旱共胁迫胁迫时, 在一定的条件下, 植物会表现出比在单一胁迫下生长更好的现象。例如, 2012年朱金方等研究发现, 盐旱胁迫下柽柳表现出一定的交叉适应性, 适度的干旱胁迫能增强柽柳的耐盐能力[6];另外2017年翁亚伟等研究发现, 盐旱复合胁迫可以通过增强根系水分吸收及降低根叶中ABA含量以维持较高光合能力, 进而提高小麦适应性[7];2017年华智锐等研究也发现, 在盐旱共胁迫下小麦幼苗表现出交叉适应性[8]。这些研究都说明了在干旱和盐分这两种胁迫因子的共同作用下, 植物出现了交叉适应现象。
沙枣的生存能力极强, 具有抗干旱、耐盐碱、抗风沙、耐贫瘠、根蘖性强的特点, 可用于水土保持, 还可以调节气候、净化空气、改良土壤, 并且沙枣根系上有固氮根瘤菌, 能提高土壤肥力, 改善土壤环境[9]。沙枣作为耐逆的先锋树种是否可以更好的适应盐旱共胁迫?关于这个问题没有相关的报道, 所以本实验模拟黄河三角洲地区春、秋季的盐旱共同胁迫, 来研究盐分和干旱共同胁迫对沙枣幼苗生长及生理特性的影响。从而为将沙枣引进黄河三角洲地区提供理论依据。
1 材料与方法 1.1 实验材料实验材料为沙枣(Elaeagnus angustifolia L.)幼苗。本实验室于2017年从新疆吐鲁番购入沙枣果实, 将果实去果肉后得到干净的种子, 将种子种植于培养盘中获得沙枣幼苗备用。
1.2 材料的培养机械方法去除果肉, 对种子进行统一的消毒处理, 用蒸馏水反复冲洗, 备用。种植前将其置于低温(0—4℃)下进行冷层积处理(低温处理采用低温培养箱, 品牌:上海捷呈;型号:LRH-600F), 以打破休眠并软化种皮以便于种子萌发, 10 d后将种子取出(此时种子已经露白)。种植于装有洗净细沙的塑料盆中, 每盆播种20粒种子(种子距沙面约2 cm), 然后移至温室(光强:600 μmol m-2 s-1;相对湿度:70%;温度:24℃;光照:16 h光亮, 8 h黑暗)内。培养期间每天浇水保持沙子湿润, 待种子萌发后, 停止浇水改为浇1/2 Hoagland营养液。待沙枣幼苗长出2—3片真叶时, 进行间苗(保留盆内生长较健壮且生长状况大致相同的7—8株幼苗), 待幼苗生长到5—6片真叶时, 开始胁迫实验。
1.3 实验处理实验分为6个处理, 每个处理5个重复。处理方法如下:
① 对照(the control, CK):一直浇灌1/2 Hoagland营养液。
② 轻度干旱(the treatment of mild drought, LS):浇灌1/2 Hoagland营养液7 d, 之后保持土壤含水量在轻度干旱标准(土壤含水量, 7%—9%, 下同)14 d, 每天于早中晚3次测量含水量(Soil water content were determined with an HH2 moisture meter with a WET-2 sensor (Delta-T, Cambridge, UK)), 补浇1/2 Hoagland营养液, 使含水量保持不变, 下同。
③ 重度干旱(the treatment of severe drought, SS):浇灌1/2 Hoagland营养液7 d, 之后保持土壤含水量在轻度干旱标准(土壤含水量3%—5%, 下同)14 d, 每天于早中晚3次测量含水量, 补浇1/2 Hoagland营养液, 使含水量保持不变, 下同。
④ 盐(the treatment of 100 mmol/L NaCl, 100CK):一直浇灌含有100 mmol/L NaCl的1/2 Hoagland营养液。
⑤ 盐+轻度干旱(the treatment of 100 mmol/L NaCl and mild drought, 100LS):浇灌含有100 mmol/L NaCl的1/2 Hoagland营养液7 d, 之后保持土壤含水量在轻度干旱标准14 d, 每天于早中晚三次测量含水量。
⑥ 盐+重度干旱(the treatment of 100 mmol/L NaCl and severe drought, 100SS):浇灌含有100 mmol/L NaCl的1/2 Hoagland营养液7 d, 之后保持土壤含水量在重度干旱标准14 d, 每天于早中晚三次测量含水量。
1.4 生理指标的测定 1.4.1 株高、根长、含水量及根冠比的测定处理2周后, 用刻度尺测量株高(沙面以上为株高), 然后小心将整株植物取出, 洗净后用刻度尺测量根长并称量地上与地下部分的鲜重(fresh mass, FW)。将鲜材料放入105℃的烘箱中杀青10 min后, 80℃烘干至恒重, 分别称地上与地下部分干重(dry mass, DW)。根据下列方式计算组织含水量和幼苗根冠比:

|
采用高奔等的方法[10]。根据下式计算组织中叶绿素含量:
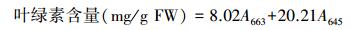
