 2019, Vol. 39
2019, Vol. 39文章信息
- 林艳, 何跃军, 何敏红, 吴春玉, 方正圆, 韩勖, 徐鑫洋, 王世雄
- LIN Yan, HE Yuejun, HE Minhong, WU Chunyu, FANG Zhengyuan, HAN Xu, XU Xinyang, WANG Shixiong
- 喀斯特植被演替过程土壤丛枝菌根真菌(AMF)多样性
- Species diversity of soil arbuscular mycorrhizal fungi in karst vegetation succession process
- 生态学报. 2019, 39(11): 4127-4137
- Acta Ecologica Sinica. 2019, 39(11): 4127-4137
- http://dx.doi.org/10.5846/stxb201807061475
-
文章历史
- 收稿日期: 2018-07-06
- 网络出版日期: 2019-03-21

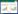




丛枝菌根真菌(Arbuscular mycorrhizal fungi, AMF)是一类能与绝大部分植物根系形成共生体, 广泛分布于自然界中, 并对植物的生长及发育有着极其重要作用的菌根真菌[1-2]。AMF多样性影响着不同生态系统的植物群落结构、多样性和生产力, 并在生态系统的植被演替及恢复过程中扮演着重要角色[3-4]。
中国西南喀斯特溶岩地区是世界三大溶岩区之一, 且是连片裸露碳酸盐岩面积最大的片区, 由于其特殊而复杂的土壤侵蚀过程导致该地区水土流失并形成石漠化, 严重阻碍了区域生态平衡[5-6], 石漠化治理成为维护区域社会经济与生态环境的可持续发展的一项重要内容[7]。喀斯特植被恢复受微生物群落结构的影响, 有研究者认为AMF可能成为喀斯特石漠化治理中采用菌根生物技术应用的重要选择, 其前提是要筛选出适合于该区域的AMF菌种[8]。虽然已有研究学者对喀斯特地区AMF的研究开展了一些前期工作, 如AMF对喀斯特生境植物光合生理的作用[9]、氮磷营养吸收[10]及对寄主植物抗旱性的影响[11-12]等方面, 也有少数作者进行喀斯特生境下的AMF形态鉴定的多样性分析[13]或遗传多样性分析[14-16]。充分了解喀斯特生境中的AMF多样性及分布状况, 有助于将AMF运用于石漠化治理[17], 有学者曾研究草原生态系统及退化生态系统中的AMF多样性, 表明AMF多样性会随着植被演替的进程而发生动态的变化[18-20], 而喀斯特植被自然演替过程中AMF多样性变化的研究仍然不够深入, 喀斯特生境中不同植被自然演替阶段的土壤AMF多样性概况如何?因此, 本研究旨在通过高通量测序的方法, 探究喀斯特不同植被自然演替过程中土壤AMF种质资源多样性, 为AMF在喀斯特石漠化生态恢复过程中采用菌根生物技术提供理论基础。
1 材料与方法 1.1 试验设计与土样采集本研究选取了3个典型喀斯特自然演替区域作为研究对象, 其中, 贵州贵阳市花溪区(简称花溪, 106°27′—106°52′E, 26°11′—26°34′N), 为高原季风湿润气候;关岭县花江板贵乡(简称花江, 105°36′30″—105°46′30″E, 25°39′13″—25°41′00″N), 属亚热带季风气候;毕节市织金县珠藏镇(简称织金, 105°44′42″—106°11′38″E, 26°38′31″—26°52′35″N), 属亚热带季风气候。在每个研究区根据植被类型的分布概况, 以空间替代时间方法按植被演替方向选择草本、灌木及乔木3种不同植被类型覆盖的地段进行样地设置, 共9个样地(每一样地3个重复)。按乔木10 m×10 m、灌木5 m×10 m、草本2 m×2 m的样方面积进行群落学调查, 并按样方对角线五点取样法进行土壤样品采集。采样时挖取土壤剖面, 每取样点按5、10、15cm分层取样, 将各层土壤等量充分混匀装入保鲜袋, 分为两份, 一份用于理化性质测定, 一份带回实验室置于-80℃超低温冰箱中保存, 送至北京诺禾致源测序公司检测。研究样地其他信息概况如表 1。
| 采样点 Sample site |
海拔 Elevation/m |
岩石裸露度 Rock exposed degree/% |
植被覆盖率 Vegetation coverage/% |
年均温 Average annual temperature/℃ |
年均降雨量 Average annual rainfall/mL |
主要植物 Main plants |
|
| HX | T | 1165 | <5 | 100 | 15.3 | 1129.5 | 香樟Cinnamomum camphora 、青冈栎Cyclobalanopsis glauca |
| B | 1263 | 20—30 | 70—80 | 15.6 | 1130 | 鼠刺Itea chinensis、 火棘Pyracantha fortuneana |
|
| H | 1106 | <5 | 80—90 | 15.9 | 1200 | 五节芒Miscanthus floridulus、 白茅Imperata cylindrica |
|
| HJ | T | 951 | 20 | 90 | 16.2 | 1200.2 | 臭椿Ailanthus altissima、 山桐子Idesia polycarpa |
| B | 844 | 20—30 | 70—80 | 16.8 | 1200 | 乌桕Sapium sebiferum、 地瓜藤Caulis Fici Tikouae |
|
| H | 957 | 70—80 | 40—50 | 16.5 | 1210 | 荩草Arthraxon hispidus、 艾蒿Artemisia argyi |
|
| ZJ | T | 1564 | <57 | 100 | 14.1 | 1430 | 光皮桦Betula luminifera、 杉木Cunninghamia lanceolata |
| B | 1437 | <5 | 100 | 14.3 | 1400 | 鼠刺Itea chinensis、 铁芒萁Dicranopteris dichotoma |
|
| H | 1437 | 20—30 | 20 | 14.8 | 1450 | 白酒草Conyza japonica、 荩草Arthraxon hispidus |
|
| HX:花溪, HuaXi;HJ:花江, HuaJiang;ZJ:织金, ZhiJing;T:乔木演替阶段, Tree;B:灌木演替阶段, Bush;H:草本演替阶段, Herb | |||||||
