 2019, Vol. 39
2019, Vol. 39文章信息
- 宗宁, 石培礼
- ZONG Ning, SHI Peili
- 模拟增温对西藏高原高寒草甸土壤供氮潜力的影响
- Effects of simulated warming on soil nitrogen supply potential in an alpine meadow on the Tibetan Plateau
- 生态学报. 2019, 39(12): 4356-4365
- Acta Ecologica Sinica. 2019, 39(12): 4356-4365
- http://dx.doi.org/10.5846/stxb201802260388
-
文章历史
- 收稿日期: 2018-02-26
- 网络出版日期: 2019-04-01

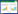




2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
全球变暖是目前面临的重要环境问题。IPCC第五次评估报告指出, 在全球温度升高的同时也存在明显的季节差异性:非生长季升温显著高于生长季的非对称趋势, 在高纬度和高海拔地区更是如此[1]。同时, 在未来气候变化情景下, 这种季节非对称性升温趋势会更加显著[2-3]。虽然目前针对增温对陆地生态系统影响已有诸多研究, 但主要集中于全年增温对生态系统群落组成[4-5]、碳氮循环[6-7]、生产力[5, 8-9]、物候[10-11]的影响等的研究, 季节非对称性增温对生态系统影响的研究却鲜有开展[12], 这在高海拔、高纬度地区忽略了季节非对称性升温的事实, 故难以预测未来增温情景对高寒生态系统的影响。
氮素是限制生态系统初级生产的最重要元素之一, 土壤氮素的矿化与周转是反映土壤供氮潜力的重要指标。土壤微生物量作为土壤“矿化-固持”过程中易转化氮的存在形式, 对土壤供氮潜力有重要的影响。已有的诸多研究表明气候变暖会显著影响生态系统氮素循环过程:在水分不亏缺的情况下, 增温会促进土壤氮矿化速率[13-15], 在生长季节会提高土壤氮的可利用性满足植物生长发育的需求。利用控制实验, Auyeung等[16]研究了增温和干旱对土壤氮素周转的影响, 增温和干旱均显著降低氮矿化速率对温度的敏感性, 但在湿润环境中增温对氮矿化速率温度敏感性的影响最大, 说明水分条件对增温的影响有重要的调节作用。冬季一般被认为是植物的休眠季节。但相关研究表明, 许多影响植物生长季节生理生态活动的过程如土壤矿化作用、养分淋溶、土壤物理性质以及温室气体排放等过程都与上个冬季降雪与土壤冰冻厚度有关[3, 17-18]。在高寒生态系统, 由于积雪或冰冻层的绝热作用, 土壤中微生物代谢活动仍比较活跃, 且能利用土壤可溶性有机碳[17, 19], 这对接下来生长季植物生长发育及碳、氮循环都会产生重要影响[19-20]。在冬季多数植物处于休眠状态, 冬季增温以及伴随的冻融交替过程的增加会增加土壤氮的损失[21]。Turner和Henry[21]利用原位培养试验研究发现, 全年增温对土壤净氮矿化速率和淋溶损失影响很小, 而冬季增温处理中生长季土壤净氮矿化速率是对照及全年增温处理的2倍。由此可见, 冬季增温对土壤氮周转速率影响更大, 且对土壤氮周转的影响具有延迟效应:不仅对增温期间氮循环过程产生影响, 还会影响随后生长季甚至更长时间尺度的氮循环过程[12, 22], 进而影响土壤供氮潜力, 但其中机理尚不明确。
作为世界上海拔最高的地区, 青藏高原高寒生态系统对全球变化响应敏感, 且呈现出季节更不均衡的增温特征, 已有报道显示, 青藏高原冬季增温幅度显著高于全年与生长季[23-25]。受高海拔影响, 低温限制了高寒草甸土壤氮矿化速率, 导致土壤氮素可利用性较低, 成为限制高寒草甸初级生产的最重要元素之一[26-27]。高寒草地占据了青藏高原面积的60%以上, 是维持高原地区生产力和畜牧业发展的基础。近几十年来, 全球环境变化已经对青藏高原高寒生态系统碳、氮循环产生了重要影响[28]。我们前期的研究发现, 经历冬季增温后群落生产和碳排放量均显著降低[29], 并显著改变群落盖度和结构组成, 导致建群种高山嵩草盖度显著降低[30], 这可能与增温导致的养分循环过程的改变有关。本研究以西藏高原高山嵩草(Kobresia pygeama)草甸为研究对象, 设置长期模拟增温样地, 对比研究全年增温和冬季增温对高寒草甸土壤氮过程的影响, 为青藏高原高寒草甸的管理提供科学指导。