|
式中, A645为645 nm处的吸光度;A663为663 nm处的吸光度。
1.4.3 光合参数的测定采用李艳等的方法[11], 使用CIRAS-2型英国PP.Systems公司便携式光合作用测定仪对沙枣完全展开的第二叶进行净光合速率(netphotosynthetic rate, Pn)、胞间CO2浓度(intercellular CO2 concentration, Ci)、气孔导度(stomatal conductance, gs)、蒸腾速率(transpiration rate, Tr)等光合参数的测定。测定过程中光强约为1500 μmol m-2 s-1, 大气温度变化范围为20—24℃, 大气CO2浓度变化范围为477—497 μmol/L, 测定时间为10:00—11:30。
1.4.4 荧光参数的测定采用冯伟等的方法[12], 下列方式计算组织含水量和幼苗根冠比(9:30—12:00), 用OS5-FL型调制式叶绿素荧光分析仪(OPTI-SCI-ENCES, 美国)测定光适应下的最大光化学效率(maximum quantumefficiency, Fv/Fm)、光系统Ⅱ实际光化学量子效率(quantum efficiency of photosystem Ⅱ, ΦPSII)以及光化学猝灭系数(photochemical quenching coefficient, qP)。取完全展开的同一叶位的沙枣叶片测定, 测定时先固定好沙枣叶片, 使叶片受光一致, 每个处理测定至少20片叶。测定暗适应荧光参数前将叶片充分暗适应20 min。
1.4.5 无机离子含量的测定采用张志良的方法[13], 将烘干的沙枣材料分别碾碎后于马弗炉中550℃灰化, 灰分用浓硝酸溶解, 定容后用原子吸收光谱仪(日立Z-8000型)测定Na+含量、K+含量, AgNO3滴定法测定Cl-含量。
1.4.6 脯氨酸含量的测定采用酸性茚三酮法[14]。根据下式计算组织中脯氨酸含量:
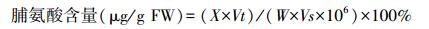
|
式中, X为从标曲中查得的脯氨酸含量(μg);Vt为提取液体积(mL);W为样品的质量(g);Vs为测定时所用样品的体积。
1.4.7 可溶性糖含量的测定采用蒽酮法[15], 称取0.2 g沙枣叶片磨细的烘干样品置10 mL离心管中, 用80%乙醇浸提离心后定容至25 mL。准确吸取糖的乙醇提取液2 mL水浴中蒸去乙醇, 准确加水10 mL搅拌使糖完全溶解。吸取上清液2 mL加入试管中, 记录620 nm下的OD值并计算可溶性糖含量。
1.4.8 总酚含量的测定采用Folin-Ciocalteu比色法[16]。根据下式计算组织中总酚含量:
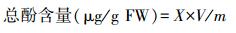
|
式中, X为由吸光度按标准曲线查的结果;V为粗提液总体积(mL);m为称样质量(g)。
1.4.9 类黄酮含量的测定采用亚硝酸钠-硝酸铝比色法[17]。根据下式计算组织中类黄酮含量:
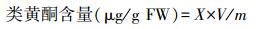
|
式中, X为由吸光度按标准曲线查的结果;V为粗提液总体积(mL);m为称样质量(g)。
1.4.10 丙二醛(MDA)含量的测定紫外-可见分光光度计法[18]。根据下式计算组织中MDA的含量:
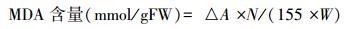
|
式中, △A为A532和A600的差;N为上清液总体积;155为1 mmol为反应产物在532 nm处的吸收系数;W为称取植物材料的鲜重(g)。
1.5 数据统计分析上述各处理5个重复, 所得数据用SigmaPlot 10.0作图, 用IBM SPSS Statistics 19软件进行显著性分析, 同一种处理的各参数以平均值加减标准差的形式表示(mean±SD), 同时采用字母标注法(P < 0.05表示存在显著性差异)。
2 结果 2.1 盐旱共胁迫对沙枣幼苗生长速率和生物量的影响轻度干旱除了降低了沙枣的株高和根长外, 对沙枣的生物量和含水量没有影响, 但是重度干旱却明显降低了沙枣的各生长指标(图 1, 表 1)。除了含水量以外, 100 mmol/L NaCl明显降低了沙枣的各生长指标。和100 mmol/L NaCl处理条件下的各项生理指标相比, 轻度干旱和100 mmol/L NaCl共胁迫对沙枣的生长没有显著影响, 但是重度干旱和100 mmol/L NaCl共胁迫却明显降低了沙枣的生长。

|