采用SDS方法对样本基因组DNA进行提取, 利用琼脂糖凝胶电泳检测DNA的纯度和浓度, 取适量的样品于离心管中, 使用无菌水稀释样品至1 ng/μL。以稀释后的基因组DNA为模板, 根据测序区域的选择, 使用带Barcode的特异引物, New England Biolabs公司的Phusion® High-Fidelity PCR Master Mix with GC Buffer, 和高效高保真酶进行PCR。引物参照Beenhouwer[21]、Dai[22]、Geel[23]、Liang[24]、Xiang等[25]人提出的土壤AMF特异性引物, 序列为:AMV4.5NF(F)—AAGCTCGTAGTTGAATTTCG、AMDGR(R)—CCCAACTATCCCTATT AATCAT, 由诺禾致源公司合成。PCR产物使用2%浓度的琼脂糖凝胶进行电泳检测;根据PCR产物浓度进行等量混样, 充分混匀后使用2%的琼脂糖凝胶电泳检测PCR产物, 对目的条带使用qiagen公司提供的胶回收试剂盒回收产物。
1.2.2 文库构建和上机测序使用TruSeq®DNA PCR—Free Sample Preparation Kit建库试剂盒进行文库构建, 构建好的文库经过Qubit和Q—PCR定量, 文库合格后, 使用HiSeq2500 PE250进行上机测序。
1.2.3 测序数据处理根据Barcode序列和PCR扩增引物序列从下机数据中拆分出各样品数据, 截去Barcode和引物序列后使用FLASH[26]对每个样品的序列进行拼接, 将得到的拼接序列经过严格过滤处理[27]得到高质量Tags数据, 参照Qiime(Version 1.7.0)[28]的Tags质量控制流程。将经过以上处理后得到的Tags序列通过UCHIME Algorithm [29]与Unite database数据库进行比对检测嵌合体序列, 去除其中的嵌合体序列[30], 获得最终有效数据。
1.2.4 OTU聚类和物种注释利用Uparse软件(Uparse v7.0.1001)[31]对所有样品的全部有效数据进行聚类, 默认以97%的一致性将序列聚类成为OTU(Operational Taxonomic Units), 依据其算法原则, 筛选OTU中出现频数最高的序列作为OTU的代表序列。对OTU代表序列进行物种注释, 用Qiime软件(Version 1.7.0)中的blast方法[32]与Unit数据库[33]进行物种注释分析, 并分别在各个分类水平:phylum(门)、class(纲)、order(目)、family(科)、genus(属)、species(种)统计各样本的AMF群落组成。使用MUSCLE[34]软件进行快速多序列比对, 得到所有OTU代表序列的系统发生关系。
1.3 土壤理化性质测定采用常规方法测定土壤理化性质[35], 其中, 土壤pH值酸度计法;速效钾采用火焰分光光度法;有效磷采用钼锑抗比色法;全磷采用钼锑抗比色法;全氮和水解氮均用半微量开氏法;有机质用油浴加热重铬酸钾氧化法测定(表 2)。
| 采样点 Sample site |
有机质 Organic matter/ (g/kg) |
pH | 速效磷 Available phosphorus/ (mg/kg) |
水解氮 Hydrolysis nitrogen/ (mg/kg) |
速效钾 Available kalium/ (mg/kg) |
全氮 Total nitrogen/ (g/kg) |
全磷 Total phosphorus/ (g/kg) |
|
| HX | T | 10.08±1.06a | 7.57±0.06a | 0.47±0.03a | 0.45±0.04b | 17.97±3.26b | 0.12±0.01ab | 0.18±0.02b |
| B | 11.70±3.89a | 7.67±0.03a | 0.51±0.06a | 0.58±0.06a | 27.01±2.39a | 0.16±0.03a | 0.21±0.02ab | |
| H | 13.36±0.15a | 7.58±0.06a | 0.52±0.02a | 0.46±0.12ab | 25.16±4.39a | 0.11±0.01b | 0.23±0.02a | |
| HJ | T | 9.84±0.77b | 7.68±0.04a | 0.57±0.04a | 0.58±0.04a | 28.67±3.09a | 0.15±0.01a | 0.50±0.06a |
| B | 11.52±0.88ab | 7.76±0.02a | 0.48±0.01b | 0.24±0.02c | 27.72±2.81a | 0.22±0.13a | 0.18±0.01b | |
| H | 11.54±0.92a | 7.49±0.05b | 0.35±0.01c | 0.32±0.05b | 8.86±0.37b | 0.07±0.01b | 0.18±0.01b | |
| ZJ | T | 10.04±1.04b | 5.44±0.10b | 0.26±0.07b | 0.26±0.03b | 14.65±3.03a | 0.09±0.02b | 0.39±0.01a |
| B | 10.06±0.42b | 5.96±0.01b | 0.28±0.05b | 0.34±0.02a | 6.89±0.41b | 0.09±0.01b | 0.16±0.01b | |
| H | 12.32±0.65a | 7.53±0.05a | 0.44±0.02a | 0.19±0.05b | 5.35±0.23b | 0.47±0.10a | 0.18±0.02b | |
| 不同字母表示在5%的显著性水平下有差异(P<0.05), a、b、c表示同一研究区不同演替阶段间土壤理化性质的差异 | ||||||||
使用Qiime软件(Version 1.7.0)计算AMF多样性指数;运用SPSS18.0单因素方差分析比较乔、灌、草阶段之间多样性指数间的差异, Spearman相关性分析描述AMF多样性指数与土壤理化性质的相关性;采用Oringin8.0绘制柱状堆积图反映AM真菌丰度;运用R软件进行主成分分析(Principal Component Analysis, PCA)反映不同演替阶段的AMF群落组成差异, 典范对应分析(Canonical Correlation Analysis, CCA)反映土壤理化性质对AMF群落组成的影响。
2 结果与分析 2.1 采样点土壤AMF分类27个土壤样本共得到3871个OTU, 其中属于AMF的OTU为275个。Silva数据库进行比对共得到4目, 其中球囊霉目Glomerales的OTU为249, 多样孢囊霉目Diversisporales的OTU为14个, 原囊霉目Archaeosporales的OTU为10个, 类球囊霉目Paraglomerales的OTU为2个。从4个目中共得到8科, 13属及19个分子AMF分子种。其中斗管囊霉属Funneliformis 1种、根孢囊霉属Rhizophagus 1种、硬囊霉属Sclerocystis 1种、隔球囊霉属Septoglomus 1种、近明球囊霉属Claroideoglomus 3种、巨孢囊霉属Gigaspora 1种、盾巨孢囊霉属Scutellospora 1种、无梗囊霉属Acaulospora 2种、多样孢囊霉属Diversispora 3种、类球囊霉属Paraglomus 2种、双型囊霉属Ambispora 1种、原囊霉属Archaeospora 1种、内养囊霉属Entrophospora 1种[36] (表 3)。