1 研究区概况与实验方法 1.1 研究区概况试验布置在西藏拉萨市当雄县草原站(91°05′ E, 30°29′ N, 海拔4333 m), 地处念青唐古拉山南缘, 属于丘间盆地地形。研究区气候属于大陆性高原季风气候, 太阳辐射强、气温低、日较差大、年较差小[31]。多年平均气温1.3℃, 最冷月(1月)平均气温-10.4℃, 最热月(7月)平均气温10.7℃, 气温年较差为21.0℃。冰冻期一般从11月持续至次年1月。多年年均降水量476.8 mm, 其中约有85%集中在6—8月。年蒸发量1726.7 mm, 年平均湿润系数0.28, 介于半湿润半干旱之间[31-32]。
研究区优势植被是以高山嵩草为优势种的草原化高寒草甸, 建群种有高山嵩草、丝颖针茅(Stipa capillace)、窄叶苔草(Carex montis-everestii)等, 并伴生木根香青(Anaphalis xylorhiza)、二裂委陵菜(Potentilla bifurca)等杂类草[31-32]。由于地处湿润区向半干旱区过渡地带, 故该地区高寒草甸中丝颖针茅所占比例较高, 具有草原化草甸的特征。本地区土壤属于高寒草甸土, 结构为砂壤, 土壤有机质含量较为丰富, 但活性养分含量低。土壤厚度一般为0.3—0.5 m, 下层砾石含量较高, 详细土壤理化特征参照文献Zong等[32]。
1.2 实验设计模拟增温样地布设于2010年7月初, 样地所在区域植被均一, 属天然植被, 群落总盖度约为30%—50%。试验包含3种处理:对照(no warming, NW)、冬季增温(warming only in winter, WW)、全年增温(year-round warming, YW), 每种增温处理5个重复。
增温采用国际冻原研究计划(ITEX)通用的开顶箱(OTCs)法, OTCs规格的规格为底部直径1.5 m, 上口直径1.0 m, 高度0.4 m。材料为3 mm的聚碳酸酯, 该材料可见光和紫外线的透射率约为90%, 而近红外光线透射率小于5%[33]。2012年9月底开始冬季增温处理, 至2013年5月初植物开始返青前撤掉增温装置, 2013年9月底再次安装冬季增温装置, 2014年5月初撤掉增温装置[30]。样地内安装HOBO气象站(Onset Inc., Bourne, MA, USA), 在增温处理与非增温处理中分别埋设温度和土壤湿度探头, 用于观测增温处理对地表空气温度、0—5 cm土壤温度和湿度的影响, 数据记录频度为15 min, 日尺度降水数据来自距离本研究站点约4 km的西藏当雄县气象站(91°06′ E, 30°29′ N, 海拔4288 m), 从中国气象科学数据共享服务网(http://cdc.cma.gov.cn)下载。
1.3 样品处理与分析表层土壤(0—15 cm)样品用土钻(直径3.8 cm)约每月获取一次。将每次取回的新鲜土壤样品用镊子剔除大的植物根系与石块, 再过2 mm土壤筛除去细根并充分混匀, 用KCl(1.0 mol/L)溶液浸提(土水比为1:5), 并用全自动连续流动分析仪(AA3, Seal, Germany)测定其中的无机氮(包括铵态氮(NH4+-N)和硝态氮(NO3--N))含量。
在每个实验小区内随机选取2个样点, 齐地面剪掉植物地上部分并去除地表的凋落物, 分别埋入2个PVC管(长度12 cm, 管口直径5 cm)至10 cm刻线。随机选择其中一个用保鲜膜封口, 并用橡皮筋和胶布捆住, 以防降水带来的氮素输入和对管内氮素的淋洗, 并能保持适度的透气性[34]。同时, 把成对的另一个PVC管取出, 将其中的土壤装入自封袋, 冷藏带回实验室, 测定其NH4+-N和NO3--N的含量, 作为本阶段培养土壤无机氮的初始值。每次培养时间为20—30 d, 2012和2014年生长季分别培养3次, 2013年生长季培养4次。培养结束后将封口的PVC管内土壤取出, 处理方法同培养前土壤样品。
土壤净氮矿化速率采用下面公式进行计算:
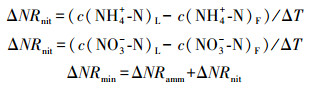
|
式中:ΔNRamm、ΔNRnit和ΔNRmin分别表示净氨化速率、净硝化速率和净矿化速率, 单位均为mg kg-1 d-1;ΔT表示培养的时间间隔(d);c(NH4+-N)L和c(NO3--N)L分别表示培养后土壤中铵态氮和硝态氮含量, c(NH4+-N)F和c(NO3--N)F分别表示培养前土壤铵态氮和硝态氮含量, 单位均为mg/kg。
