 2019, Vol. 39
2019, Vol. 39文章信息
- 张维伟, 薛文艳, 杨斌, 赵忠
- ZHANG Weiwei, XUE Wenyan, YANG Bin, ZHAO Zhong
- 桥山栎林群落结构特征与物种多样性相关关系分析
- Differences in community characteristics, species diversity, and their coupling associations among three Quercus acutissima forests in Qiaoshan
- 生态学报. 2019, 39(11): 3991-4001
- Acta Ecologica Sinica. 2019, 39(11): 3991-4001
- http://dx.doi.org/10.5846/stxb201804200901
-
文章历史
- 收稿日期: 2018-04-20
- 网络出版日期: 2019-03-21

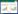




植物群落是生活在一定区域内所有植物个体通过互惠、竞争等相互作用而形成的一个有机整体, 不同类型群落在结构和功能上具有很大的差异[1-2]。树高、胸径、枝下高、冠幅、盖度等是描述群落结构特征的常用指标[3-4], 物种多样性则是群落功能的主要体现[5]。因此, 阐明群落结构特征与物种多样性的关系对于认识群落结构与功能及其相互关系意义重大, 研究群落结构特征为对天然林的合理经营管理及物种多样性的保育提供依据[6-7]。
麻栎(Quercus acutissima)属壳斗科栎属乔木, 是黄土高原南部桥山林区阔叶落叶林地带性植被的建群种[8], 常以纯林、麻栎阔叶混交林、麻栎油松混交林的形式构成当地最重要森林景观, 在当地涵养水源、保持水土、生物多样性保护等方面发挥着重要作用[9-10]。多年来, 由于不合理经营及乱砍滥伐, 导致当地栎林遭到严重破坏, 逐渐生长为残败次生林, 林分质量及林内物种多样性均不高。为实现黄土高原森林质量提升及生态功能发挥的目标, 最主要是研究清楚群落结构特征和物种多样性的关系, 但迄今为止, 尚未见到有关黄土高原南部麻栎林及其物种多样性关系的研究, 而这些正是麻栎林持续经营和管理的基础, 也是亟待解决的问题。
本文以麻栎林发育完整, 结构类型较多的桥山林区为基地, 选择生境相似、林龄相近的麻栎天然纯林、麻栎阔叶混交林和麻栎油松混交林为对象, 通过群落结构特征、物种多样性系统调查, 采用RDA冗余分析方法, 对群落结构特征与物种多样性的关系进行分析, 比较不同类型麻栎林群落结构特征与物种多样性相关关系, 目的是阐明不同类型麻栎林的结构特征及其与物种多样性关系, 为合理经营及物种多样性保育提供依据。
1 研究区概况本研究区域选择在黄土高原麻栎发育最好的陕西省延安市桥山林业局店头林场及双龙林场, 地理位置108°91′—109°12′E, 35°64′—35°72′N, 海拔900—1500 m, 属暖温带气候区, 年平均降水量为683.2 mm, 平均气温10.2℃。土壤类型主要为森林褐土和灰褐土。麻栎林是地带性植被类型, 麻栎纯林(Pure forest of Q.acutissima, PFQ)、麻栎阔叶混交林(Mixed forest of Q. Acutissima with broad leaved tree, MFQB)、麻栎油松混交林(Mixed forest of Q. Acutissima with Pinus tabulaeformis, MFQP)是麻栎分布区最重要的3种林分, 占桥山林区森林类型的60%。PFQ主要乔木树种为麻栎, 占乔木组成90%以上, 主要灌木为狼牙刺(Sophora viciifolia)、南蛇藤(Celastrus orbiculatus)。MFQB乔木树种以麻栎(Quercus acutissima)为主, 其他阔叶树有辽东栎(Quercus wutaishansea)、槲栎(Quercus aliena)、茶条槭(Acer ginnala)等, 主要灌木树种为胡枝子(Lespedeza bicolor)、狼牙刺、黄蔷薇(Rosa hugonis)。MFQP乔木树种以麻栎(Q. acutissima)和油松(Pinus tabulaeformis)为主, 主要灌木树种为南蛇藤、胡枝子、忍冬(Lonicera japonica)。3种林分最主要草本植物皆为苔草(Carex tristachya)。
2 研究方法 2.1 样地设置与调查样地设置采用典型选样的办法, 2017年7月—8月在店头林场、双龙林场选择麻栎林分布较为集中的地区, 在充分探查的基础上在阴坡选择林相相对整齐的地段作为样地, 共计45块。其中PFQ样地15块、MFQB样地15块、MFQP样地15块。样地面积均为20 m×30 m, 每个样地沿对角线于左下、中、右上设置灌木样方3块, 面积均为5 m×5 m;同时在每个灌木样方的左下角设置草本样方, 面积均为1 m×1 m。在样地周边设置1 m×20 m的样方带, 用于测定每一物种的频度。