| 目Orders | 科Families | 属Genera | 种Species |
| 球囊霉目Glomerales | 球囊霉科Glomeraceae | 斗管囊霉属 Funneliformis | 摩西斗管囊霉Funneliformis mosseae |
| 根孢囊霉属 Rhizophagus | 根内根孢囊霉Rhizophagus intraradices | ||
| 硬囊霉属 Sclerocystis | Sclerocystis sp.1 | ||
| 隔球囊霉属 Septoglomus | 缩隔球囊霉Septoglomus constrictum | ||
| 近明球囊霉科Claroideoglomeraceae | 近明球囊霉属Claroideoglomus | Claroideoglomus sp. MIB8381 | |
| Claroideoglomus sp. NBR_PP1 | |||
| Claroideoglomus sp. W3234 | |||
| 多样孢囊霉目Diversisporales | 巨孢囊霉科Gigasporaceae | 巨孢囊霉属 Gigaspora | 球状巨孢囊霉Gigaspora margarita |
| 盾巨孢囊霉属 Scutellospora | 双紫盾巨孢囊霉Scutellospora dipurpurescens | ||
| 无梗囊霉科Acaulosporaceae | 无梗囊霉属 Acaulospora | 巴西无梗囊霉Acaulospora brasliensis | |
| 稍长无梗囊霉Acaulospora longula | |||
| 多样孢囊霉科Diversisporaceae | 多样孢囊霉属 Diversispora | Diversispora sp. w2423 | |
| Diversispora sp. w4568 | |||
| 隐多样孢囊霉Diversispora celata | |||
| 类球囊霉目Paraglomerales | 类球囊霉科Paraglomeraceae | 类球囊霉属 Paraglomus | 巴西类球囊霉Paraglomus brasilianum |
| 隐类球囊霉Paraglomus occultum | |||
| 原囊霉目Archaeosporales | 双型囊霉科Ambisporaceae | 双型囊霉属 Ambispora | 芬兰双型囊霉Ambispora fennica |
| 原囊霉科Archaeosporaceae | 原囊霉属 Archaeospora | Archaeospora sp. MIB8442 | |
| 内养囊霉属 Entrophospora | 稀有内养囊霉Entrophospora infrequens |
花溪、花江、织金属水平上AMF丰度分别为:乔木>灌木>草地、灌木>乔木>草地、草地>灌木>乔木, 总体AMF丰度为花江>花溪>织金, 且花江各样地AMF丰度均高于其余各样地丰度。各样地优势属及次优势属均为根孢囊霉属和斗管囊霉属, 花溪灌木第三优势属为硬囊霉属, 花江灌木和草本第三优势属为隔球囊霉属, 其余样地第三优势属均为近明球囊霉属(图 1)。

|
| 图 1 AMF属水平OTU丰度 Fig. 1 The OTU abundance of AMF at the genus level AM:丛枝菌根, arbuscular mycorrhizal;OTU:操作分类单元, operational taxonomic units;HXT:花溪乔木演替阶段, HuaXi Tree;HXB:花溪灌木演替阶段, HuaXi Bush;HXH:花溪草本演替阶段, HuaXi Herb;HJT:花江乔木演替阶段, HuaJiang Tree;HJB:花江灌木演替阶段, HuaJiang Bush;HJH:花江草本演替阶段, HuaJiang Herb;ZJT:织金乔木演替阶段, ZhiJin Tree; ZJB:织金灌木演替阶段, ZhiJin Bush;ZJH:织金草本演替阶段, ZhiJin Herb |
摩西斗管囊霉和Sclerocystis sp. 1仅在花溪乔木、灌木、草地演替阶段分布, 可见是花溪特有种。缩隔球囊霉、根内根孢囊霉、Claroideoglomus sp. MIB8381和稀有内养囊霉在各个样地均有分布, 可见是喀斯特地区的常见种。稍长无梗囊霉丰度在各种中最小, 且仅在织金乔木及花江灌木演替阶段分布。花溪、织金种水平上AMF丰度均为乔木>灌木>草地、而花江为乔木>草地>灌木。总体种丰度趋势为:花江>花溪>织金(表 4)。
| AMF分子种 Molecular species of AMF |
HJ | ZJ | HX | 合计 Total |
||||||||
| T | B | H | T | B | H | T | B | H | ||||
| 摩西斗管囊霉 Funneliformis mosseae |
30 | 42 | 16 | 88 | ||||||||
| 根内根孢囊霉 Rhizophagus intraradices |
344 | 35 | 70 | 3 | 210 | 25 | 586 | 63 | 101 | 1437 | ||
| Sclerocystis sp. 1 | 146 | 28 | 338 | 512 | ||||||||
| 缩隔球囊霉 Septoglomus constrictum |
616 | 392 | 815 | 2 | 42 | 18 | 212 | 24 | 21 | 2142 | ||
| Claroideoglomus sp. MIB8381 | 666 | 25 | 1 | 61 | 3 | 2 | 61 | 310 | 13 | 1142 | ||
| Claroideoglomussp. NBR_PP1 | 8 | 1 | 1 | 59 | 1 | 5 | 75 | |||||
| Claroideoglomussp. W3234 | 25 | 8 | 7 | 3 | 1 | 62 | 103 | 17 | 226 | |||
| 稀有内养囊霉 Entrophospora infrequens |
505 | 11 | 85 | 17 | 1 | 32 | 140 | 3 | 6 | 800 | ||
| 球状巨孢囊霉 Gigaspora margarita |
9 | 14 | 3 | 26 | ||||||||