将新鲜土壤过2 mm土壤筛, 去除植物根系和石块, 冷冻于冰箱中。土壤微生物量碳(Soil microbial biomass C, SMBC)测定方法如下:采用氯仿熏蒸-硫酸钾(K2SO4)浸提-总有机碳(TOC)分析测定土壤微生物量碳[35]。将20 g新鲜土样用纯化后的氯仿熏蒸24 h后, 与未熏蒸土样采用0.5 mol/L K2SO4溶液(土水比为1:5)进行浸提, 抽滤通过0.45 μm滤膜后用TOC自动分析仪(Elementar Liqui TOC, Elementar Co., Hanau, Germany)测定浸提液中溶解性有机碳含量(DOC)。土壤微生物量氮(Soil microbial biomass N, SMBN)的测定采用氯仿熏蒸-浸提-比色法进行, 熏蒸、浸提方法与SMBC一致, 浸提液中全氮的测定方法采用过硫酸钾(K2S2O8)氧化-紫外分光光度计比色法。土壤微生物量碳计算方法如下:
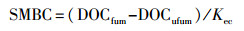
|
式中, DOCfum和DOCufum分别表示熏蒸与未熏蒸溶解性有机碳含量, Kec为校正系数0.45[36]。土壤微生物量氮(Soil microbial biomass N, SMBN)计算公式与上式相同。
1.4 数据计算与统计分析对于2012、2013和2014年土壤无机氮含量、净氮矿化速率、微生物碳、氮含量, 以增温处理为影响因素, 采用单因素方差分析方法进行统计分析。采用回归分析的方法分析土壤含水量与土壤净氮矿化速率之间的关系, 并分析寒冷季节土壤微生物量与接下来生长季初期土壤无机氮含量的关系。所有统计分析采用SPSS 16.0(SPSS for Windows, Version 16.0, Chicago, USA), 显著性水平为P < 0.05。所有图件采用Origin Pro 8.0(OriginLab Corporation, Northampton, MA, USA)制作。
2 结果与分析 2.1 增温对小气候的影响开顶箱增温装置显著提高了地表空气温度和表层土壤(0—5 cm)温度, 降低了表层土壤含水量(图 1)。2012年6月到2014年12月期间, 全年增温处理平均提高了1.6℃空气温度, 1.4℃土壤温度, 将土壤含水量降低了4.7%(体积分数)。冬季增温期间, 空气和土壤温度分别提高了1.7℃和1.6℃的, 土壤湿度降低了4.8%(体积分数)。

|
| 图 1 2012年6月—2014年12月全年增温与不增温处理地表空气温度、0—5 cm土壤温度、0—5 cm土壤湿度、自然降雨量的月尺度动态变化 Fig. 1 Seasonal dynamic of meteorological factors in year-round warming and no warming treatments, along with rainfall distribution from June 2012 to December 2014 |
全年降雨主要集中在6—9月之间, 其中2012、2013和2014年6—9月降雨量分别是312.8、332.5、431.9 mm, 分别占全年降雨量的86%、83%和94%(图 1)。2012年10月—2013年5月和2013年10月—2014年5月, 冬季增温期间降雨量分别是66.4 mm和105.8 mm(图 1)。
2.2 增温对土壤无机氮含量的影响生长季初期, 不管冬季增温还是全年增温, 土壤无机氮含量均显著低于对照处理(图 2)。与对照相比较, 2012、2013、2014年生长季初期全年增温降低了61%、40%和60%的土壤无机氮, 而2013和2014年冬季增温降低了65%和25%的土壤无机氮。而在生长季中后期, 增温对土壤无机氮含量无显著影响(图 2)。

|
| 图 2 不同施氮梯度下增温对高寒草甸土壤无机氮含量的影响 Fig. 2 Effects of warming treatments on the variations of soil inorganic N content 同一时期不同小写字母代表不同增温处理间差异显著(P < 0.05);NS代表增温处理间无显著差异;图中数据代表均值±标准差 |