这3种类型的样地区域规划为薪炭林, 在20世纪50—60年代, 经过持续砍伐沦为残败次生林, 稀疏分布林木个体直径约7—8 cm。1965年以后处于封禁状态。1985年左右进行过一次砍灌及强度为20%的抚育间伐。过后一直处于封禁状态。
乔木调查:每木需坐标定位(沿地表等高线为X轴, 垂直边为Y轴), 测定并记录每个林木的种名、胸径、树高、冠幅、枝下高及新稍长。灌木层调查:在灌木样方中分物种测定每一植株高度、地径、个体数、冠幅、盖度;草本层调查:在草本样方中, 分物种测定株高度、基径、个体数、冠幅、盖度。沿频度样方带, 测定每一乔灌草物种的频数, 并计算频度。
2.2 物种多样性计算[11]物种重要值、多样性指数等参数均以样地为单位统计, 同类型样地合并后取平均值作为该群落类型的特征参数。
(1) 重要值
乔木层重要值= (相对密度+相对显著度+相对频度)/3
灌木层重要值= (相对密度+相对盖度+相对频度)/3
草本层重要值= (相对密度+相对盖度+相对频度)/3
相对密度=该种的所有株数/所有种的株数之和×100%
相对显著度=该种个体胸高断面积/所有中的胸高断面积之和×100%
相对盖度=该种的盖度/所有种的盖度之和×100%
相对频度=该种的频度/所有种的频度之和×100%
(2) 物种丰富度指数
物种丰富度即为样地内乔、灌、草物种的数目, 以同一群落类型物种丰富度, 以各样地物种数的平均值表示。
(3) Simpson多样性指数

|
式中, Ni为样地中第i物种的个体数, N为样地中所有物种的个体种数, S为物种数。
(4) Shannon-wiener指数
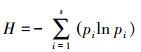
|
式中, pi为样地中第i物种的重要值, 即pi=Ni/N, S为物种数。
(5) Pielou指数
|
式中, Hmax=lnS, H′是样地的物种多样性实测值Hmax为样地中给定物种数S下完全均匀群落的多样性。
(6) Alatalo均匀度指数

|
各类型群落结构特征参数以样地为单位进行统计, 同类样地合并后取平均值为该群落类型的参数。运用SPSS 19.0对不同群落类型林分特征差异及多样性指数差异进行单因素方差分析, 若差异显著, 则进行Duncan多重比较, 显著性水平α=0.05。
运用R语言Vegan包进行排序分析[12]。在排序分析之前, 用decorana()函数判断应使用单峰模型(CA和CCA)或线性模型(PCA和RDA)[13]。本文中Axis lengths < 3, 因此用RDA()函数对群落结构特征与物种多样性进行约束排序[14]。
3 结果与分析 3.1 三种林分群落结构特征差异分析群落结构特征最重要的参数是群落组成、层次结构及其组分参数。表 1展示了3种类型麻栎群落特征及组成、结构和相关参数差异。MFQP乔木层树高、胸径均显著高于PFQ与MFQB(P < 0.05), PFQ与MFQB差异不显著;冠幅表现为MFQB>MFQP>PFQ;新稍长则正好相反, 以PFQ最大, MFQP最小。
| 层Layer | 特征参数 Characteristic index |
PFQ | FQB | MFQP |
| 乔木层Tree | 树高TH/m | 9.687b | 9.250b | 9.833a |
| 胸径TDBH/cm | 12.827b | 13.219b | 13.827a | |
| 枝下高TCBH/m | 3.595a | 2.878b | 3.687a | |
| 冠幅TCD/m | 3.567b | 4.206a | 3.873ab | |
| 新稍长TNTL/cm | 15.146a | 14.007b | 13.548b | |
| 灌木层Shrub | 株高SH/cm | 58.453b | 83.322a | 63.185b |
| 盖度SC/% | 0.060a | 0.101a | 0.041a | |
| 地径SGD/cm | 5.053b | 7.123a | 4.738b | |
| 冠幅SCD/cm | 45.139b | 63.446a | 48.593b | |
| 草本层Herb | 基径HD/mm | 2.382a | 1.551a | 1.292a |
| 盖度HC/% | 0.106a | 0.058b | 0.078b | |
| 高度HH/cm | 23.255b | 29.266a | 22.621b | |
| 冠幅HCD/cm | 24.548a | 23.182a | 24.933a | |
