 2019, Vol. 39
2019, Vol. 39文章信息
- 郭燕, 杨邵, 沈雅飞, 肖文发, 程瑞梅
- GUO Yan, YANG Shao, SHEN Yafei, XIAO Wenfa, CHENG Ruimei
- 三峡水库消落带现存植物自然分布特征与群落物种多样性研究
- Study on the natural distribution characteristics and community species diversity of existing plants in the Three Gorges Reservoir
- 生态学报. 2019, 39(12): 4255-4265
- Acta Ecologica Sinica. 2019, 39(12): 4255-4265
- http://dx.doi.org/10.5846/stxb201801240190
-
文章历史
- 收稿日期: 2018-01-24
- 网络出版日期: 2019-04-01





2. 南京林业大学南方现代林业协同创新中心, 南京 210037
2. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
消落带(Water-level-fluctuation zone)随着大型水利工程兴建而兴起并越来越引起人类关注, 是由于水位周期性涨落而使库区周边被淹土地周期性出露于水面而形成的30 m落差的水陆系统交互地带, 其内部产生了明显的特殊环境因子、生态过程和植被群落梯度[1], 具水文、生物地球化学循环及生态和环境服务功能[2]。而且, 受反季节性水库水位周期性涨落的影响, 三峡库区消落带对外界扰动非常敏感, 容易发生水土流失、土壤受水陆交叉污染、生境退化、生态系统多样性减少等生态系统退化状况[3], 现已成为库区生态系统中物质转化、能量流动、信息传输与转换等最为活跃且最不稳定的生态脆弱带[4-5]。
消落带植被是长期生活在陆地的植物与周期性水陆交替变化生境下协同进化产生的, 具有特定的生态适应性, 其作为水库管理的重要对象, 是生态系统功能的主体[6], 在一定程度上可维护消落带生态系统的稳定和生态功能的发挥。消落区形成以后, 陆地生态环境尤其是植物群落发生巨大变化, 生态环境十分脆弱[7]。水淹是三峡库区消落带植被面临的最大干扰, 是影响植被组分及生活型、物种多样性、植物生长的一个重要环境因素[8]。当水淹干扰发生时, 植物群落组成的变化在很大程度上依赖于植物对水淹造成的缺氧土壤环境的耐受能力, 在水淹干扰梯度上, 海拔梯度、水淹持续时间及水淹强度是决定植被物种组成及物种分布的主要环境参数[9-10], 如Lomolino认为[11], 多样性与海拔梯度格局的关系(正相关、负相关或单峰关系)在很大程度上依赖于环境变量之间的协作与互作。植物群落的物种组成和结构变化是植物群落演变的具体表现特征[5]。目前, 前人对三峡库区消落带在水淹初期及经历短期水位涨落周年后的植被组成、群落结构及其动态进行了广泛研究, 但是随着水淹年限的延长, 消落带植物对于渐变的环境, 会开始通过其自身形态变异或生理过程的调节来增加与环境的适合度[12]。由于消落带生境的复杂性及外界干扰的影响, 库区群落结构仍具有很强的不稳定性和多变性, 在经历长期水位涨落周年后的库区物种空间分布特征和多样性会如何改变?植物会表现怎样特定的生活型来适应周期性的水陆交互环境?本研究即以2008年消落带典型区域固定样地野外调查资料为依托, 于2016年9月对经历长期水位涨落周年后的消落带植被的自然分布特征与群落物种多样性等进行复位调查研究, 以期从物种多样性角度探讨三峡库区消落带在经历长期水位涨落周年后(长期生境干湿交替)植物群落的结构变化特征, 同时对消落带植被现状进行分析, 进而为库区消落带植被恢复和重建提供科学的理论依据。
1 研究地点与研究方法 1.1 研究区域自然概况本研究区域位于湖北省秭归县茅坪镇, 紧邻三峡库区。位于30°38′14″—31°11′31″ N, 110°00′04″—110°18′41″ E, 地处亚热带大陆性季风气候区域, 气候温暖湿润, 四季分明;地质岩层以花岗岩为主, 研究区域土壤类型以黄壤土和黄棕壤为主, 土壤厚度约40 cm。≥10℃积温5723.6℃, 年均气温为19.0℃。年均降水为1100.0 mm, 年均相对湿度为78%。消落带回水区域内地形起伏较大, 易风化岩层出露较多, 降雨多且集中, 水土流失较严重。
1.2 样地设置与调查为了使调查样地能够较为全面地反映消落带秭归段植被及土壤环境特征, 于2008年8月将固定样地设置在秭归县茅坪镇典型消落带区域内, 固定样地从研究区有一定代表性的次生植被和弃耕地植被中选择。在每个样地沿消落带下部(高程145—155 m)、中部(155—165 m)和上部(165—175 m), 分别设置10 m×30 m样带, 每个样带内设置5个2 m×2 m的乔木幼苗幼树和灌木调查样方, 并在每个样方内分别设置5个1 m×1 m的草本调查样方, 间距10 m, 现场测量样方内每种植物的种名、盖度、数量、高度及植物生活型。其中, 丛生草本和灌木植株数量调查的方法是:以地面以上是否为单株存在为判别标准, 至于其地面以下植株与植株之间或不同植株的根系之间是否连接不予考虑。同时采用多功能坡度仪测定每个样方的坡度, 采用便携式手持GPS测定每个样方的海拔。野外调查时不能直接识别的, 现场直接依据彭镇华(2005)主编的《中国长江三峡植物大全》辨认识别[13]。