| 双紫盾巨孢囊霉 Scutellospora dipurpurescens |
3 | 11 | 5 | 23 | 6 | 3 | 51 | |||||
| 巴西无梗囊霉 Acaulospora brasliensis |
2 | 25 | 27 | |||||||||
| 稍长无梗囊霉 Acaulospora longula |
1 | 1 | 2 | |||||||||
| Diversispora sp. w2423 | 178 | 13 | 16 | 4 | 35 | 246 | ||||||
| Diversispora sp. w4568 | 9 | 3 | 18 | 7 | 6 | 1 | 7 | 2 | 53 | |||
| 隐多样孢囊霉 Diversispora celata |
10 | 89 | 13 | 11 | 123 | |||||||
| 巴西类球囊霉 Paraglomus brasilianum |
15 | 119 | 7 | 141 | ||||||||
| 隐类球囊霉 Paraglomus occultum |
2 | 4 | 6 | |||||||||
| 芬兰双型囊霉 Ambispora fennica |
10 | 57 | 6 | 73 | ||||||||
| Archaeospora sp. MIB8442 | 15 | 120 | 80 | 215 | ||||||||
| 合计Total | 2344 | 548 | 982 | 460 | 285 | 186 | 1447 | 600 | 533 | |||
所有采样点测序文库的覆盖度均达到90%以上, 说明绝大部分AMF序列可以被测出, 测序结果具有较好的代表性, 各样地AMF多样性指数均较高。在同一研究区不同演替阶段, 香农与辛普森指数变化趋势基本一致, 即在花溪和织金样点:乔木≈灌木>草地, 且花溪乔木和灌木与草地之间差异显著(P<0.05), 织金乔木与草地间差异显著(P<0.05), 说明花溪乔木和灌木AMF物种多样性和常见种优势度高于草地, 织金乔木AMF物种多样性和常见种优势度高于草地;在花江样点则为:草地>灌木>乔木, 但是差异不显著。花江乔、灌、草样地的AMF香农指数均大于花溪和织金草地的指数, 且差异显著(P<0.05), 说明花江AMF物种多样性大于花溪和织金草地演替阶段。Chao1和ACE指数基本呈现一致的变化趋势:花溪采样点AMF群落丰度为乔木>灌木>草地, 织金采样点AMF群落丰度为灌木>乔木>草地, 且差异均不显著;而花江采样点则为灌木≈草地>乔木, 且差异显著(P<0.05)。花江灌木和乔木演替阶段分别拥有各样地最高和最低的Chao1和ACE指数(表 5)。
| 采样点 Sample site |
97%相似水平Similarity of 97% | |||||
| 香农指数 Simpson index |
辛普森指数 Simpson index |
Chao1指数 Chao1 index |
ACE指数 ACE index |
覆盖度 Coverage |
||
| HX | T | 5.04±0.21aα | 0.95±0.006aα | 94.86±10.15aα | 100.33±11.42aα | 0.92±0.01aβ |
| B | 4.8±0.11aα | 0.94±0.004aα | 94.25±12.97aαβ | 100.71±13.93aα | 0.92±0.01aα | |
| H | 4.02±0.17bβ | 0.89±0.02bβ | 70.23±10.03bβ | 83.55±11.89aα | 0.94±0.01aαβ | |
| HJ | T | 4.39±0.19aβ | 0.92±0.011bβ | 56.22±9.65bγ | 59.64±10.34bβ | 0.96±0.01aα |
| B | 4.76±0.3aα | 0.93±0.015abα | 119.70±22.64aα | 108.67±16.98aα | 0.92±0.01bα | |
| H | 4.96±0.15aα | 0.95±0.011aα | 98.00±17.06aα | 86.43±6.24aα | 0.93±0.01abβ | |
| ZJ | T | 5.07±0.07aα | 0.95±0.005aα | 74.14±8.08aβ | 75.17±5.92aβ | 0.95±0.01aα |
| B | 4.83±0.31abα | 0.93±0.027abα | 78.81±8.63aβ | 83.72±10.02aα | 0.94±0.01aα | |
| H | 4.37±0.31bβ | 0.90±0.035bβ | 68.43±10.92aβ | 71.43±11.22aα | 0.95±0.00aα | |
| 不同字母表示在5%的显著性水平下有差异(P<0.05), a、b表示同一研究区不同演替阶段间AMF丰富度与多样性的差异, α, β, γ表示同一演替阶段不同研究区间AMF丰富度与多样性的差异;ACE:ACE指数, abundance-based coverage estimation index | ||||||
在同一演替阶段不同研究区, 乔木演替阶段的AMF香农与辛普森指数变化趋势为花溪≈织金>花江;草本演替阶段为花江>花溪≈织金(P<0.05);灌木演替阶段基本一致。乔木演替阶段AMF Chao1和ACE指数为花溪>花江≈织金(P<0.05), 灌木和草本演替阶段均为花江>花溪≈织金, Chao1指数差异显著(P<0.05), ACE指数差异不显著(表 5)。
2.4 不同植被演替阶段AMF群落主成分分析PCA分析表明各采样点除织金乔木与草地演替阶段外, 其余各采样点重复间的群落组成均差异较小。在同一研究区不同演替阶段:花江草地与灌木演替阶段AMF群落组成差异小, 而乔木与草地和灌木间AMF群落组成差异大;花溪乔木与灌木演替阶段AMF群落组成差异小, 与草地演替阶段差异大;织金乔、灌、草演替阶段AMF群落组成差异均较大。在同一演替阶段不同研究区:各研究区的乔木、灌木及草地演替阶段均相隔较远的距离, 表明同一演替阶段的不同研究区AMF群落组成均有较大差异(图 2)。

|
| 图 2 不同演替阶段PCA分析 Fig. 2 Analysis of PCA in different stage of succession |
AMF多样性指数与环境因子的Spearman相关性分析表明:土壤全磷与ACE指数显著负相关(P<0.05), 且与Chao1指数极显著负相关(P<0.01);速效磷与香农和辛普森指数显著负相关(P<0.05);速效钾与各多样性指数呈现正相关关系, 全氮和有机质与各多样性指数呈现负相关关系, 但差异均不显著(表 6)。
| pH | 全磷 Total phosphorus |
全氮 Total nitrogen |
速效磷 Available phosphorus |
速效钾 Available kalium |