土壤氮矿化作用存在年际间的变异。2012年, 生长季前期(7月)和生长季中后期(8月下旬)增温促进土壤净氮矿化作用, 而在生长季旺盛期(8月上旬)两者之间无显著差异(图 3)。2013和2014年, 生长季初期(6月)全年增温显著降低土壤净氮矿化作用, 而冬季增温无显著影响(图 3)。2013年生长中期(7月和8月), 全年增温显著促进土壤净氮矿化作用(图 3)。2014年中后期(8月)全年增温也显著促进土壤净氮矿化作用。三年的培养试验均表明生长旺盛期土壤净氮矿化表现为负值(图 3), 这说明此时土壤氮净固持大于净矿化作用, 对植物生长的供氮能力很低。

|
| 图 3 不同增温处理对2012—2014年土壤净氮矿化速率的影响 Fig. 3 Effects of warming treatments on the variations of soil net N mineralization during growing seasons in 2012, 2013 and 2014 |
2012年生长季, 土壤微生物量碳、氮表现出一致的规律。与对照相比较, 生长季初期(6月)全年增温显著降低土壤微生物量碳、氮, 旺盛期(8月)增温处理无显著影响, 而生长末期(9月)增温显著提高土壤微生物量碳、氮(图 4)。2013年, 生长季初期(6月)冬季增温显著降低土壤微生物量碳、氮, 而全年增温只降低土壤微生物量碳, 冬季(11月份)全年和冬季增温均显著提高土壤微生物量碳、氮(图 4)。

|
| 图 4 2012—2013年增温处理对土壤微生物量碳、氮的影响 Fig. 4 Effects of warming on soil microbial biomass C (SMBC) and N (SMBN) in 2012 and 2013 |
土壤含水量与净氮矿化速率呈抛物线关系(图 5)。当土壤含水量低于14%时, 土壤净氮矿化速率随土壤含水量的增加呈现逐渐增加趋势;而当土壤含水量高于14%时, 土壤净氮矿化速率随土壤含水量的增加逐渐降低, 这说明过高的土壤水分含量不利于土壤氮矿化。同时相关分析发现, 寒冷季节土壤微生物量与生长季初期土壤无机氮含量呈显著负相关关系(图 5)。

|
| 图 5 土壤含水量对氮矿化速率的影响, 以及寒冷季节土壤微生物量碳与生长季初期无机氮含量的关系 Fig. 5 Effects of soil water content on N mineralization rate, and the relationship between soil microbial biomass C (SMBC) in late growing seasons and soil inorganic N content in the following early growing seasons |
高寒草甸生态系统占据青藏高原的35%以上, 因其高海拔、温度低等因素, 植物生存环境条件严酷, 各生态因子常常处于植物生存的阈值边缘, 土壤养分矿化速率较慢, 植物生长受到有效氮的限制[27]。故气候变化导致的土壤养分变化会对高寒生态系统过程产生重要影响。我们的研究发现生长季初期, 增温显著降低土壤无机氮含量, 而在生长季中后期增温无显著影响, 这与已有一些研究结果不一致。一般来讲, 在水分不受限的情况下增温会促进土壤氮矿化速率, 在生长季节会提高土壤氮的可利用性满足植物生长发育的需求[13-15]。整合分析发现, 增温提高了52.2%的土壤净矿化速率和32.2%的净硝化速率, 与森林生态系统相比草原和灌丛对增温的响应更弱, 主要是由于增温导致的土壤含水量的降低部分抵消了增温的影响[13]。利用控制实验, Auyeung等[16]通过增温与干旱对土壤氮素周转温度敏感性研究发现, 水分状况调控增温与干旱对氮矿化速率温度敏感性的影响。由于位于半湿润区向半干旱区的过渡地带, 土壤水分是限制高寒嵩草草甸生态系统生产与土壤养分动态的重要环境因子[37-38], 而土壤水分主要受自然降雨的调控。由土壤含水量与氮矿化速率的相关关系看出, 当低于最适含水量时,随着含水量增加氮矿化速率逐渐提高(图 5)。在生长季初期降雨较少的季节, 增温导致含水量降低, 故土壤供氮能力较低;而伴随着降雨量的增加, 土壤含水量逐渐增加, 温度升高会促进氮矿化速率, 从而补偿了生长季初期土壤无机氮含量的降低。
增温会影响土壤微生物量和酶活性等过程。我们研究发现, 与对照相比较, 生长季初期全年增温显著降低土壤微生物量碳、氮, 旺盛期增温处理无显著影响, 而生长末期增温显著提高土壤微生物量碳、氮(图 4)。这与土壤无机氮的季节动态基本一致。土壤微生物量碳、氮是土壤有机碳、氮中较为活跃的组分之一, 土壤酶是土壤中具有生物活性的蛋白质, 通过对土壤中有机物质产生的生物转化过程, 使生态系统各组分间相互关联[39]。温度可以直接影响到土壤微生物量和土壤酶活性, 同时温度升高会通过影响土壤微生物群落组成、生物量等过程间接影响土壤酶活性。