1.3 数据处理与分析数据处理及作图均采用Excel 2016、SPSS 17.0和Oringin 8.6软件进行。采用单因素方差分析(one-way ANONA)相关性分析和多重比较方法中的最小显著差异法(LSD)对植物物种多样性等指标进行显著性检验。
1.3.1 重要值选取重要值(Important value, IV)作为评价群落中各物种相对重要性的综合数量指标[14-15]。
分别计算乔木层、灌木层及草本层的重要值, 计算公式为:
重要值=(相对密度+相对频度+相对优势度)/3
灌草重要值=(相对密度+相对频度+相对盖度)/3
式中:相对密度= (某种植物的个体数/全部植物的个体数)×100%;
相对频度= (该种的频度/所有种的频度总和)×100%;
相对优势度= (样方中该种个体胸面积和/样方中全部个体胸面积总和)×100%;
相对盖度=(样方中该种个体的分盖度/样方中全部个体所有分盖度之和)×100%。
1.3.2 生物多样性指数生物多样性指数分析, 根据植被生态特征指数进行生物多样性的空间分布特征分析, 具体指标包括Shannon-Weiner指数、Pielou均匀度指数、Simpson优势度指数、丰富度指数S[16-17]。
Shannon-Weiner指数(H)表征群落的复杂程度, 即群落中生物种类增多, Shannon-Weiner指数值愈大, 群落所含的信息量愈大;
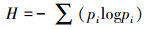
|
均匀度指数(E)表征群落中不同物种的多度、生物量、盖度或其他指标分布的均匀程度;
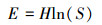
|
优势度指数又称Simpson指数, 表示植物群落内各植物种类处于何种优势或劣势状态的群落测定度, 是对多样性的反面即集中性的度量;
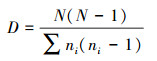
|
丰富度指数(S)可采用单位面积的物种数目表征, 即物种密度。
S=样方物种数
式中, pi表示第i个种的多度比例;S代表出现物种的个数;ni表示第i个物种的个体数;N代表出现物种的个体总数。
1.3.3 植物生活型祖成应用《中国植物志》对研究区植物生活型进行划分[18], 共区分出乔木、灌木、藤本、多年生草本、一年生草本等9种生活型。
2 结果与分析 2.1 植物种类组成消落带秭归段区域共发现和确认草本植被群落共有22科40属49种(表 1), 其中:禾本科(Gramineae) 6种6属;菊科(Compositae)9种9属;蓼科(Polygonaceae)3种1属;大戟科(Euphorbiaceae)6种5属;玄参科(Scrophulariaceae)2种2属;莎草科(Cyperaceae)3种2属;桑科(Moraceae)2种2属;壳斗科(Fagaceae)2种1属;葫芦科(Cucurbitaceae)2种2属;豆科(Leguminosae)5种4属;其他科:如荨麻科(Urticaceae)、旋花科(Convolvulaceae)、苋科(Amaranthaceae)、藤黄科(Guttiferae)、山矾科(Symplocaceae)、马钱科(Loganiaceae)、马齿苋科(Portulacaceae)、金缕梅科(Hamamelidaceae)、海金沙科(Lygodiaceae)、防己科(Menispermaceae)、紫草科(Boraginaceae)、黍亚科(Subfam)均为1属1种。禾本科植物最丰富, 其次是菊科植物, 蓼科植物和大戟科植物, 分别占此区域草本总科数的41.7%、29.1%、11.5%、7.6%, 是该区的主要优势科。其他科差别不大, 单种、属现象明显。
| 物种名称 Species |
科 Families |
属 Genera |
海拔 Altitude/m |
||
| 145—155 IV |
155—165 IV |
165—175 IV |
|||
| 狗牙根 Cynodon dactylon |
禾本科 Gramineae |
狗牙根属 Cynodon |
0.468 | 0.184 | 0.122 |
| 蜜甘草 Phyllanthus ussuriensis |
大戟科 Euphorbiaceae |
叶下珠属 Phyllanthus |
0.092 | 0.039 | 0.012 |
| 狗尾草 Setaria viridis |
禾本科 Gramineae |
狗尾草属 Setaria |
0.090 | 0.257 | 0.265 |
| 水田稗 Echinochloa oryzoides |
禾本科 Gramineae |
稗属 Echinochloa |
0.075 | 0.066 | 0.000 |
| 香附子 Rhizoma cyperi |
莎草科 Cyperaceae |
莎草属 Cyperus |
0.050 | 0.039 | 0.008 |
| 水蓼 Polygonum hydropiper |
蓼科 Polygonaceae |
蓼属 Polygonum |
0.039 | 0.014 | 0.001 |
| 酸模叶蓼 Polygonum lapathifolium |
蓼科 Polygonaceae |
蓼属 Polygonum |
0.035 | 0.007 | 0.002 |
| 毛马唐 Digitaria chrysoblephara |
禾本科 Gramineae |