| TH:乔木树高, tree height; TNTL:乔木枝下高, the new twig length; TDBH:乔木胸径, tree diameter breast height; TCBH:乔木枝下高, tree clear tree bole height; SH:灌木株高, shrub height; SC:灌木盖度, shrub coverage; SGD:灌木地径, shrub ground diameter; SCD:灌木冠幅, shrub crown width; HA:草本多度, herb abundance; HC:草本盖度, herb coverage; HH:草本高度, herb height。同列中不同小写字母表示差异显著(P<0.05);PFQ(麻栎纯林Pure forest of Q. acutissima)、MFQB(麻栎阔叶混交林Mixed forest of Q. acutissima with broad leaved tree)、MFQP (麻栎油松混交林Mixed forest of Q. acutissima with Pinustabulaeformis)(下同Here in after the same) | ||||
灌木层株高与冠幅在3种类型林分间呈现出一致规律, 即MFQB>MFQP>PFQ, 地径则呈现出MFQB>PFQ>MFQP的趋势。MFQB灌木层盖度最高, 但3种林分类型间差异不显著(P>0.05)。
草本层基径、冠幅均呈现出PFQ>MFQB>MFQP。PFQ草本层盖度较MFQB高82.76%, 较MFQP高35.90%;MFQB草本层平均高度最大, 较PFQ高25.84%, 较MFQP高29.38%。
从对3种类型栎林结构的比较中可以看出, MFQP属当地典型地带性植被类型, 对当地土壤气候的适合程度优于MFQB和PFQ, 乔木层林木胸径、树高等指标达到最优状态。MFQB与MFQP比较, 统计上没有显著性差异, 但优于PFQ。
3.2 重要值分析群落物种组成是群落主要特征。3种麻栎群落中, 在PFQ乔木层中, 共统计到物种6个(重要值大于1, 下同), 其中麻栎重要值最高(重要值88.75), 共计988株, 其余树种如辽东栎、茶条槭、杜梨等零星存在, 重要值介于1.05—3.40之间。灌木层中重要值最高的物种是南蛇藤(17.74)及狼牙刺(17.48)。MFQB乔木层中, 物种8个, 以麻栎重要值最高, 重要值达到37.67, 总株数409株。灌木层中重要值最高的物种是胡枝子(21.11)及狼牙刺(18.64)。MFQP乔木层中, 共统计到9个物种, 以麻栎、油松的重要值最大, MFQP灌木层中胡枝子与南蛇藤重要值最高, 分别达到11.99与18.24。3种类型麻栎林的草本重要值最高的物种均为苔草, 其值介于69.34—81.51之间(表 2)。
| 种名 Species |
乔木层Tree layer | 灌木层Shrub layer | 草本层Herb layer | ||||||||
| PFQ | MFQB | MFQP | PFQ | MFQB | MFQP | PFQ | MFQB | MFQP | |||
| 麻栎Quercus acutissima Carruth | 88.75 | 37.67 | 32.91 | ||||||||
| 油松Pinus tabulaeformis Carr. | 20.92 | ||||||||||
| 茶条槭Acer ginnala | 2.03 | 3.77 | 2.21 | ||||||||
| 辽东栎Quercus wutaishansea Mary | 3.40 | 26.31 | 26.20 | ||||||||
| 槲栎Quercus aliena Bl. | 1.13 | 19.34 | 5.07 | ||||||||
| 山楂Crataegus pinnatifida Bunge | 1.20 | ||||||||||
| 毛梾Swida walteri (Wanger.) Sojak | 1.13 | 1.69 | 3.13 | ||||||||
| 白桦Betulapla typhylla Suk | 2.55 | 3.38 | |||||||||
| 山杏Armeniaca sibirica (L.) Lam. | 2.73 | 0.64 | |||||||||
| 杜梨Pyrus betulifolia Bunge | 1.05 | 3.72 | 1.18 | ||||||||
| 忍冬Lonicera japonica Thunb | 10.95 | 7.36 | 10.58 | ||||||||
| 胡颓子Elaeagnus pungensThunb. | 1.54 | 1.14 | 2.33 | ||||||||
| 荚蒾Viburnum dilatatum Thunb | 7.55 | 7.05 | 9.18 | ||||||||
| 胡枝子Lespedeza bicolor Turcz | 6.44 | 21.11 | 11.99 | ||||||||
| 红麸杨Rhus punjabensis var. sinica | 0.92 | 0.93 | 12.83 | 2.57 | 3.34 | ||||||
| 卫矛Euonymus alatus | 1.91 | 0.98 | 4.94 | ||||||||