| 图 1 盐旱共胁迫对沙枣幼苗生长的影响 Fig. 1 The effects of salt and drought co-stress on the growth of E. angustifolia seedlings CK:对照, the control;LS:轻度干旱, mild drought;SS:重度干旱, severe drought;100CK:盐, 100 mmol/L NaCl;100LS:盐+轻度干旱, 100 mmol/L NaCl and mild drought co-stress;100SS:盐+重度干旱, 100 mmol/L NaCl and severe drought co-stress |
| 参量 Parameters |
处理Treatment | |||||
| CK | LS | SS | 100+CK | 100+LS | 100+SS | |
| 地上鲜重Shoot fresh weight | 1.11±0.29a | 1.11±0.30a | 0.92±0.17b | 0.88±0.31b | 0.70±0.22bc | 0.59±0.13c |
| 地上干重Shoot dry weight | 0.23±0.06a | 0.25±0.05a | 0.23±0.04b | 0.20±0.06b | 0.19±0.05bc | 0.17±0.04c |
| 地下鲜重Root fresh weight | 0.73±0.03a | 0.65±0.04ab | 0.51±0.02bc | 0.40±0.02cd | 0.35±0.02d | 0.30±0.02de |
| 地下干重Root dry weight | 0.12±0.04a | 0.11±0.04ab | 0.07±0.02b | 0.07±0.06b | 0.07±0.03b | 0.07±0.02b |
| 株高Plant length | 11.06±0.98a | 9.40±1.00b | 6.87±0.58d | 9.68±0.75b | 8.31±0.58c | 6.77±0.80d |
| 根长Root length | 8.77±0.76a | 7.36±0.55b | 6.04±0.62de | 6.92±0.63bc | 6.46±0.83cd | 5.69±0.69e |
| 地上含水量Shoot water content | 78.73±2.40a | 77.72±3.14a | 74.42±4.40b | 76.82±3.07a | 72.40±2.30c | 71.47±2.53c |
| 根冠比Root shoot ratio | 0.16±0.04a | 0.14±0.03ab | 0.11±0.03e | 0.13±0.04bc | 0.12±0.03cd | 0.09±0.02d |
| 表中数据为平均数±标准差(n=5);不同小写字母表示不同处理之间差异显著(P < 0.05); CK:对照, the control;LS:轻度干旱, mild drought;SS:重度干旱, severe drought;100+CK:盐, 100 mmol/L NaCl;100+LS:盐+轻度干旱, 100 mmol/L NaCl and mild drought co-stress;100+SS:盐+重度干旱, 100 mmol/L NaCl and severe drought co-stress | ||||||
轻度干旱对沙枣幼苗叶绿素含量没有影响, 但是重度干旱却明显降低了沙枣幼苗的叶绿素含量。100 mmol/L NaCl显著降低了沙枣幼苗的叶绿素含量(P < 0.05)。和100 mmol/L NaCl处理条件下的各项生理指标相比, 轻度干旱和100 mmol/L NaCl共胁迫对其没有显著影响(P> 0.05), 但是重度干旱和100 mmol/L NaCl共胁迫却明显降低了沙枣幼苗的叶绿素含量(P < 0.05) (图 2)。

|
| 图 2 盐旱共胁迫对沙枣幼苗叶绿素含量的影响 Fig. 2 The effects of salt and drought co-stress on the chlorophyll content of E. angustifolia seedlings 图中数据为平均数±标准差(n=5);不同小写字母表示不同处理之间差异显著(P < 0.05) |
轻度干旱除了降低了沙枣叶片净光合速率和气孔导度外, 对沙枣的胞间CO2浓度和蒸腾速率没有影响(P>0.05), 但是重度干旱却明显降低了除胞间CO2浓度外的各光合参数(P < 0.05)。除了净光合速率(降低)和胞间CO2浓度(升高)外, 100 mmol/L NaCl对沙枣的各光合参数没有影响(P>0.05)。和100 mmol/L NaCl处理条件下的各项光合参数相比, 除净光合速合以及气孔导度外, 轻度干旱和100 mmol/L NaCl共胁迫对沙枣的光合参数没有显著影响(P>0.05), 但是重度干旱和100 mmol/LNaCl共胁迫却明显降低了沙枣的净光合速率、气孔导度、蒸腾速率且明显增加了胞间CO2浓度(P < 0.05) (图 3)。