马唐属 Digitaria |
0.027 | 0.159 | 0.124 |
| 喜旱莲子草 Alternanthera philoxeroides |
苋科 Amaranthaceae |
莲子草属 Alternanthera |
0.018 | 0.001 | 0.002 |
| 红蓼 Polygonum orientale |
蓼科 Polygonaceae |
蓼属 Polygonum |
0.017 | 0.018 | 0.019 |
| 狼把草 Bidentis Tripartitae |
菊科 Compositae |
鬼针草属 Bidens |
0.016 | 0.050 | 0.042 |
| 翅茎冷水花 Pilea subcoriacea |
荨麻科 Urticaceae |
冷水花属 Pilea |
0.011 | 0.014 | 0.008 |
| 铁苋菜 Acalypha australis |
大戟科 Euphorbiaceae |
铁苋菜属 Acalypha |
0.009 | 0.002 | — |
| 苍耳 Xanthium sibiricum |
菊科 Compositae |
苍耳属 Xanthium |
0.008 | 0.053 | 0.126 |
| 旱莲草 Eclipta prostrata |
菊科 Compositae |
鳢肠属 Eclipta |
0.008 | 0.016 | 0.028 |
| 葡茎通泉草 Mazus miquelii |
玄参科 Scrophulariaceae |
通泉草属 Mazus |
0.006 | 0.011 | — |
| 附地菜 Trigonotis peduncularis |
紫草科 Boraginaceae |
附地菜属 Trigonotis |
0.006 | — | — |
| 藿香蓟 Ageratum conyzoides |
菊科 Compositae |
藿香蓟属 Ageratum |
0.006 | 0.004 | 0.002 |
| 稗 Echinochloa crusgalli |
禾本科 Gramineae |
稗属 Echinochloa |
0.006 | 0.044 | 0.009 |
| 西瓜 Citrullus lanatus |
葫芦科 Cucurbitaceae |
西瓜属 Citrullus |
0.004 | — | — |
| 雍菜 Ipomoea aquatica |
旋花科 Convolvulaceae |
番薯属 Ipomoea |
0.003 | — | — |
| 冬瓜 Benincasa hispida |
葫芦科 Cucurbitaceae |
冬瓜属 Benincasa |
0.002 | — | — |
| 叶下珠 Phyllanthus urinaria |
大戟科 Euphorbiaceae |
叶下珠属 Phyllanthus |
0.002 | 0.001 | — |
| 一年蓬 Erigeron annuus |
菊科 Compositae |
飞蓬属 Erigeron |
0.001 | 0.002 | 0.062 |
| 斑地锦 Euphorbia maculata |
大戟科 Euphorbiaceae |
大戟属 Euphorbia |
0.001 | — | — |
| 紫萼蝴蝶草 Impatiens platychlaena |
玄参科 Scrophulariaceae |
蝴蝶草属 Torenia |
0.001 | 0.002 | 0.004 |
| 野艾蒿 Artemisia lavandulaefolia |
菊科 Compositae |
蒿属 Artemisia |
— | 0.009 | 0.080 |
| 鬼针草 Bidens bipinnata |
菊科 Compositae |
鬼针草属 Bidens |
— | 0.003 | 0.013 |
| 巴东醉鱼草 Buddleja albiflora |
马钱科 Loganiaceae |
醉鱼草属 Buddleja |
— | 0.002 | — |
| 乌桕 Sapium sebiferum |
大戟科 Euphorbiaceae |
乌桕属 Sapium |
— | 0.001 | 0.013 |
| 具芒碎米莎草 Cyperus microiria |
莎草科 Cyperaceae |
莎草属 Cyperus |
— | 0.001 | — |
| 马齿苋 Portulaca oleracea |
马齿苋科 Portulacaceae |
马齿苋属 Portulaca |
— | 0.000 | — |
| 枫香树 Liquidambar formosana |
金缕梅科 Hamamelidaceae |
枫香树属 Liquidambar |
— | — | 0.010 |
| 荩草 Arthraxon hispidus |
禾本科 Gramineae |
荩草属 Arthraxon |
— | — | 0.010 |
| 算盘子 Glochidion puberum |
大戟科 Euphorbiaceae |
算盘子属 Glochidion |
— | — | 0.006 |
| 土荆芥 Chenopodium ambrosioides |
藜科 Chenopodiaceae |
藜属 Chenopodium |
— | — | 0.004 |
| 合萌 Aeschynomene indica |