3.2 冬季增温对高寒生态系统供氮潜力及生态过程的影响未来全球变暖背景下, 高纬度和高海拔地区增温更加明显, 但在这些地区冬季增温幅度最大[23-24], 并且这种季节非对称性增温趋势在未来会更加显著[2]。这种增温趋势在研究区所在的西藏当雄县得到验证:1963—1990年, 年均温、生长季(5—9月)和非生长季(10月到次年4月)均温分别以0.35、0.16℃/10a和0.49℃/10a的速率升高;1991—2010年, 年均温、生长季均温和非生长季均温分别以0.91、0.55℃/10a和1.15℃/10a的速率升高[40]。非生长季一般被认为是植物休眠期, 但已有研究表明许多影响生长季植物生理生态活动的过程, 如土壤矿化作用、养分淋溶、物理性质以及温室气体排放等过程, 都与上个冬季雪被覆盖及土壤冰冻厚度有关[17-18]。在高寒生态系统, 由于积雪覆盖或冰冻层的绝热作用, 土壤微生物代谢活动活跃, 能利用土壤可溶性有机碳[17, 19], 这对接下来生长季植物生长发育及碳、氮循环都会产生重要影响[19-20]。
研究发现, 土壤微生物量碳、氮表现出生长季较低, 而生长季末期和非生长季高的特征(图 4), 这与已有在高寒生态系统的研究呈现一致的规律。已有研究发现, 在养分受限的高寒生态系统中, 为减少对有效养分的竞争, 植物与土壤微生物对氮素的利用表现出季节分化的特征[41-42]。植物主要在生长季节吸收氮素供给生长发育, 而土壤微生物的氮固持主要发生在寒冷的时期(晚秋、冬季等), 此时土壤微生物将大量的氮素储存在体内[42-43]。冬春转换时期嗜冷微生物的死亡、底物可利用性的降低、冻融交替的破坏作用以及渗透压的变化会导致氮素从微生物体内被释放出来[44], 供植物返青和生长季利用[43, 45-46]。在自然条件下, 冬春转换时期的养分脉冲式释放是高寒生态系统在年际上最大的养分输入过程[44, 47]。同时我们研究也发现, 寒冷季节土壤微生物量与生长季初期无机氮含量呈显著负相关关系(图 5), 而在寒冷季节冬季增温导致土壤微生物量碳、氮显著高于对照处理(图 4), 这就解释了生长季初期增温处理中土壤无机氮含量低于对照处理。
土壤冻融交替是高海拔地区常见的自然现象, 早春冻融交替时期的气温变化是影响土壤氮素转化过程和氮素流失的重要因素之一[17, 48]。冻融交替过程通过影响土壤理化性质和微生物活动而影响土壤中氮素转化及生态系统养分流失[17, 49]。在全球变暖情景下, 尤其冬季增温的短期效应是导致土壤冻融交替时间提前, 增加冻融交替的频率[21, 50], 进而影响该时期生态系统碳、氮循环[2, 17]。也有研究报道, 冬季增温对生态系统的影响具有后续效应:不仅对增温期间氮素循环过程产生影响, 还会影响随后生长季甚至更长时间尺度的氮循环过程[12, 21, 51]。冬季增温导致的寒冷季节土壤微生物量较高(图 4), 以及冻融交替活动的增加, 会使得冻融交替时期土壤养分释放过多, 此时植物尚未生长发育, 故会使得更多的活动养分从生态系统中以气体或淋溶的形式流失, 导致生长季初期土壤养分的降低。土壤养分含量的降低会影响接下来生长季群落的生产、结构组成与碳氮循环等过程, 对生态系统过程产生深远的影响。
4 结论本研究量化了增温对高寒草甸土壤氮周转及其供氮潜力的影响, 并探讨了冬季增温与全年增温影响的不同。我们发现, 经历冬季增温后的生长季前期, 增温处理中土壤无机氮含量显著降低, 在植物不活跃的非生长季进行增温, 氮素不仅不会被植物吸收利用, 而可能会通过离子淋溶或气体形式从生态系统中损失, 冬季增温导致的土壤养分含量变化会影响随后生长季植物群落的生产力、结构组成与碳氮循环等过程, 对生态系统过程产生深远的影响。在生长季节, 土壤氮的周转速率受土壤水分的调控, 14%土壤含水量是土壤氮周转速率的阈值。在降雨较少的季节, 全年增温引起的土壤含水量的降低会限制土壤氮周转。由此我们推断, 未来气候变化情景下降雨格局的改变会对土壤供氮潜力产生重要影响, 进而影响植物群落的生产力、结构组成与碳氮循环等其他生态过程。
| [1] |
IPCC. Climate Change 2013:the Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2014.