|
| 图 3 盐旱共胁迫对沙枣幼苗光合参数的影响 Fig. 3 The effects of salt and drought co-stress on photosynthesis of E. angustifolia seedlings |
轻度干旱或100 mmol/L NaCl降低了沙枣的最大光化学效率和光化学猝灭系数外, 但对沙枣的实际光化学效率没有影响(P>0.05), 但是重度干旱却明显降低了沙枣的各荧光参数(P < 0.05)。和100 mmol/L NaCl处理条件下的各荧光参数相比, 轻度干旱和100 mmol/L NaCl共胁迫对沙枣的荧光参数没有显著影响(P>0.05), 但是重度干旱和100 mmol/L NaCl共胁迫却明显降低了沙枣的荧光参数(P < 0.05) (图 4)。

|
| 图 4 盐旱共胁迫对沙枣幼苗荧光参数的影响 Fig. 4 The effects of salt and drought co-stress on fluorescence parameters of E. angustifolia seedlings |
轻度干旱和重度干旱均显著降低了沙枣的地上和地下的K+含量, 显著增加了沙枣地上、地下的Na+含量(P < 0.05)。100 mmol/L NaCl极显著降低了沙枣的地上和地下的K+含量, 极显著增加了沙枣地上、地下的Na+含量(P < 0.01)。和100 mmol/L NaCl处理条件下的沙枣离子含量相比, 轻度干旱和100 mmol/L NaCl共胁迫以及重度干旱和100 mmol/L NaCl共胁迫都显著降低了地上和地下的K+含量, 显著增加了沙枣地上、地下的Na+含量(P < 0.05) (图 5)。

|
| 图 5 盐旱共胁迫对沙枣幼苗离子含量的影响 Fig. 5 The effects of salt and drought co-stress on ionic content of E. angustifolia seedlings |
轻度干旱除了对沙枣叶片中可溶性糖、脯氨酸和总酚含量没有影响外(P>0.05), 显著提高了沙枣幼苗其他有机物质和次生代谢产物含量(P < 0.05), 但是重度干旱显著提高了各指标含量(P < 0.05)。除脯氨酸含量外, 100 mmol/L NaCl明显提高了沙枣幼苗的有机物质及次生代谢产物含量(P < 0.05)。和100 mmol/L NaCl处理条件下的各项指标相比, 轻度干旱和100 mmol/L NaCl共胁迫以及重度干旱和100 mmol/L NaCl共胁迫都显著增加了各指标含量(P < 0.05) (图 6)。

|
| 图 6 盐旱共胁迫对沙枣有机物质及次生代谢产物含量的影响 Fig. 6 The effects of salt and drought co-stress on organic matter content and secondary metabolites content of E. angustifolia seedlings |
复水一周后, 轻度干旱除了降低了沙枣幼苗的地下鲜重和干重外, 对沙枣的地上鲜重、地上鲜重增加百分比、地上干重、地上干重增加百分比、地下鲜重增加百分比、地下干重增加百分比没有影响(P>0.05), 但是重度干旱却显著降低了复水后沙枣的生物量指标(P < 0.05)。100 mmol/L NaCl显著降低了复水后沙枣幼苗的各生物量指标(P < 0.05)。和100 mmol/L NaCl处理条件下的各项生物量指标相比, 轻度干旱和100 mmol/L NaCl共胁迫除地上鲜重以及地上鲜重增加百分比外的其他生物量指标没有影响(P>0.05), 但是重度干旱和100 mmol/L NaCl共胁迫却明显降低了复水后沙枣的生长(P < 0.05) (表 2)。
| 参量 Parameters |
处理Treatment | |||||
| CK | LS | SS | 100+CK | 100+LS | 100+SS | |
| 地上鲜重Shoot fresh weight | 1.44±0.05a | 1.43±0.04a | 1.07±0.06b | 1.09±0.05b | 0.85±0.06c | 0.68±0.05d |
| 地上鲜重增加百分比 Shoot fresh weight increase percentage |
0.26±0.01a | 0.26±0.01a | 0.16±0.01d | 0.24±0.01b | 0.22±0.01bc | 0.15±0.01d |