豆科 Leguminosae |
合萌属 Aeschynomene |
— | — | 0.004 |
| 截叶铁扫帚 Lespedeza cuneata |
豆科 Leguminosae |
胡枝子属 Lespedeza |
— | — | 0.003 |
| 白檀 Symplocos paniculata |
山矾科 Symplocaceae |
山矾属 Symplocos |
— | — | 0.003 |
| 短柄枹栎 Quercus serrata var . brevipetiolata |
壳斗科 Fagaceae |
栎属 Quercus |
— | — | 0.003 |
| 海金沙 Lygodium japonicum |
海金沙科 Lygodiaceae |
海金沙属 Lygodium |
— | — | 0.002 |
| 千里光 Senecio scandens |
菊科 Compositae |
千里光属 Senecio |
— | — | 0.002 |
| 茵陈蒿 Artemisia capillaris |
菊科 Compositae |
蒿属 Artemisia |
— | — | 0.002 |
| 地果 Ficus tikoua |
桑科 Moraceae |
榕属 Ficus |
— | — | 0.002 |
| 麻栎 Quercus acutissima |
壳斗科 Fagaceae |
栎属 Quercus |
0.001 | ||
| 两型豆 Amphicarpaea edgeworthii |
豆科 Leguminosae |
两型豆属 Amphicarpaea |
— | — | 0.001 |
| 胡枝子 Lespedeza chinensis |
豆科 Leguminosae |
胡枝子属 Lespedeza |
— | — | 0.001 |
| 葎草 Humulus scandens |
桑科 Moraceae |
葎草属 Humulus |
— | — | 0.001 |
| 木防己 Cocculus orbiculatus |
防己科 Menispermaceae |
木防己属 Cocculus |
— | — | 0.000 |
| IV:重要值Important value; “—”表示物种消失 | |||||
低海拔消落带草本植物物种数最少, 对物种要求最为严格, 如附地菜(Trigonotis peduncularis)、雍菜(Ipomoea aquatica)、斑地锦(Euphorbia maculata)等5个物种只出现在此海拔;随海拔增加, 许多“新”植物陆续出现:相比海拔145—155 m区段, 海拔155—165 m区段增加铁苋菜(Acalypha australis)、葡茎通泉草(Mazus miquelii)、叶下珠(Phyllanthus urinaria)等6个物种;海拔165—175 m区段则增加巴东醉鱼草(Buddleja albiflora)、具芒碎米莎草(Cyperus microiria)、马齿苋(Portulaca oleracea)等13个物种。可见经历水库水位的涨落次数、淹水深度、水淹持续时间等对消落带植物具有不同的影响效应。另外, 不同海拔处的优势物种不同:海拔145—155 m处主要优势物种是狗牙根(Cynodon dactylon)、蜜甘草(Phyllanthus ussuriensis)和狗尾草(Setaria viridis);海拔155—165 m处主要优势物种是狗尾草、狗牙根和毛马唐(Digitaria chrysoblephara);而海拔165—175 m处主要优势物种是狗尾草、苍耳(Xanthium sibiricum)和毛马唐。
2.2 群落物种多样性根据各海拔记录物种数据, 分析物种生活型组成沿海拔的变化, 得出消落带秭归段植物群落的物种多样性指数(即丰富度指数S、Shannon-Weiner指数H、均匀度指数E、Simpson指数)沿水位海拔的变化如图 1, 可以看出:受三峡库区水位波动的影响, 消落带植物群落中的物种多样性指数均在低海拔区段(145—155 m)达到最低值, 其中:物种丰富度指数随海拔的变化范围为29.6—45.8;Shannon-Weiner指数随海拔的变化范围为2.24—2.84;以个体数量和物种重要值为基础的物种均匀度指数随海拔的变化范围为0.68—0.76;优势度指数随海拔的变化范围为0.67—0.81, 最高值均出现在海拔155—165 m区段, 这可能是因为低海拔区段在水库运行期间的淹水时间最长, 受水淹胁迫程度最大, 仅有狗牙根等少数耐水淹胁迫种可以形成稳定群落, 其他植物都很难定居。

|
| 图 1 沿高程梯度的消落带物种多样性格局 Fig. 1 Species diversity in the falling zone along the elevation gradient |
一般认为水淹强度越大, 出陆时间越短, 植物多样性越低;水淹强度越小, 出陆时间越长, 植物多样性越高[19]。通过对库区消落带不同海拔区段物种多样性指数沿海拔进行模拟分析可得:海拔高度与物种丰富度、多样性指数和均匀度指数之间的趋势模拟均呈幂指数关系, 其拟合系数r值分别为0.834、0.824、0.817和0.808。由此可推出:库区物种多样性随海拔梯度的变化呈现“单峰”分布格局, 即随着海拔的升高, 物种丰富度、多样性、均匀度和优势度均表现先增加后降低的趋势, 中等海拔(155—165 m)的植物物种多样性最高, 这与一些研究学者的研究结论相一致[20]。这可能是由于中海拔区段资源组合呈现最优化状态, 就资源可利用性而言, 物种多样性的海拔梯度分布格局表达了多样性-资源生产力之间的单调关系内涵[21]。