|
| [2] |
Kreyling J. Winter climate change:a critical factor for temperate vegetation performance. Ecology, 2010, 91(7): 1939-1948. DOI:10.1890/09-1160.1 |
| [3] |
Kreyling J, Jurasinski G, Grant K, Retzer V, Jentsch A, Beierkuhnlein C. Winter warming pulses affect the development of planted temperate grassland and dwarfshrub heath communities. Plant Ecology & Diversity, 2011, 4(1): 13-21. |
| [4] |
Klein J A, Harte J, Zhao X Q. Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecology Letters, 2004, 7(12): 1170-1179. DOI:10.1111/ele.2004.7.issue-12 |
| [5] |
Wang S P, Duan J C, Xu G P, Wang Y F, Zhang Z H, Rui Y C, Luo C Y, Xu B, Zhu X X, Chang X F, Cui X Y, Niu H S, Zhao X Q, Wang W Y. Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology, 2012, 93(11): 2365-2376. DOI:10.1890/11-1408.1 |
| [6] |
Fu G, Shen Z X, Zhang X Z, Zhou Y T, Zhang Y J. Response of microbial biomass to grazing in an alpine meadow along an elevation gradient on the Tibetan Plateau. European Journal of Soil Biology, 2012, 52: 27-29. DOI:10.1016/j.ejsobi.2012.05.004 |
| [7] |
Luo C Y, Xu G P, Wang Y F, Wang S P, Lin X W, Hu Y G, Zhang Z H, Chang X F, Duan J C, Su A L, Zhao X Q. Effects of grazing and experimental warming on DOC concentrations in the soil solution on the QinghaiTibet plateau. Soil Biology and Biochemistry, 2009, 41(12): 2493-2500. DOI:10.1016/j.soilbio.2009.09.006 |
| [8] |
Klein J A, Harte J, Zhao X Q. Decline in medicinal and forage species with warming is mediated by plant traits on the Tibetan Plateau. Ecosystems, 2008, 11(5): 775-789. DOI:10.1007/s10021-008-9160-1 |
| [9] |
周华坤, 周兴民, 赵新全. 模拟增温效应对矮嵩草草甸影响的初步研究. 植物生态学报, 2000, 24(5): 547-553. |
| [10] |
Dorji T, Totland Ø, Moe S R, Hopping K A, Pan J B, Klein J A. Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet. Global Change Biology, 2013, 19(2): 459-472. DOI:10.1111/gcb.2012.19.issue-2 |
| [11] |
Zhu J T, Zhang Y J, Wang W F. Interactions between warming and soil moisture increase overlap in reproductive phenology among species in an alpine meadow. Biology Letters, 2016, 12(7): 20150749. DOI:10.1098/rsbl.2015.0749 |
| [12] |
Mori A S, Fujii S, Kurokawa H. Ecological consequences through responses of plant and soil communities to changing winter climate. Ecological Research, 2014, 29(4): 547-559. DOI:10.1007/s11284-013-1091-4 |
| [13] |
Bai E, Li S L, Xu W H, Li W, Dai W W, Jiang P. A metaanalysis of experimental warming effects on terrestrial nitrogen pools and dynamics. New Phytologist, 2013, 199(2): 441-451. DOI:10.1111/nph.12252 |
| [14] |
Contosta A R, Frey S D, Cooper A B. Seasonal dynamics of soil respiration and N mineralization in chronically warmed and fertilized soils. Ecosphere, 2011, 2(3): 1-21. |
| [15] |
Rustad L E, Campbell J L, Marion G M, Norby R J, Mitchell M J, Hartley A E, Cornelissen J H C, Gurevitch J, GCTENEWS. A metaanalysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia, 2001, 126(4): 543-562. DOI:10.1007/s004420000544 |
| [16] |
Auyeung D S N, Suseela V, Dukes J S. Warming and drought reduce temperature sensitivity of nitrogen transformations. Global Change Biology, 2013, 19(2): 662-676. DOI:10.1111/gcb.2012.19.issue-2 |
| [17] |
Brooks P D, Grogan P, Templer P H, Groffman P, Öquist M G, Schimel J. Carbon and nitrogen cycling in snowcovered environments. Geography Compass, 2011, 5(9): 682-699. DOI:10.1111/j.1749-8198.2011.00420.x |
| [18] |