水解氮 Hydrolysis nitrogen |
有机质 Organic matter |
|
| 香农指数 Simpson index |
-0.018 | -0.009 | -0.296 | -0.429* | 0.068 | 0.004 | -0.128 |
| 辛普森指数Simpson index | -0.063 | 0.142 | -0.253 | -0.425* | 0.015 | 0.093 | -0.212 |
| Chao1指数Chao1 index | 0.380 | -0.510** | -0.230 | -0.062 | 0.206 | -0.106 | -0.094 |
| ACE指数ACE index | 0.280 | -0.474* | -0.238 | 0.082 | 0.317 | -0.005 | -0.136 |
由土壤AMF OTU与理化性质的典型典范对应分析结果排序图可知:第一主轴和第二主轴对AMF群落分布方差的解释比例分别为30.4%和20.1%, 两者共解释50.5%的方差变化。在第一主轴上, 土壤pH、全磷和速效钾为主要影响因子(相关系数为0.976、0.971、0.888);在第二主轴上, 土壤全氮、有机质、水解氮和速效磷为主要影响因子(相关系数分别为0.993、-0.887、-0.883和-0.812)。除土壤pH与速效磷外, 其余理化性质均与AMF群落分布有显著相关性, 其中有机质为显著相关(P<0.05), 其余为极显著相关(P<0.01)。土壤全氮与其余理化性质为负相关关系(图 3)。

|
| 图 3 AMF群落与土壤理化性质CCA分析 Fig. 3 Canonical Correlation Analysis between AMF community and soil physicochemical property TN:全氮, Total nitrogen;TP:全磷, Total phosphorus;AK:速效钾, Available kalium;AP:速效磷, Available phosphorus;HN:水解氮, Hydrolysis nitrogen;OM:有机质, Organic matter |
本研究从基因水平上考察了3个典型喀斯特区域不同自然演替阶段的AMF多样性, 结果表明各样地均有较高的AMF多样性, 种属水平的AMF丰度变化趋势均为花江>花溪>织金, 且各样地优势属均为根孢囊霉属, 该结果与本研究所用形态学方法及大多数文献所报道的结果有一定的差异[13]。魏源等[15]利用巢式PCR与变性梯度凝胶电泳相结合的分子生物学方法, 表明球囊霉属(Glomus)极有可能是喀斯特地区AMF的优势菌属, Likar等[37]研究喀斯特土壤中的AMF多样性及其分布, 同样发现球囊霉属是喀斯特生境中稳定存在的优势属。有学者曾在其研究中分别采用形态学鉴定、定量PCR与PCR-DGGE及454高通量测序技术对长期不同施肥作用下的土壤AMF群落结构及物种多样性进行研究, 结果发现不同方法所得的次优势属种以及所鉴定的种属数目均有所差异, 因此, 产生差异的原因可能与不同方法本身的优劣性及分辨率有一定的关系[38]。
AMF通过菌丝网络调节植被群落结构及演替, 从而影响生态系统稳定及功能[39]。刘永俊等[40]及Zangrao等[41]研究结果均表明:不同演替阶段AMF多样性会随着植物演替进行而降低, 与本研究中花江研究区AMF多样性变化趋势一致(表 4), 而花溪和织金研究区的AMF多样性却随着演替进行而增加。其原因可能是:(1)本文的研究对象是自然演替过程中的土壤AMF多样性, 植物群落及多样性均有较大差异(表 1), 而刘永俊等研究反映的是以柠条为优势物种的人工植被演替生态系统中侵染了柠条根系的AMF多样性, 植物多样性则十分匮乏, 钟凯等[42]对泰山植被根围AMF群落结构、数量、组成及植物多样性研究发现, 植物多样性对于提高AMF多样性发挥极为重要的作用, Hiiesalu等[43]在其研究中也指出AMF物种丰度与植物物种丰度呈显著正相关关系。(2)Zangrao等采用形态学方法研究AMF多样性, 但不同种类AMF的产孢能力不同, 且AMF的侵染能力也存在很大差异, 可能不能真实地反映土壤中的AMF多样性[38]。本研究所选3个研究区所呈现的不同演替阶段AMF多样性变化趋势不一, 其原因可能与各研究区有一定的地理距离相关, Xu等[44]研究结果表明地理距离解释了AMF的系统发育格局, AMF群落组成会受到扩散限制值的影响。
已有研究表明, AMF丰度及多样性会随着土壤养分的供应情况发生变化, 土壤理化性质对AMF群落结构及多样性有重要影响[45-46]。本研究结果显示土壤全磷与AMF丰富度指数显著负相关, 速效磷与AMF多样性指数显著负相关, 表明土壤全磷和速效磷是影响AMF丰度和多样性的主导因子, 该结果与梁月明等[47-48]研究结果一致, 而杨春雪等[49]研究表明土壤全磷与AMF物种丰度呈显著正相关关系, 有学者曾指出, 一定范围内的低量速效磷会促进AMF的生长, 而过高的速效磷含量会抑制AMF的生长及发育[50-51]。CCA分析表明土壤全氮、水解氮、有机质、全磷和速效钾对AMF群落影响差异显著, pH却差异不显著, 该结果与张玉洁等[52]认为土壤pH为影响AMF多样性的主要土壤因子不一致, 导致该结果的原因可能与土壤本身固有属性有关, 成土母质及元素组成决定了pH值的相似性, 在同一区域的土壤类型, 其pH值空间变异性较低[53], 从而对AMF多样性没有显著影响。
| [1] |
Wang B, Qiu Y L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza, 2006, 16(5): 299-363. DOI:10.1007/s00572-005-0033-6 |
| [2] |
Smith S E, Read D J. Mycorrhizal Symbiosis. London: Academic Press, 1997: 1-20.
|
| [3] |
van der Heijden M G A, Bardgett R D, van Straalen N M. The unseen majority:soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters, 2010, 11(3): 296-310. |
| [4] |
刘敏, 峥嵘, 白淑兰, 王琚钢, 李龙, 段国珍. 丛枝菌根真菌物种多样性研究进展. 微生物学通报, 2016, 43(8): 1836-1843. |
| [5] |
彭旭东, 戴全厚, 李昌兰. 中国西南喀斯特坡地水土流失/漏失过程与机理研究进展. 水土保持学报, 2017, 31(5): 1-8. |
| [6] |
宋同清, 彭晚霞, 杜虎, 王克林, 曾馥平. 中国西南喀斯特石漠化时空演变特征、发生机制与调控对策. 生态学报, 2014, 34(18): 5328-5341. |