2.3 植物生活型特征消落带秭归段区域物种共包含9种生活型(表 2), 其中, 海拔145—155 m区段生活型4种;海拔155—165 m区段生活型4种;海拔165—175 m区段7种。海拔145—155、155—165、165—175 m区段内不同生活型比例差异较大, 均以一年生的草本为优势物种(图 2), 其中:海拔145—155 m区段一年生的草本19种, 占统计植物种的73.1%;多年生草本植物5种, 占统计植物种的19.2%;一、二年生草本植物1种, 占统计植物种的3.8%;蔓生草本2种, 占统计植物种的3.8%。海拔155—165 m区段中, 一年生的草本植物18种, 占统计植物种的66.7 %;多年生草本植物7种, 占统计植物种的25.9%;一、二年生草本植物1种, 占统计植物种的3.7%;落叶乔木1种, 占统计植物种的3.7%。海拔165—175 m区段中, 一年生的草本植物20种, 占统计植物种的52.6%;多年生草本植物8种, 占统计植物种的21.1%;一、二年生草本植物1种, 占统计植物种的0.9%, 其他生活型:如乔灌木、一年或多年生植物、蔓生草本、蕨类各1种, 共占统计植物种的7.9%。与低海拔145—155 m相比, 海拔165—175 m区段的植物生活型更加多样化, 如藤本、灌木以及落叶乔木等生活型均出现, 且物种数随海拔增加表现为逐渐增多的规律, 显著性分析可知:海拔增加并没有明显降低主要生活型所占的比例, 但会增加生活型的种类。
| 海拔 Altitude/m |
种数 Number |
生活型 Life type |
分比例/% Partial proportion |
总比例/% Total proportion |
| 145—155 | 19 | 一年生草本 | 73.1 | 24.4 |
| 5 | 多年生草本 | 19.2 | 6.4 | |
| 1 | 一、二年生草本 | 3.8 | 1.3 | |
| 1 | 蔓生草本 | 3.8 | 1.3 | |
| 155—165 | 18 | 一年生草本 | 66.7 | 22.2 |
| 7 | 多年生草本 | 25.9 | 8.6 | |
| 1 | 一、二年生草本 | 3.7 | 1.2 | |
| 1 | 落叶乔木 | 3.7 | 1.2 | |
| 165—175 | 20 | 一年生草本 | 52.6 | 17.5 |
| 8 | 多年生草本 | 21.1 | 7.0 | |
| 1 | 一年或二年生草本 | 2.6 | 0.9 | |
| 9 | 其他 | 23.7 | 7.9 | |
| 分比例指某一生活型占该海拔物种数的比例;总比例指某一生活型占全部海拔物种数的比例;其他生活型:一年或多年生草本、木质藤本和蕨类 | ||||

|
| 图 2 三峡库区消落带植物生活型组成沿海拔的变化规律 Fig. 2 Variation of plant life type along the elevation gradient in the riparian zone of the Three Gorges Reservoir |
三峡大坝建成后, 消落带原有物种的生境条件被彻底打破, 周期性的淹水-落干-淹水的干湿交替模式使得原有植被群落的结构和功能处于剧烈波动的退化阶段[22], 受周期性、反季节、高强度水淹影响, 植物多样性明显下降[14, 22-23]。一般认为, 植物对渐变的环境, 可以通过自身的形态变异或生理过程调节来增加其与环境的适合度[12], 但这种“变异”和“调节”需经历一个渐变过程。三峡水库建设初期, 王勇等曾对三峡库区自然消落区植物区系进行研究, 表明消落区分布有维管植物83科、240属、405种[24], 但2009年对蓄水后水库消落区植物区系的调查表明, 维管植物只有61科169属231种, 其中科、属、种分别减少了26.51%、29.58%和42.96%, 优势生活型由多年生草本转变为一年生草本, 植物群落结构也比较脆弱[25-26]。同样, 白宝伟等[1], 杨朝东等[27], 王强等[28]对三峡水库消落带植物群落格局及多样性进行研究表明:较三峡大坝建设前, 消落带植物物种数减少, 单属单种现象明显, 群落组成简单化。对消落带秭归段区域, 王晓荣等[29]曾对水淹初期(2009年)植物群落进行研究发现, 初次淹水改变了原有消落带物种组成, 未淹区段共发现植物物种84科163属223种, 以多年生草本为主要生活型;本研究则以原有固定样地为基础, 通过分析经历8次水位涨落周年后的植物自然分布特征得出:现存植被群落共发现22科40属49种, 较2009年, 其科、属、种数的减少百分比分别高达73.8%、75.5%和78.0%, 且优势生活型转为一年生草本植物, 单属、种现象明显, 群落结构简单化, 这也与上述结论相吻合, 表明在水库的30 m水位波动和“半年水淹-半年干旱”的模式产生的极其剧烈的干扰下, 只有少数物种能够适应消落带这一新生生态系统[30]。其原因可能是由于陆生植物缺乏适应这种骤然、长期深水淹没生境的机体结构和功能[12];水淹干扰后的土壤水分状况及土壤供养状况以及水库水位的高低、涨落、变动频率、持续时间、发生时间等特征也可能在一定程度影响植物群落的种类组成及空间分布, 进而成为植物定居、生长和繁殖的限制性因子[10]。另外, 本实验通过对经历短期和长期水位涨落周年后的物种特性进行整理和比较可得:随水淹年限及海拔梯度增加, 消落带植物自身在其结构和生理等方面产生了一系列适应性, 不同海拔梯度均出现不同于其他海拔梯度的“新”植物, 当然这并非是植物与环境长期协同进化意义上的“新种”, 其可能来自邻近区域, 而更多的可能是在经历水库水位涨落前以种子库的形式在消落带存在的植物种[5]。