| 悬钩子Rubus corchorifolius L.f. | 0.97 | 0.88 | 0.95 | ||||||||
| 菝葜Smilax china L. | 1.71 | 2.67 | 1.41 | ||||||||
| 南蛇藤Celastrus orbiculatus Thunb. | 17.74 | 10.14 | 18.24 | ||||||||
| 蛇葡萄Ampelopsis glandulosa | 1.28 | 1.73 | 3.08 | ||||||||
| 绣线菊Spiraea Salicifolia L. | 2.06 | 3.94 | 7.39 | ||||||||
| 栒子 Cotoneaster zabelii | 0.93 | 4.42 | 1.70 | ||||||||
| 狼牙刺Sophora viciifolia | 17.48 | 18.64 | 10.05 | ||||||||
| 黄蔷薇Rosa hugonis Hemsl. | 7.68 | 11.14 | 9.94 | ||||||||
| 苦参Sophora flavescens | 2.30 | 1.33 | |||||||||
| 杠柳Periploca sepium | 1.72 | ||||||||||
| 鼠李Rhamnus davurica Pall. | 1.25 | ||||||||||
| 红瑞木Swida alba Opiz | 5.62 | 4.09 | |||||||||
| 苔草Carex tristachya | 81.51 | 69.34 | 79.49 | ||||||||
| 茜草Rubia cordifolia L. | 2.04 | 4.19 | 1.45 | ||||||||
| 银兰Cephalanthera erecta Bl. | 5.10 | 3.70 | 2.44 | ||||||||
| 披碱草Elymus dahuricus Turcz. | 3.29 | 7.54 | 4.42 | ||||||||
| 白茅草Imperata cylindrica | 2.79 | 7.37 | 2.82 | ||||||||
| 薯蓣Dioscorea opposita | 1.82 | 1.27 | 1.41 | ||||||||
| 败酱Patrinia scabiosaefolia Fisch. ex Trev | 1.77 | ||||||||||
| 荩草Arthraxon hispidus (Thunb.) Makino | 1.13 | ||||||||||
| 紫苑Tatarian Aster Root | 2.19 | ||||||||||
以麻栎3种群落中重要值大于1的物种为基础, 分别计算各个群落乔木层、灌木层、草本层多样性, 不同林分类型物种多样性存在差异(表 3)。
| 层 Layer |
物种多样性指数 Species diversity index |
PFQ | MFQB | MFQP |
| 乔木层Tree | Simpson指数D | 0.231c | 0.503b | 0.615a |
| Shannon-wiener指数H′ | 0.528c | 1.062b | 1.325a | |
| Pielou指数J′H | 0.347c | 0.580b | 0.664a | |
| Alatalo指数EA | 0.449c | 0.543b | 0.619a | |
| 丰富度Richness R | 4.267c | 5.813b | 5.933a | |
| 灌木层Shrub | Simpson指数D | 0.765a | 0.776a | 0.697b |
| Shannon-wiener指数H′ | 1.702a | 1.883a | 1.774a | |
| Pielou指数J′H | 0.741b | 0.859a | 0.785ab | |
| Alatalo指数EA | 0.770a | 0.807a | 0.762a | |
| 丰富度Richness R | 9.067a | 8.437a | 9.467a | |
| 草本层Herb | Simpson指数D | 0.110b | 0.239a | 0.108b |
| Shannon-wiener指数H′ | 0.270b | 0.426a | 0.425a | |
| Pielou指数J′H | 0.171b | 0.395a | 0.198b | |
| Alatalo指数EA | 0.388b | 0.503a | 0.424ab | |
| 丰富度Richness R | 4.867b | 4.6252b | 5.467a | |