| [7] |
陈圣子, 周忠发, 闫利会. 人为干预对石漠化治理过程中生态系统健康的影响. 水土保持研究, 2016, 23(2): 213-219. |
| [8] |
邢丹, 王震洪, 张爱民, 付文婷, 韩世玉. 菌根桑恢复喀斯特石漠化地区的研究探讨(英文). Agricultural Science & Technology, 2014, 15(11): 1998-2002. DOI:10.3969/j.issn.1009-4229.2014.11.037 |
| [9] |
Chen K, Shi S M, Yang X H, Huang X Z. Contribution of arbuscular mycorrhizal inoculation to the growth and photosynthesis of mulberry in Karst Rocky Desertification Area. Applied Mechanics and Materials, 2014, 488(-489): 769-773. |
| [10] |
杨应, 蒋长洪, 何跃军, 欧静, 王鹏鹏, 司建朋, 何敏红, 林艳. 丛枝菌根网对喀斯特适生植物氮、磷化学计量特征的影响. 植物生理学报, 2017, 53(12): 2078-2090. |
| [11] |
吴春玉, 蒋长洪, 谢佩耘, 何跃军, 杨俊松. 干旱胁迫对接种AMF的香樟幼苗生物量分配及生长的影响. 重庆师范大学学报:自然科学版, 2015, 32(6): 109-115. |
| [12] |
何跃军, 钟章成. 喀斯特土壤上香樟幼苗接种不同AM真菌后的耐旱性效应. 植物研究, 2011, 31(5): 597-602. |
| [13] |
何跃军, 吴春玉, 何丙辉, 丁贵杰, 孙学广, 高秀兵. 喀斯特不同植被恢复阶段土壤AMF组成及多样性研究. 水土保持学报, 2016, 30(5): 305-309, 321-321. |
| [14] |
魏源, 王世杰, 刘秀明, 黄天志. 喀斯特地区丛枝菌根真菌遗传多样性. 生态学杂志, 2011, 30(10): 2220-2226. |
| [15] |
魏源, 王世杰, 刘秀明, 黄天志. 不同喀斯特小生境中土壤丛枝菌根真菌的遗传多样性. 植物生态学报, 2011, 35(10): 1083-1090. |
| [16] |
梁月明, 苏以荣, 何寻阳, 陈香碧. 岩性对喀斯特灌丛土壤固氮菌与丛枝菌根真菌群落结构及丰度的影响. 环境科学, 2017, 38(3): 1253-1261. |
| [17] |
魏源, 王世杰, 刘秀明, 黄天志. 丛枝菌根真菌及在石漠化治理中的应用探讨. 地球与环境, 2012, 40(1): 84-92. |
| [18] |
Barni E, Siniscalco C. Vegetation dynamics and arbuscular mycorrhiza in old-field successions of the western Italian Alps. Mycorrhiza, 2000, 10(2): 63-72. DOI:10.1007/s005720000059 |
| [19] |
Roy J, Reichel R, Brüggemann N, Hempel S, Rillig M C. Succession of arbuscular mycorrhizal fungi along a 52-year agricultural recultivation chronosequence. FEMS Microbiology Ecology, 2017, 93(9): 1-13. |
| [20] |
de León D G, Moora M, Öpik M, Neuenkamp L, Gerz M, Jairus T, Vasar M, Bueno C G, Davison J, Zobel M. Symbiont dynamics during ecosystem succession:co-occurring plant and arbuscular mycorrhizal fungal communities. FEMS Microbiology Ecology, 2016, 92(7): 1-9. |
| [21] |
Xiang D, Chen B D, Li H. Specificity and selectivity of arbuscular mycorrhizal fungal polymerase chain reaction primers in soil samples by clone library analyses. Acta Agriculturae Scandinavica, Section B - Soil & Plant Science, 2015, 66(4): 333-339. |
| [22] |
De Beenhouwer M, Van Geel M, Ceulemans T, Muleta D, Lievens B, Honnay O. Changing soil characteristics alter the arbuscular mycorrhizal fungi communities of Arabica coffee (Coffea arabica) in Ethiopia across a management intensity gradient. Soil Biology and Biochemistry, 2015, 91: 133-139. DOI:10.1016/j.soilbio.2015.08.037 |
| [23] |
Liang Y M, He X Y, Chen C Y, Feng S Z, Liu L, Chen X B, Zhao Z W, Su Y R. Influence of plant communities and soil properties during natural vegetation restoration on arbuscular mycorrhizal fungal communities in a karst region. Ecological Engineering, 2015, 82: 57-65. DOI:10.1016/j.ecoleng.2015.04.089 |
| [24] |