| 地上干重Shoot dry weight | 0.30±0.02a | 0.31±0.02a | 0.27±0.01b | 0.25±0.01bc | 0.24±0.02c | 0.19±0.01d |
| 地上干重增加百分比 Shoot dry weight increase percentage |
0.28±0.01a | 0.28±0.01a | 0.17±0.01c | 0.26±0.01b | 0.25±0.01b | 0.17±0.01c |
| 地下鲜重Root fresh weight | 0.98±0.04a | 0.87±0.05b | 0.64±0.04c | 0.51±0.02d | 0.48±0.04d | 0.37±0.03e |
| 地下鲜重增加百分比 Root fresh weight increase percentage |
0.35±0.01a | 0.34±0.01a | 0.23±0.01c | 0.30±0.01b | 0.29±0.01b | 0.23±0.03c |
| 地下干重Root dry weight | 0.15±0.01a | 0.14±0.01b | 0.09±0.01cd | 0.10±0.01c | 0.09±0.01cd | 0.08±0.01d |
| 地下干重增加百分比 Root dry weight increase percentage |
0.29±0.01a | 0.29±0.01a | 0.15±0.01cd | 0.24±0.01b | 0.22±0.01bc | 0.17±0.01c |
| 表中数据为平均数±标准差(n=5);不同小写字母表示不同处理之间差异显著(P < 0.05) | ||||||
生物量是植物遭遇胁迫的综合反应, 干旱胁迫和盐分胁迫抑制植物的生长, 且胁迫程度越高抑制程度越明显[19]。对于本研究的沙枣幼苗干旱胁迫及盐胁迫均可抑制沙枣幼苗的生长、降低植物组织含水量和生物量, 尤其是重度干旱更为明显。干旱或盐处理降低沙枣幼苗的根冠比, 沙枣在轻度干旱和盐共胁迫下表现出一定的交叉适应性。
处于逆境条件下的植物, 为减少蒸腾作用关闭气孔, 同时也阻碍了CO2进入叶片, 影响了CO2参与羧化作用, 降低了植物的净光合速率[20-21];CO2同化量降低、吸收的过剩光能会引起光抑制, 严重时还会破坏光合结构, 从而使植物光合作用下降[22]。本研究发现:随着干旱程度增加, 除胞间CO2浓度外, 沙枣幼苗的其他光合参数和各荧光参数均呈现降低的趋势;盐旱共胁迫条件下, 各光合参数及荧光参数除胞间CO2浓度外其他参数也呈现降低趋势。从这些结果可以看出轻度干旱和盐共胁迫下, 沙枣幼苗在一定程度上通过减少光捕获、热耗散和酶活性调节稳定光合机构功能, 阻止光合速率的下降;重度盐旱胁迫下光系统Ⅱ和抗氧化酶系统损伤, 光合速率下降更加显著, 这种非气孔限制因素与已有的研究相一致[23]。
渗透调节物质在植物适应盐旱胁迫环境中也发挥着重要作用[24]。在盐、旱等逆境条件下, 植物为了保持水分以适应盐旱胁迫环境, 常常通过生理代谢活动增加无机离子以及渗透调节物质的方式来提高细胞液浓度、降低渗透势, 从而维持正常的细胞膨压进而使植物体内与之相关的生理活动正常进行, 提高植物的抗逆适应性[25-26]。本研究也表明沙枣幼苗在干旱、盐分以及干旱和盐分共胁迫下, 通过积累无机离子(主要是Na+)和有机溶质来保持较低的渗透势, 以增加保水能力。这与沙棘植物在盐碱胁迫下的结果一致[27]。对于沙枣植物, 积累的有机溶质主要是脯氨酸, 脯氨酸作为一种重要的渗透调节物质, 在植物的耐逆过程中发挥重要的作用, 大量研究表明脯氨酸的增加明显了提高了植物的耐逆性[28-29]。逆境条件下, 植物除了积累渗透调节物质外, 植物还会积累次生代谢产物以抵御逆境, 有利于植物体适应逆境环境[30]。本研究的沙枣幼苗在逆境条件下通过大量积累类黄酮和总酚含量来提高沙枣幼苗的耐逆性。这与蔡娜等[31]、Wang等[32]的研究结果一致。
总之, 盐分和干旱处理明显抑制了沙枣幼苗的生长, 轻度干旱和盐分共胁迫条件下, 沙枣幼苗表现出一定的交叉适应现象。沙枣幼苗通过降低生长、积累渗透调节物质、积累次生代谢产物来抵抗逆境。
| [1] |
Jia J, Huang C, Bai J H, Zhang G L, Zhao Q Q, Wen X J. Effects of drought and salt stresses on growth characteristics of euhalophyte Suaeda salsa in coastal wetlands. Physics and Chemistry of the Earth, Parts A/B/C, 2018, 103: 68-74. DOI:10.1016/j.pce.2017.01.002 |
| [2] |