消落带是水位反复周期变化的干湿交替区, 周期性的反季节淹水环境形成了大量次生裸地, 消落带植物在该环境下生长繁衍, 其植物群落的结构特征沿环境梯度存在一定的变化规律, 而海拔被认为是影响植物群落多样性分布格局的重要因素之一[31-35], 不同海拔梯度植物由于受水淹胁迫程度不同, 其植被分布自低海拔向高海拔也发生了很大的变化。众多研究表明, 物种多样性随着海拔梯度的变化有多种形式, 其中普遍认为二者呈负相关关系, 即随着海拔高度的增加物种多样性降低[21, 36];另一种形式是“中间高度膨胀”, 即随着海拔高度的增加, 物种多样性呈先增加、后减少的趋势[21, 37]。在本研究中所得出的结论符合“中间高度膨胀”理论:受三峡库区水位波动的影响, 消落带植被的物种多样性指数均在低海拔区段(145—155 m)达到最低值;经历长期水位涨落周年后, 随海拔的增加, 库区消落带草本植被的S、H、E、D指数均表现为先升后降的趋势, 即物种多样性在中海拔区段(155—165 m)达到最高。这可能是因为水库消落带高海拔区段所受水淹胁迫较小, 物种定居较易, 导致某些物种数量较多, 其中狗尾草、毛马唐、苍耳群落的优势度较为明显, 其他植物群落多属于从属地位。水库消落区中海拔区段各植物群落相比上部差距缩小, 部分竞争种选择退避(如铁苋菜、附地菜、匍茎通泉草和叶下珠等), 而另一部分耐水淹胁迫种竞争力上升(如狗牙根, 香附子、水田稗等), 这就造成了水库消落区中部植物群落结构相比上部和下部都更为复杂, 共优群落相比单优群落在竞争中更具有优势[19], 从而造成群落稳定性更强、物种多样性更高, 其中物种狗尾草和狗牙根成为主要优势种。水库消落带低海拔区段则由于所受水淹胁迫最强, 出露时间最短, 仅有少数几种耐水淹胁迫种可以形成稳定群落(如狗牙根等), 植物群落结构趋于单一化, 这说明随水淹年限增加, 消落带物种的多样性受各海拔梯度的环境资源结构的影响显著。
Grime提出了植物适应对策的C-R-S对策模型, R-对策种适应于临时性资源丰富的环境;C-对策种生存于资源一直处于丰富的生境中, 竞争力强, 称为竞争种;S-对策种生存于资源较为贫瘠的生境, 抗逆性强, 称之为耐胁迫种[19], 一般认为先锋种多为R-对策种, 演替中期多为C-对策种, 而顶级群落中多为S-对策种[38]。经过多次水位涨落后成为消落带优势种的生活史对策大多具有R对策种群特征。毛马唐、狗尾草、苍耳等优势种多属于田间和坡地常见的一年生草本。毛马唐自然生长多见于路旁、田野, 喜湿、喜光, 种子耐受水淹的能力较强;狗尾草多生于林边、山坡、路边和荒芜的园地及荒野, 对干燥生境有较强的适应能力;苍耳喜温暖稍湿润气候, 耐干旱瘠薄, 且具钩状的硬刺, 常贴附于家畜和人体上, 种子易于散布。这些物种多以短的生命周期, 高的相对生长率和高的种子产量为特征, 能够适应生境的强烈干扰并将有限生境资源大部分分配给种子, 属于“杂草策略”。狗牙根、香附子等优势种则属于多年生草本。狗牙根多生长于村庄附近、道旁河岸、荒地山坡, 根系发达, 具有能够克隆繁殖的匍匐状地下根茎, 其侵占性、再生性及抗恶劣环境能力极强;香附子多生于荒地、路边、沟边或田间向阳处, 以块茎和种子繁殖, 耐胁迫能力较强, 这些物种多采用以防御和减轻恶劣环境胁迫为特征的“胁迫忍耐策略”。对经历长期水位涨落周年后物种特性总结可知, 消落带物种能够适应其周期性、反季节、高强度的水淹条件与自身生长繁殖策略密切有关, 特征为:多以大量细小种子繁殖或以耐无氧呼吸的茎段进行营养繁殖, 能利用退水后的有限出露时间(夏季和秋季)完来成其生活史[29-30, 39], 然后次年依靠临近种源或者土壤种子库开始新的生命周期[40-43], 这在一定程度上可以很好的解释消落带物种的耐胁迫机制。
各个生态因子是综合对植物产生影响的, 植物对这些因子产生选择, 在生理、结构、形态上发生相应变化, 其对外界环境的外部形态表现形式即生活型;生活型的差别可以一定程度反映群落生境条件的改变, 尤其是小生境、小气候的变化。海拔梯度的变化综合了温度、湿度和光照等多种环境因子, 因此, 在一定程度上能够显著地影响植物种群的空间分布格局[37]。王晓荣等[29]对秭归段消落带水淹初期后植物群落进行研究发现, 初次淹水后消落带优势生活型为多年生草本植物, 其物种组成中一年生草本占20.45%, 多年生草本占36.36%, 灌木或藤本占28.18%, 乔木占15%。而本研究调查时期内不同海拔区段消落带植物优势生活型为一年生草本植物, 占统计植物种的34.1%, 多年生草本占22.0%, 灌木或藤本占2.2%, 乔木占6.6%, 可见周期性的淹水环境使得群落结构简单化, 不利于消落带生态系统的稳定性。低海拔区段仅发现四种生活型, 但随海拔梯度的增加, 库区植物的生活型更加多样化, 其他生活型如藤本、灌木以及落叶乔木逐渐出现且表现为逐渐增多, 至2016年, 秭归段消落带各海拔区域共出现9种生活型, 其中, 海拔145—155 m区段生活型4种;海拔155—165 m区段生活型4种;海拔165—175 m区段8种。植物生活型是不同植物对相同生境趋同适应的外在表现[44], 植物生活型适应于水陆生境变化的策略决定了植物群落的物种组成和替代变化趋势, 同时也是消落带植物群落在水陆生境变化下演替的基础[5]。因此本实验一定程度证明:在三峡水库反复周期性水位涨落影响下, 草本植物(包括一年生和多年生植物)生活型取代乔木、灌木和藤本及其他生活型是三峡库区消落带植物生活型组成变化的必然趋势。