| T-D:乔木Simpson指数, simpson index of tree; T- H′:乔木Shannon-wiener指数, Shannon-wiener index of tree; T-J′H:乔木Pielou指数, Pielou index of tree; T-EA:乔木Alatalo指数, Alatalo index of tree; T-R:乔木丰富度, Richness index of tree; S-D:灌木Simpson指数, simpson index of tree; S- H′:灌木Shannon-wiener指数, Shannon-wiener index of tree; S-J′H:灌木Pielou指数, Pielou index of tree; S-EA:灌木Alatalo指数, Alatalo index of tree; S-R:灌木丰富度, Richness index of tree; H-D:草本Simpson指数, simpson index of tree; H- H′:草本Shannon-wiener指数, Shannon-wiener index of tree; H-J′H:草本Pielou指数, Pielou index of tree; H-EA:草本Alatalo指数, Alatalo index of tree; H-R:草本丰富度, Richness index of tree | ||||
麻栎林3种林分类型乔木层多样性指数(Simpson指数、Shannon-wiener指数)、均匀度指数(Pielou指数、Alatalo指数)均表现为MFQP>MFQB>PFQ (P < 0.05)。3种林分类型灌木层多样性指数与乔木层略有不同, MFQB Simpson指数、Pielou指数、Shannon-wiener指数、Alatalo指数均最高;丰富度指数则呈现出MFQP>PFQ>MFQB的规律。3种类型林分草本层Simpson指数表现为MFQB>PFQ>MFQP;Shannon-wiener指数、Pielou指数及Alatalo指数则呈现出MFQB>MFQP>PFQ的规律;MFQP物种丰富度最高, 分别是PFQ、MFQB的1.12倍、1.18倍。
3.4 三种林分群落结构特征与物种多样性相关关系分析鉴于3种麻栎群落结构特征与物种多样性复杂性, 本文对3种类型麻栎群落结构特征与物种多样性关系进行冗余分析, 试图通过解释变量(群落结构特征)与响应变量(物种多样性)的相关性, 阐明出群落结构特征对物种多样性的影响程度。分析结果见表 4。
| 群落结构特征 Community characteristics |
PFQ | MFQB | MFQP | ||||||
| RDA1 | RDA2 | RDA1 | RDA2 | RDA1 | RDA2 | ||||
| 乔木层Tree | TDBH | 0.408 | -0.245 | -0.210 | -0.345 | 0.392 | -0.077 | ||
| TH | 0.230 | -0.382 | -0.087 | -0.333 | 0.047 | 0.080 | |||
| TCD | -0.180 | 0.410 | -0.152 | 0.077 | 0.017 | 0.138 | |||
| TCBH | -0.193 | 0.636 | -0.405 | -0.341 | 0.153 | -0.480 | |||
| TNTL | -0.274 | 0.498 | -0.193 | 0.028 | 0.335 | 0.244 | |||
| 灌木层Shrub | SH | 0.098 | -0.622 | 0.311 | -0.105 | 0.352 | -0.599 | ||
| SGD | 0.271 | -0.581 | 0.189 | -0.123 | -0.115 | -0.156 | |||
| SCD | 0.151 | -0.713 | 0.297 | 0.059 | -0.047 | -0.285 | |||
| SC | 0.357 | -0.404 | -0.439 | 0.079 | 0.067 | -0.006 | |||
| 草本层Herb | HH | 0.203 | -0.263 | 0.079 | 0.201 | -0.491 | 0.527 | ||
| HD | -0.046 | -0.080 | 0.051 | -0.220 | -0.099 | 0.343 | |||
| HCD | 0.220 | -0.243 | 0.025 | -0.255 | 0.244 | 0.062 | |||
| HC | 0.238 | -0.083 | 0.086 | -0.019 | -0.126 | -0.344 | |||
| 特征参数Characteristic parameter | |||||||||
| 特征值Eigenvalues | 7.8575 | 1.8505 | 2.1451 | 1.5516 | 10.5900 | 1.8023 | |||
| 累计贡献率Explained variation | 0.6749 | 0.8338 | 0.3444 | 0.5935 | 0.7463 | 0.8734 | |||