Van Geel M, Ceustermans A, Van Hemelrijck W, Lievens B, Honnay O. Decrease in diversity and changes in community composition of arbuscular mycorrhizal fungi in roots of apple trees with increasing orchard management intensity across a regional scale. Molecular Ecology, 2015, 24(4): 941-952. DOI:10.1111/mec.13079 |
| [25] |
Dai M L, Hamel C, Bainard L D, Arnaud M S, Grant C A, Lupwayi N Z, Malhi S S, Lemke R. Negative and positive contributions of arbuscular mycorrhizal fungal taxa to wheat production and nutrient uptake efficiency in organic and conventional systems in the Canadian prairie. Soil Biology and Biochemistry, 2014, 74: 156-166. DOI:10.1016/j.soilbio.2014.03.016 |
| [26] |
Magoč T, Salzberg S L. FLASH:fast length adjustment of short reads to improve genome assemblies. Bioinformatics, 2011, 27(21): 2957-2963. DOI:10.1093/bioinformatics/btr507 |
| [27] |
Bokulich N A, Subramanian S, Faith J J, Gevers D, Gordon J I, Knight R, Mills D A, Caporaso J G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nature Methods, 2013, 10(1): 57-59. DOI:10.1038/nmeth.2276 |
| [28] |
Caporaso J G, Kuczynski J, Stombaugh J, Bittinger K, Bushman F D, Costello E K, Fierer N, Peña A G, Goodrich J K, Gordon J I, Huttley G A, Kelley S T, Knights D, Koenig J E, Ley R E, Lozupone C A, McDonald D, Muegge B D, Pirrung M, Reeder J, Sevinsky J R, Turnbaugh P J, Walters W A, Widmann J, Yatsunenko T, Zaneveld J, Knight R. QⅡME allows analysis of high-throughput community sequencing data. Nature Methods, 2010, 7(5): 335-336. DOI:10.1038/nmeth.f.303 |
| [29] |
Edgar R C, Haas B J, Clemente J C, Quince C, Knight R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics, 2011, 27(16): 2194-2200. DOI:10.1093/bioinformatics/btr381 |
| [30] |
Haas B J, Gevers D, Earl A M, Feldgarden M, Ward D V, Giannoukos G, Ciulla D, Tabbaa D, Highlander S K, Sodergren E, Methé B, DeSantis T Z, Petrosino J F, Knight R, Birren B W. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Research, 2011, 21(3): 494-504. DOI:10.1101/gr.112730.110 |
| [31] |
Edgar R C. UPARSE:highly accurate OTU sequences from microbial amplicon reads. Nature Methods, 2013, 10(10): 996-998. DOI:10.1038/nmeth.2604 |
| [32] |
Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. Journal of Molecular Biology, 1990, 215(3): 403-410. DOI:10.1016/S0022-2836(05)80360-2 |
| [33] |
Kõljalg U, Nilsson R H, Abarenkov K, Tedersoo L, Taylor A F S, Bahram M, Bates S T, Bruns T D, Bengtsson-Palme J, Callaghan T M, Douglas B, Drenkhan T, Eberhardt U, Dueñas M, Grebenc T, Griffith G W, Hartmann M, Kirk P M, Kohout P, Larsson E, Lindahl B D, Lücking R, Martín M P, Matheny P B, Nguyen N H, Niskanen T, Oja J, Peay K G, Peintner U, Peterson M, Põldmaa K, Saag L, Saar I, Schüßler A, Scott J A, Senés C, Smith M E, Suija A, Taylor D L, Telleria M T, Weiss M, Larsson K H. Towards a unified paradigm for sequence-based identification of fungi. Molecular Ecology, 2013, 22(21): 5271-5277. DOI:10.1111/mec.12481 |
| [34] |
Edgar R C. MUSCLE:multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 2004, 32(5): 1792-1797. DOI:10.1093/nar/gkh340 |
| [35] |
张韫. 土壤·水·植物理化分析教程. 北京: 中国林业出版社, 2011.