Lin J, Li J P, Yuan F, Yang Z, Wang B S, Chen M. Transcriptome profiling of genes involved in photosynthesis in Elaeagnus angustifolia L. under salt stress. Photosynthetica, 2018, 56(4): 998-1009. |
| [3] |
郭丽君, 王玉涛. 沙枣种质资源特性及利用价值. 中国野生植物资源, 2008, 27(5): 32-34. DOI:10.3969/j.issn.1006-9690.2008.05.008 |
| [4] |
管文轲, 徐娜. 沙枣资源利用研究与开发现状述评. 安徽农学通报, 2012, 18(19): 119-120, 213-213. DOI:10.3969/j.issn.1007-7731.2012.19.060 |
| [5] |
Qi Y, Li J P, Chen C X, Li L X, Zheng X L, Liu J, Zhu T T, Pang C H, Wang B S, Chen M. Adaptive growth response of exotic Elaeagnus angustifolia L. to indigenous saline soil and its beneficial effects on the soil system in the Yellow River Delta, China. Trees, 2018. DOI:10.1007/s00468-018-1746-4 |
| [6] |
朱金方, 夏江宝, 陆兆华, 刘京涛, 孙景宽. 盐旱交叉胁迫对柽柳幼苗生长及生理生化特性的影响. 西北植物学报, 2012, 32(1): 124-130. DOI:10.3969/j.issn.1000-4025.2012.01.020 |
| [7] |
翁亚伟, 张磊, 张姗, 田中伟, 靳雪莹, 李梦雅, 余钟毓, 姜东, 戴廷波. 盐旱复合胁迫对小麦幼苗生长和水分吸收的影响. 生态学报, 2017, 37(7): 2244-2252. |
| [8] |
华智锐, 于浩世, 王新军. 盐旱交叉胁迫对商麦5226幼苗光合指标的影响. 云南师范大学学报:自然科学版, 2017, 37(2): 52-57. |
| [9] |
杨升, 刘涛, 张华新, 李焕勇, 张丽. 盐胁迫下沙枣幼苗的生长表现和生理特性. 福建林学院学报, 2014, 34(1): 64-70. DOI:10.3969/j.issn.1001-389X.2014.01.013 |
| [10] |
高奔, 宋杰, 刘金萍, 隋娜, 范海, 王宝山. 盐胁迫对不同生境盐地碱蓬光合及离子积累的影响. 植物生态学报, 2010, 34(6): 671-677. DOI:10.3773/j.issn.1005-264x.2010.06.006 |
| [11] |
李艳, 赵小明, 夏秀英, 栾雨时, 杜昱光, 李凤兰. 壳寡糖对干旱胁迫下油菜光合参数的影响. 作物学报, 2008, 34(2): 326-329. DOI:10.3321/j.issn:0496-3490.2008.02.023 |
| [12] |
冯伟, 李晓, 王永华, 王晨阳, 郭天财. 小麦叶绿素荧光参数叶位差异及其与植株氮含量的关系. 作物学报, 2012, 38(4): 657-664. |
| [13] |
王素平, 郭世荣, 胡晓辉, 贾永霞, 焦彦生. NaCl胁迫对黄瓜幼苗体内K+、Na+和Cl-分布的影响. 生态学杂志, 2007, 26(3): 348-354. DOI:10.3321/j.issn:1000-4890.2007.03.010 |
| [14] |
职明星, 李秀菊. 脯氨酸测定方法的改进. 河南科技学院学报:自然科学版, 2005, 33(4): 10-12. |
| [15] |
刘海英, 王华华, 崔长海, 王曼, 郭净净, 文昭普, 李安琪. 可溶性糖含量测定(蒽酮法)实验的改进. 实验室科学, 2013, 16(2): 19-20. DOI:10.3969/j.issn.1672-4305.2013.02.007 |
| [16] |
梅进, 丁文平, 熊金娟, 庄坤. Folin-Ciocalteu比色法测定大麦糟中总酚条件的优化. 食品与机械, 2016, 32(2): 51-54. |
| [17] |
郭亚健, 范莉, 王晓强, 张兰珍. 关于NaNO2—Al(NO3)3—NaOH比色法测定总黄酮方法的探讨. 药物分析杂志, 2002, 22(2): 97-99. |
| [18] |