| 多样性-林分特征相关系数Correlation | 7.8575 | 1.8505 | 2.1451 | 1.5516 | 10.5900 | 1.8023 | |||
| 多样性-林分特征累积贡献率Explained fitted variation | 0.7061 | 0.8724 | 0.4376 | 0.7541 | 0.7937 | 0.9287 | |||
| RDA:冗余分析, Redundancy analysis | |||||||||
PFQ群落结构特征对物种多样性的影响分析, 可以看出物种多样性特征在RDA2轴上具有明显分异, 而在RDA1轴上分异不明显。由图 1可知, 随TDBH、SC及SGD(RDA 1轴)的增加, T-R、S-R及H-R均减小;而当TCBH、TNTL及TCD(RDA 2轴)增加时, S-R及H-R增加, T-R减小。

|
| 图 1 PFQ群落结构特征对多样性影响的RDA二维排序图 Fig. 1 The two-dimensional RDA ordination diagram of effect of community characteristics on species diversity in pure forest of Quercus acutissima PFQ:麻栎纯林, Pure forest of Q. acutissima |
通过MFQB群落结构特征对物种多样性的影响分析, 可以看出物种多样性特征在RDA1轴上具有明显分异, 而在RDA2轴上分异不明显。由图 2可知, 随SH、SCD及SGD(RDA 1轴)的增加, T-R增加, S-R及S-H减小;当HH、SC、TCD增加时, T-R、S-R、H-R及S-H均增加。

|
| 图 2 MFQB群落结构特征对多样性影响的RDA二维排序图 Fig. 2 The two-dimensional RDA ordination diagram of effect of community characteristics on species diversity in mixed forest of Q. acutissima with various broad-leaved trees MFQB:麻栎阔叶混交林, Mixed forest of Q. Acutissima with broad leaved tree |
MFQP群落结构特征对物种多样性的影响分析, 可以看出物种多样性特征在RDA1、RDA2轴上均存在分异现象。由图 3可知, 随TDBH、SH、TNTL(RDA 1轴)的增加, T-R及H-R增加, S-R物种丰富度均减小;而当HH、HD、TNTL(RDA 2轴)增加时, S-R增加, T-R及H-R减小。

|
| 图 3 MFQP群落结构特征对多样性影响的RDA二维排序图 Fig. 3 The two-dimensional RDA ordination diagram of effect of community characteristics on species diversity in mixed forest of Q. acutissima with Pinus tabuliformis MFQP:麻栎油松混交林, Mixed forest of Q. acutissima with Pinus tabulaeformis |
本研究中, 3种类型麻栎群落结构特征存在明显差异。麻栎油松混交林乔木层树高、胸径、冠幅等最高, 栎类混交林以灌木层株高、冠幅、地径等最高, 栎类纯林则以草本层基径、冠幅、盖度等最高。群落结构特征受林分类型及混交程度的影响, 麻栎油松混交林、麻栎阔叶混交林是3种林分中最佳组合, 适合于在生态防护功能提升的基础上生产更多林产品和木材类型。3种类型群落封禁前, 均为阴坡的残败次生林, 1985年前后抚育间伐及砍灌强度基本一致, 最终演替成麻栎纯林、麻栎阔叶混交林和麻栎油松混交林, 可能与当年残败次生林基础有关。
此前关于纯林与混交林群落结构特征分析比较, 多集中于乔木层。从本文分析中也可以看到, 森林结构特征最重要的是乔木层树种组成, 不仅是群落类型划分的依据, 也决定了灌草层植被结构和生长特征。Montagnini等认为混交林乔木层树高、胸径、冠幅等均较纯林高[15]。杨恩让等认为, 混交林较高的胸径、树高及材积与其叶量、叶面积及单位重量的叶绿素含量高有关[16]。群落结构特征这些观点也适合解释3种类型麻栎群落的差异。
4.2 不同类型麻栎群落物种重要值的差异及分析3种麻栎群落类型中, 乔木层重要值最高的物种均为麻栎, 是绝对优势树种。除此之外, 麻栎阔叶混交林中的辽东栎、槲栎重要值也较高;麻栎油松混交林中, 油松重要值仅次于麻栎。3种森林类型中, 灌木层重要值较高的物种均为狼牙刺、南蛇藤、胡枝子;草本层重要值最高的物种为苔草, 说明在同一生境条件下, 林下重要值较高的物种种类受林分类型影响不大, 但不同林分类型中, 林下各物种重要值有差异。群落中, 物种组成及其个体数量是物种之间相互作用结果[17], 也可能是受外界干扰影响[18-20]。本研究3种类型麻栎群落, 封禁前和恢复过程中生境条件基本一致, 之所以出现物种组成及其重要值差异, 原因可能是封禁前林地基础(如在封禁前就有发育成3种林分类型的幼苗幼树)或者1985年抚育间伐方式有差异。