|
| [36] |
王幼珊, 刘润进. 球囊菌门丛枝菌根真菌最新分类系统菌种名录. 菌物学报, 2017, 36(7): 820-850. |
| [37] |
Likar M, Hančević K, Radić T, Regvar M. Distribution and diversity of arbuscular mycorrhizal fungi in grapevines from production vineyards along the eastern Adriatic coast. Mycorrhiza, 2013, 23(3): 209-219. DOI:10.1007/s00572-012-0463-x |
| [38] |
林先贵, 胡君利, 戴珏, 王发园, 冯有智. 丛枝菌根真菌群落结构与多样性研究方法概述及实例比较. 应用与环境生物学报, 2017, 23(2): 343-350. |
| [39] |
向丹, 徐天乐, 李欢, 陈保冬. 丛枝菌根真菌的生态分布及其影响因子研究进展. 生态学报, 2017, 37(11): 3597-3606. |
| [40] |
刘永俊, 冯虎元. 不同演替阶段人工柠条林丛枝菌根真菌分子多样性研究. 中国沙漠, 2009, 29(6): 1141-1147. |
| [41] |
Zangaro W, Alves R A, Lescano L E, Ansanelo A P, Nogueira M A. Investment in fine roots and arbuscular mycorrhizal fungi decrease during succession in Three Brazilian Ecosystems. Biotropica, 2012, 44(2): 141-150. DOI:10.1111/j.1744-7429.2011.00781.x |
| [42] |
钟凯, 袁玉清, 赵洪海, 王淼焱, 刘润进. 泰山丛枝菌根真菌群落结构特征. 菌物学报, 2010, 29(1): 44-50. |
| [43] |
Hiiesalu I, Pärtel M, Davison J, Gerhold P, Metsis M, Moora M, Öpik M, Vasar M, Zobel M, Wilson S D. Species richness of arbuscular mycorrhizal fungi:associations with grassland plant richness and biomass. New Phytologist, 2014, 203(1): 233-244. DOI:10.1111/nph.12765 |
| [44] |
Xu T L, Veresoglou S D, Chen Y L, Rillig M C, Xiang D, Ondřej X D, Hao Z P, Liu L, Deng Y, Hu Y J, Chen W P, Wang J T, He J Z, Chen B D. Plant community, geographic distance and abiotic factors play different roles in predicting AMF biogeography at the regional scale in northern China. Environmental Microbiology Reports, 2016, 8(6): 1048-1057. DOI:10.1111/emi4.2016.8.issue-6 |
| [45] |
Alguacil M M, Torrecillas E, Lozano Z, Roldán A. Arbuscular mycorrhizal fungi communities in a coral cay system (Morrocoy, Venezuela) and their relationships with environmental variables. Science of the Total Environment, 2015, 505: 805-813. DOI:10.1016/j.scitotenv.2014.10.030 |
| [46] |
Xiang D, Verbruggen E, Hu Y J, Veresoglou S D, Rillig M C, Zhou W P, Xu T L, Li H, Hao Z P, Chen Y L, Chen B D. Land use influences arbuscular mycorrhizal fungal communities in the farming-pastoral ecotone of northern China. New Phytologist, 2014, 204(4): 968-978. DOI:10.1111/nph.12961 |
| [47] |
梁月明, 苏以荣, 何寻阳, 陈香碧, 胡亚军. 喀斯特灌丛土壤丛枝菌根真菌群落结构及丰度的影响因子. 环境科学, 2017, 38(11): 4828-4835. |
| [48] |
Gosling P, Mead A, Proctor M, Hammond J P, Bending G D. Contrasting arbuscular mycorrhizal communities colonizing different host plants show a similar response to a soil phosphorus concentration gradient. New Phytologist, 2013, 198(2): 546-556. DOI:10.1111/nph.12169 |
| [49] |
杨春雪, 陈飞, 岳英男, 阎秀峰. 松嫩盐碱草地26种植物根围丛枝菌根真菌多样性特征. 草业科学, 2015, 32(12): 2008-2020. DOI:10.11829/j.issn.1001-0629.2015-0232 |
| [50] |
Tawaraya K, Saito M, Morioka M, Wagatsuma T. Effect of phosphate application to arbuscular mycorrhizal onion on the development and succinate dehydrogenase activity of internal hyphae. Soil Science and Plant Nutrition, 1994, 40(4): 667-673. DOI:10.1080/00380768.1994.10414306 |
| [51] |
Deepika S, Kothamasi D. Soil moisture-a regulator of arbuscular mycorrhizal fungal community assembly and symbiotic phosphorus uptake. Mycorrhiza, 2015, 25(1): 67-75. DOI:10.1007/s00572-014-0596-1 |
| [52] |
张玉洁, 贺学礼, 程春泉, 赵金莉, 赵丽莉. 新疆沙冬青AM真菌群落结构与遗传多样性分析. 菌物学报, 2015, 34(3): 375-385. |
| [53] |
高鹏, 付同刚, 王克林, 陈洪松, 曾馥平. 喀斯特峰丛洼地小流域表层土壤养分的空间异质性. 农业现代化研究, 2013, 34(3): 362-366. |