张秋萍, 吴霞红, 郑剑恒, 孙孟炜. 生物样本中丙二醛测定方法的研究进展. 理化检验-化学分册, 2016, 52(8): 979-985. |
| [19] |
贺少轩, 梁宗锁, 蔚丽珍, 周自云. 土壤干旱对2个种源野生酸枣幼苗生长和生理特性的影响. 西北植物学报, 2009, 29(7): 1387-1393. DOI:10.3321/j.issn:1000-4025.2009.07.015 |
| [20] |
Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33: 317-345. DOI:10.1146/annurev.pp.33.060182.001533 |
| [21] |
张守仁. 叶绿素荧光动力学参数的意义及讨论. 植物学通报, 1999, 16(4): 444-448. DOI:10.3969/j.issn.1674-3466.1999.04.021 |
| [22] |
齐学礼, 胡琳, 董海滨, 张磊, 王根松, 高崇, 许为钢. 强光高温同时作用下不同小麦品种的光合特性. 作物学报, 2008, 34(12): 2196-2201. |
| [23] |
张仁和, 郑友军, 马国胜, 张兴华, 路海东, 史俊通, 薛吉全. 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响. 生态学报, 2011, 31(5): 1303-1311. |
| [24] |
荣少英, 郭蜀光, 张彤. 干旱胁迫对甜高粱幼苗渗透调节物质的影响. 河南农业科学, 2011, 40(4): 56-59. DOI:10.3969/j.issn.1004-3268.2011.04.015 |
| [25] |
刘艳, 陈贵林, 蔡贵芳, 张中霞, 岳鑫. 干旱胁迫对甘草幼苗生长和渗透调节物质含量的影响. 西北植物学报, 2011, 31(11): 2259-2264. |
| [26] |
刘建新, 王金成, 王瑞娟, 贾海燕. 旱盐交叉胁迫对燕麦幼苗生长和渗透调节物质的影响. 水土保持学报, 2012, 26(3): 244-248. |
| [27] |
Yang C W, Shi D C, Wang D L. Comparative effects of salt and alkali stresses on growth, osmotic adjustment and ionic balance of an alkali-resistant halophyte Suaeda glauca (Bge.). Plant Growth Regulation, 2008, 56(2): 179-190. DOI:10.1007/s10725-008-9299-y |
| [28] |
史玉炜, 王燕凌, 李文兵, 高述民, 李霞. 水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响. 新疆农业大学学报, 2007, 30(2): 5-8. DOI:10.3969/j.issn.1007-8614.2007.02.002 |
| [29] |
Sperdouli I, Moustakas M. Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsis thaliana to drought stress. Journal of Plant Physiology, 2012, 169(6): 577-585. DOI:10.1016/j.jplph.2011.12.015 |
| [30] |
Foyer C H, Lelandais M, Kunert K J. Photooxidative stress in plants. Physiologia Plantarum, 1994, 92(4): 696-717. DOI:10.1111/ppl.1994.92.issue-4 |
| [31] |
蔡娜, 淡荣, 陈鹏. 水分胁迫对苦荞幼苗黄酮类物质含量的影响. 西北农业学报, 2008, 17(4): 91-93. DOI:10.3969/j.issn.1004-1389.2008.04.020 |
| [32] |
Wang D H, Du F, Liu H Y, Liang Z S. Drought stress increases iridoid glycosides biosynthesis in the roots of Scrophularia ningpoensis seedlings. Journal of Medicinal Plants Research, 2010, 4(24): 2691-2699. DOI:10.5897/JMPR |