4.3 不同类型麻栎群落物种多样性差异及分析不同林分类型物种多样性存在差异[21]。总体而言, 麻栎油松混交林乔木层、灌木层及草本层物种多样性最高, 其次为麻栎落叶阔叶混交林[22-24]。主要原因可能是, 混交林中, 乔木层植物种类多, 生态位差异大, 对资源利用更充分。特别是麻栎和油松生态位比其他两种类型林分中差异更大, 再加上油松树冠相对其他阔叶树更窄, 更有利于光照进入林地, 导致物种多样性最高。麻栎纯林物种多样性之所以低, 不仅由于乔木层树种单一, 而且单一的乔木层形成的单一胁迫, 导致了灌草层的物种及其个体数量的减少, 进而影响到林下枯落物的分解和微生物的活动, 使麻栎纯林内的生境条件远低于混交林。
4.4 不同类型麻栎群落结构特征与物种多样性相关关系不同群落结构特征对物种多样性的影响不同[25-27]。松栎混交林中, 对物种多样性影响最大的为乔木胸径、新稍长及灌木高度;麻栎阔叶混交林对物种多样性影响最大的为灌木层高度及冠幅;麻栎纯林中, 对物种多样性影响最大的为乔木胸径。目前, 关于不同类型林分群落结构特征对物种多样性影响的研究尚无定论[28]。张建宇等[29]研究认为, 在落叶松群落中, 对物种多样性影响最大的群落结构特征为灌木冠幅、草本层盖度和多度。汪媛燕和王磊等人研究认为, 盖度、多度影响物种丰富度[30-31]。张佳等[32]研究则表明灌木冠幅影响草本多样性。因此, 关于群落结构特征对物种多样性影响的机理还需进一步研究。
5 建议在相似的生境和外界干扰条件下, 经过多年恢复, 麻栎油松混交林、麻栎阔叶混交林的胸径、树高、物种多样性较高, 具有较高木材生产能力和生态防护功能, 是未来森林经营培育的方向。未来森林经营中要充分利用这一规律, 通过人为抚育间伐, 促进麻栎纯林向混交林转换, 促进麻栎落叶阔叶混交林向麻栎油松针阔混交林转化。
不同类型麻栎林的抚育间伐要顺应森林群落演替规律, 采用近自然经营理念, 以复层、混交、异龄为目标林相, 通过定期抚育, 伐除干扰树、病虫木、劣质木, 提高林分质量;对目标树进行合理修枝, 提高枝下高, 改善林地光照, 增加灌草层物种多样性, 促进林地幼苗幼树发育, 促进林地多功能发挥。
| [1] |
欧阳明, 杨清培, 陈昕, 杨光耀, 施建敏, 方向民. 毛竹扩张对次生常绿阔叶林物种组成、结构与多样性的影响. 生物多样性, 2016, 24(6): 649-657. |
| [2] |
苏宇乔, 薛跃规, 范蓓蓓, 莫佛艳, 冯慧喆. 广西流星天坑植物群落结构与多样性. 西北植物学报, 2016, 36(11): 2300-2306. DOI:10.7606/j.issn.1000-4025.2016.11.2300 |
| [3] |
丁晖, 方炎明, 杨新虎, 袁发银, 何立恒, 姚剑飞, 吴俊, 迟斌, 李垚, 陈水飞, 陈婷婷, 徐海根. 黄山亚热带常绿阔叶林的群落特征. 生物多样性, 2016, 24(8): 875-887. |
| [4] |
李皓宇, 彭逸生, 刘嘉健, 王树功, 陈桂珠. 粤东沿海红树林物种组成与群落特征. 生态学报, 2016, 36(1): 252-260. |
| [5] |
刘灿然, 马克平. 北京东灵山地区植物群落多样性的研究. 生态学报, 1997, 17(6): 584-592. DOI:10.3321/j.issn:1000-0933.1997.06.001 |
| [6] |
Lindenmayer D B, Franklin J F, Fischer J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biological Conservation, 2006, 131(3): 433-445. DOI:10.1016/j.biocon.2006.02.019 |
| [7] |
Bordelon M A, McAllister D C, Holloway R. Sustainable forestry:Oregon style. Journal of Forestry, 2000, 98(1): 26-34. |
| [8] |
吕江鱼. 陕西省森林分区抚育经营初探. 陕西林业科技, 2016(2): 32-35. DOI:10.3969/j.issn.1001-2117.2016.02.008 |
| [9] |
薛文艳, 杨斌, 张文辉, 于世川. 桥山林区麻栎种群不同发育阶段空间格局及关联性. 生态学报, 2017, 37(10): 3375-3384. |
| [10] |
黄财智, 张文辉, 李罡, 于世川, 尤健健. 间伐强度对黄桥林区辽东栎林结实规律的影响. 应用生态学报, 2016, 27(11): 3413-3419. |
| [11] |