Edwards A C, Scalenghe R, Freppaz M. Changes in the seasonal snow cover of alpine regions and its effect on soil processes:a review. Quaternary International, 2007, 162-163: 172-181. DOI:10.1016/j.quaint.2006.10.027 |
| [19] |
Brooks P D, Williams M W, Schmidt S K. Microbial activity under alpine snowpacks, Niwot Ridge, Colorado. Biogeochemistry, 1996, 32(2): 93-113. |
| [20] |
Christiansen C T, Svendsen S H, Schmidt N M, Michelsen A. High arctic heath soil respiration and biogeochemical dynamics during summer and autumn freezeineffects of longterm enhanced water and nutrient supply. Global Change Biology, 2012, 18(10): 3224-3236. DOI:10.1111/j.1365-2486.2012.02770.x |
| [21] |
Turner M M, Henry H A L. Net nitrogen mineralization and leaching in response to warming and nitrogen deposition in a temperate old field:the importance of winter temperature. Oecologia, 2010, 162(1): 227-236. DOI:10.1007/s00442-009-1435-5 |
| [22] |
Turner M M, Henry H A L. Interactive effects of warming and increased nitrogen deposition on 15N tracer retention in a temperate old field:seasonal trends. Global Change Biology, 2009, 15(12): 2885-2893. DOI:10.1111/gcb.2009.15.issue-12 |
| [23] |
Li L, Yang S, Wang Z Y, Zhu X D, Tang H Y. Evidence of warming and wetting climate over the QinghaiTibet Plateau. Arctic, Antarctic, and Alpine Research, 2010, 42(4): 449-457. DOI:10.1657/1938-4246-42.4.449 |
| [24] |
Liu X D, Chen B D. Climatic warming in the Tibetan Plateau during recent decades. International Journal of Climatology, 2000, 20(14): 1729-1742. DOI:10.1002/(ISSN)1097-0088 |
| [25] |
杜军, 杨志刚, 石磊, 马鹏飞. 近50年西藏冷暖冬的气候变化特征. 地理学报, 2011, 66(7): 885-894. |
| [26] |
Bowman W D, Theodose T A, Schardt J C, Conant R T. Constraints of nutrient availability on primary production in two alpine tundra communities. Ecology, 1993, 74(7): 2085-2097. DOI:10.2307/1940854 |
| [27] |
曹广民, 张金霞. 中国嵩草草甸. 北京: 科学出版社, 2001.
|
| [28] |
Chen H, Zhu Q A, Peng C H, Wu N, Wang Y F, Fang X Q, Gao Y H, Zhu D, Yang G, Tian J Q, Kang X M, Piao S L, Ouyang H, Xiang W H, Luo Z B, Jiang H, Song X Z, Zhang Y, Yu G R, Zhao X Q, Gong P, Yao T D, Wu J H. The impacts of climate change and human activities on biogeochemical cycles on the QinghaiTibetan Plateau. Global Change Biology, 2013, 19(10): 2940-2955. DOI:10.1111/gcb.2013.19.issue-10 |
| [29] |
宗宁, 段呈, 耿守保, 柴曦, 石培礼, 何永涛. 增温施氮对高寒草甸生产力及生物量分配的影响. 应用生态学报, 2018, 29(1): 59-67. |
| [30] |
宗宁, 柴曦, 石培礼, 蒋婧, 牛犇, 张宪洲, 何永涛. 藏北高寒草甸群落结构与物种组成对增温与施氮的响应. 应用生态学报, 2016, 27(12): 3739-3748. |
| [31] |
Shi P L, Sun X M, Xu L L, Zhang X Z, He Y T, Zhang D Q, Yu G R. Net ecosystem CO2 exchange and controlling factors in a steppe-Kobresia meadow on the Tibetan Plateau. Science in China Series D:Earth Sciences, 2006, 49(S2): 207-218. DOI:10.1007/s11430-006-8207-4 |
| [32] |
Zong N, Song M H, Shi P L, Jiang J, Zhang X Z, Shen Z X. Timing patterns of nitrogen application alter plant production and CO2 efflux in an alpine meadow on the Tibetan Plateau, China. Pedobiologia, 2014, 57(4/6): 263-269. |
| [33] |
De Frenne P, De Schrijver A, Graae B J, Gruwez R, Tack W, Vandelook F, Hermy M, Verheyen K. The use of opentop chambers in forests for evaluating warming effects on herbaceous understorey plants. Ecological Research, 2010, 25(1): 163-171. DOI:10.1007/s11284-009-0640-3 |
| [34] |
周才平, 欧阳华. 温度和湿度对长白山两种林型下土壤氮矿化的影响. 应用生态学报, 2001, 12(4): 505-508. |
| [35] |
Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703-707. DOI:10.1016/0038-0717(87)90052-6 |