综上所述:在经历8次水位涨落周年后, 三峡库区秭归段消落带植物多样性和植物群落结构发生较大改变, 其植被多样性降低, 且海拔差异性显著, 最大的物种多样性出现在中海拔区段;区域群落结构简单化, 以一年生草本为优势生活型, 且单种、属现象明显, 群落多样性降低。就植物群落结构和稳定性而言, 三峡水库消落区植物群落结构及物种受环境资源结构决定, 在不同海拔消落带呈现出特定的R生存对策。目前, 由于各海拔消落带区段历经周期性水淹长达8年之久, 其植物群落稳定性显著低于水淹初期;且库区物种在资源匮乏且不稳定的生境中的种间竞争依然强烈, 消落带植被仍处于群落演替的初级阶段。
致谢: 沈雅飞、杨邵帮助实验,特此致谢。| [1] |
白宝伟, 王海洋, 李先源, 冯义龙, 智丽. 三峡库区淹没区与自然消落区现存植被的比较. 西南农业大学学报:自然科学版, 2005, 27(5): 684-687, 691-691. |
| [2] |
张建春. 河岸带功能及其管理. 水土保持学报, 2001, 15(6): 143-146. DOI:10.3321/j.issn:1009-2242.2001.06.038 |
| [3] |
刘冬燕.三峡库区消落带生态恢复与重建的研究——以万州消落带示范区为例.上海: 同济大学, 2005. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y820906
|
| [4] |
胡波, 张平仓, 任红玉, 罗慧敏, 岑奕. 三峡库区消落带植被生态学特征分析. 长江科学院院报, 2010, 27(11): 81-85. DOI:10.3969/j.issn.1001-5485.2010.11.017 |
| [5] |
朱妮妮, 郭泉水, 秦爱丽, 裴顺祥, 马凡强, 朱莉, 简尊吉. 三峡水库奉节以东秭归和巫山段消落带植物群落动态特征. 生态学报, 2015, 35(23): 7852-7867. |
| [6] |
郭泉水, 康义, 洪明, 金江群, 朱妮妮, 聂必红, 王佐庆. 三峡库区消落带陆生植被对首次水陆生境变化的响应. 林业科学, 2013, 49(5): 1-9. DOI:10.3969/j.issn.1672-8246.2013.05.001 |
| [7] |
饶丽, 朱振亚. 三峡库区消落带环境整治与生态修复调查研究. 环境影响评价, 2016, 38(3): 77-80. |
| [8] |
卢志军, 李连发, 黄汉东, 陶敏, 张全发, 江明喜. 三峡水库蓄水对消涨带植被的初步影响. 武汉植物学研究, 2010, 28(3): 303-314. |
| [9] |
Blom C W P M, Voesenek L A C J. Flooding:the survival strategies of plants. Trends in Ecology & Evolution, 1996, 11(7): 290-295. |
| [10] |
王正文, 邢福, 祝廷成, 李宪长. 松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应. 植物生态学报, 2002, 26(6): 708-716. DOI:10.3321/j.issn:1005-264X.2002.06.012 |
| [11] |
Lomolino M V. Elevation gradients of species-density:historical and prospective views. Global Ecology and Biogeography, 2001, 10(1): 3-13. DOI:10.1046/j.1466-822x.2001.00229.x |
| [12] |
常杰, 葛滢. 生态学. 杭州: 浙江大学出版社, 2001.
|
| [13] |
彭镇华. 中国长江三峡植物大全 (上卷). 科学出版社, 2005.
|
| [14] |
贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征. 植物生态学报, 1998, 22(4): 303-311. DOI:10.3321/j.issn:1005-264X.1998.04.003 |
| [15] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [16] |
陈忠礼, 袁兴中, 刘红, 李波. 水位变动下三峡库区消落带植物群落特征. 长江流域资源与环境, 2012, 21(6): 672-677. |
| [17] |
张志永, 程郁春, 程丽, 万成炎, 李金波. 三峡库区万州段消落带植被及土壤理化特征分析. 水生态学杂志, 2016, 37(2): 24-33. |
| [18] |
中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1977: 386.