| [36] |
Xu Z F, Hu R, Xiong P, Wan C, Cao G, Liu Q. Initial soil responses to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China:nutrient availabilities, microbial properties and enzyme activities. Applied Soil Ecology, 2010, 46(2): 291-299. DOI:10.1016/j.apsoil.2010.07.005 |
| [37] |
Chai X, Shi P L, Zong N, He Y T, Zhang X Z, Xu M J, Zhang J. A growing season climatic index to simulate gross primary productivity and carbon budget in a Tibetan alpine meadow. Ecological Indicators, 2017, 81: 285-294. DOI:10.1016/j.ecolind.2017.05.064 |
| [38] |
Shen Z X, Li Y L, Fu G. Response of soil respiration to shortterm experimental warming and precipitation pulses over the growing season in an alpine meadow on the Northern Tibet. Applied Soil Ecology, 2015, 90: 35-40. DOI:10.1016/j.apsoil.2015.01.015 |
| [39] |
李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895-905. |
| [40] |
王忠.念青唐古拉山南坡高寒草甸生产力沿海拔梯度的变化机理研究[D].北京: 中国科学院大学, 2011.
|
| [41] |
Bardgett R D, Bowman W D, Kaufmann R, Schmidt S K. A temporal approach to linking aboveground and belowground ecology. Trends in Ecology & Evolution, 2005, 20(11): 634-641. |
| [42] |
Jaeger Ⅲ C H, Monson R K, Fisk M C, Schmidt S K. Seasonal partitioning of nitrogen by plants and soil microorganisms in an alpine ecosystem. Ecology, 1999, 80(6): 1883-1891. DOI:10.2307/176666 |
| [43] |
Lipson D A, Schmidt S K, Monson R K. Links between microbial population dynamics and nitrogen availability in an alpine ecosystem. Ecology, 1999, 80(5): 1623-1631. DOI:10.1890/0012-9658(1999)080[1623:LBMPDA]2.0.CO;2 |
| [44] |
Jefferies R L, Walker N A, Edwards K A, Dainty J. Is the decline of soil microbial biomass in late winter coupled to changes in the physical state of cold soils?. Soil Biology and Biochemistry, 2010, 42(2): 129-135. DOI:10.1016/j.soilbio.2009.10.008 |
| [45] |
Edwards K A, McCulloch J, Kershaw G P, Jefferies R L. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring. Soil Biology and Biochemistry, 2006, 38(9): 2843-2851. DOI:10.1016/j.soilbio.2006.04.042 |
| [46] |
Lipson D A, Schmidt S K. Seasonal changes in an alpine soil bacterial community in the Colorado Rocky Mountains. Applied and Environmental Microbiology, 2004, 70(5): 2867-2879. DOI:10.1128/AEM.70.5.2867-2879.2004 |
| [47] |
Edwards K A, Jefferies R L. Interannual and seasonal dynamics of soil microbial biomass and nutrients in wet and dry lowArctic sedge meadows. Soil Biology and Biochemistry, 2013, 57: 83-90. DOI:10.1016/j.soilbio.2012.07.018 |
| [48] |
Vidon P, Allan C, Burns D, Duval T P, Gurwick N, Inamdar S, Lowrance R, Okay J, Scott D, Sebestyen S. Hot spots and hot moments in Riparian Zones:potential for improved water quality management. Journal of the American Water Resources Association, 2010, 46(2): 278-298. |
| [49] |
Judd K E, Likens G E, Buso D C, Bailey A S. Minimal response in watershed nitrate export to severe soil frost raises questions about nutrient dynamics in the Hubbard Brook experimental forest. Biogeochemistry, 2011, 106(3): 443-459. DOI:10.1007/s10533-010-9524-4 |
| [50] |
Henry H A L. Climate change and soil freezing dynamics:historical trends and projected changes. Climatic Change, 2008, 87(3/4): 421-434. |
| [51] |
Haei M, Öquist M G, Kreyling J, Ilstedt U, Laudon H. Winter climate controls soil carbon dynamics during summer in boreal forests. Environmental Research Letters, 2013, 8(2): 024017. DOI:10.1088/1748-9326/8/2/024017 |