|
| [19] |
刘维暐, 王杰, 王勇, 杨帆. 三峡水库消落区不同海拔高度的植物群落多样性差异. 生态学报, 2012, 32(17): 5454-5466. |
| [20] |
Wang G H, Zhou G S, Yang L M, Li Z Q. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecology, 2003, 165(2): 169-181. DOI:10.1023/A:1022236115186 |
| [21] |
王国宏. 祁连山北坡中段植物群落多样性的垂直分布格局. 生物多样性, 2002, 10(1): 7-14. DOI:10.3321/j.issn:1005-0094.2002.01.002 |
| [22] |
王勇, 吴金清, 黄宏文, 刘松柏. 三峡库区消涨带植物群落的数量分析. 植物科学学报, 2004, 22(4): 307-314. DOI:10.3969/j.issn.2095-0837.2004.04.006 |
| [23] |
Wu J G, Huang J H, Han X G, Xie Z Q, Gao X M. Three-Gorges Dam-experiment in habitat fragmentation. Science, 2003, 300(5623): 1239-1240. DOI:10.1126/science.1083312 |
| [24] |
王勇, 厉恩华, 吴金清. 三峡库区消涨带维管植物区系的初步研究. 植物科学学报, 2002, 20(4): 265-274. DOI:10.3969/j.issn.2095-0837.2002.04.005 |
| [25] |
Yang F, Liu W W, Wang J, Liao L, Wang Y. Riparian vegetation's responses to the new hydrological regimes from the Three Gorges Project:clues to revegetation in reservoir water-level-fluctuation zone. Acta Ecologica Sinica, 2012, 32(2): 89-98. DOI:10.1016/j.chnaes.2012.02.004 |
| [26] |
刘维暐, 杨帆, 王杰, 王勇. 三峡水库干流和库湾消落区植被物种动态分布研究. 植物科学学报, 2011, 29(3): 296-306. |
| [27] |
杨朝东, 张霞, 向家云. 三峡库区消落带植物群落及分布特点的调查. 安徽农业科学, 2008, 36(31): 13795-13796, 13866-13866. DOI:10.3969/j.issn.0517-6611.2008.31.132 |
| [28] |
王强, 袁兴中, 刘红, 张跃伟, 陈忠礼, 李波. 三峡水库初期蓄水对消落带植被及物种多样性的影响. 自然资源学报, 2011, 26(10): 1680-1693. DOI:10.11849/zrzyxb.2011.10.005 |
| [29] |
王晓荣, 程瑞梅, 肖文发, 潘磊, 曾立雄. 三峡库区消落带水淹初期主要优势草本植物生态位变化特征. 长江流域资源与环境, 2016, 25(3): 404-411. DOI:10.11870/cjlyzyyhj201603006 |
| [30] |
童笑笑, 陈春娣, 吴胜军, 贾振毅, 易雪梅, 马茂华. 三峡库区澎溪河消落带植物群落分布格局及生境影响. 生态学报, 2018, 38(2): 571-580. |
| [31] |
Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature, 2006, 441(7093): 629-632. DOI:10.1038/nature04742 |
| [32] |
Ives A R, Carpenter S R. Stability and diversity of ecosystems. Science, 2007, 317(5834): 58-62. DOI:10.1126/science.1133258 |
| [33] |
段敏杰, 高清竹, 郭亚奇, 万运帆, 李玉娥, 干珠扎布, 旦久罗布, 韦兰亭, 西饶卓玛. 藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局. 草业科学, 2011, 28(10): 1845-1850. |
| [34] |
齐代华, 贺丽, 周旭, 刘成, 闵鹏. 三峡水库消落带植物物种组成及群落物种多样性研究. 草地学报, 2014, 22(5): 966-970. |
| [35] |
郝占庆, 于德永, 杨晓明, 丁之慧. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 2002, 13(7): 785-789. DOI:10.3321/j.issn:1001-9332.2002.07.005 |
| [36] |
唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释. 生物多样性, 2004, 12(1): 115-122. DOI:10.3321/j.issn:1005-0094.2004.01.014 |
| [37] |
Oommen M A, Shanker K. Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants. Ecology, 2005, 86(11): 3039-3047. DOI:10.1890/04-1837 |
| [38] |
蒋志刚, 马克平, 韩兴国. 保护生物学. 杭州: 浙江科学技术出版社, 1997.
|
| [39] |
李俊清. 森林生态学(第二版). 北京: 高等教育出版社, 2010.
|
| [40] |
Favre-Bac L, Ernoult A, Mony C, Rantier Y, Nabucet J, Burel F. Connectivity and propagule sources composition drive ditch plant metacommunity structure. Acta Oecologica, 2014, 61: 57-64. DOI:10.1016/j.actao.2014.10.006 |
| [41] |
Merritt D M, Wohl E E. Processes governing hydrochory along rivers:hydraulics, hydrology, and dispersal phenology. Ecological Applications, 2002, 12(4): 1071-1087. DOI:10.1890/1051-0761(2002)012[1071:PGHARH]2.0.CO;2 |
| [42] |
Säumel I, Kowarik I. Urban rivers as dispersal corridors for primarily wind-dispersed invasive tree species. Landscape and Urban Planning, 2010, 94(3/4): 244-249. |
| [43] |
Schmiedel D, Tackenberg O. Hydrochory and water induced germination enhance invasion of Fraxinus pennsylvanica. Forest Ecology and Management, 2013, 304: 437-443. DOI:10.1016/j.foreco.2013.04.027 |
| [44] |
蒋有绪, 郭泉水, 马娟. 中国森林群落分类及其群落学特征. 北京: 科学出版社, 1998.
|