马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268-277. DOI:10.3321/j.issn:1000-0933.1995.03.006 |
| [12] |
Dixon P. VEGAN, a package of R functions for community ecology. Journal of Vegetation Science, 2010, 14(6): 927-930. |
| [13] |
赖江山, 米湘成.基于Vegan软件包的生态学数据排序分析//第九届全国生物多样性保护与持续利用研讨会论文集.厦门: 国际生物多样性计划中国委员会, 中国科学院生命科学与生物技术局, 2010.
|
| [14] |
Jan L, Petr S. Multivariate analysis of ecological data using CANOCO. Cambridge University Press, 2003, 193-193. |
| [15] |
Montagnini F, González E, Porras C, Rheingans R. Mixed and pure forest plantations in the humid neotropics:a comparison of early growth, pest damage and establishment costs. The Commonwealth Forestry Review, 1995, 74(4): 306-314. |
| [16] |
杨恩让, 韩昭侠. 秦岭华山松红桦混交林与纯林生长调查. 陕西林业科技, 2013(5): 10-12, 38-38. DOI:10.3969/j.issn.1001-2117.2013.05.003 |
| [17] |
毛志宏, 朱教君. 干扰对植物群落物种组成及多样性的影响. 生态学报, 2006, 26(8): 2695-2701. DOI:10.3321/j.issn:1000-0933.2006.08.036 |
| [18] |
Buckley D S, Crow T R, Nauertz E A, Schulz K E. Influence of skid trails and haul roads on understory plant richness and composition in managed forest landscapes in Upper Michigan, USA. Forest Ecology and Management, 2003, 175(1/3): 509-520. |
| [19] |
边巴多吉, 郭泉水, 次柏, 罗大庆, 等. 西藏冷杉原始林林隙对草本植物和灌木树种多样性的影响. 应用生态学报, 2004, 15(2): 191-194. DOI:10.3321/j.issn:1001-9332.2004.02.004 |
| [20] |
Sagar R, Raghubanshi A S, Singh J S. Tree species composition, dispersion and diversity along a disturbance gradient in a dry tropical forest region of India. Forest Ecology and Management, 2003, 186(1/3): 61-71. |
| [21] |
de Grandpre L, Bergeron Y. Diversity and stability of understorey communities following disturbance in the southern boreal forest. Journal of Ecology, 1997, 85(6): 777-784. DOI:10.2307/2960601 |
| [22] |
Gurarni D, Arya N, Yadava A, Ram J. Studies on plant biodiversity of pure Pinus roxburghii sarg. forest and mixed pine-oak forest in uttarakhand Himalaya. New York Science Journal, 2010, 3(8): 1-5. |
| [23] |
White P S. Pattern, process, and natural disturbance in vegetation. The Botanical Review, 1979, 45(3): 229-299. DOI:10.1007/BF02860857 |
| [24] |
Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199(4335): 1302-1310. DOI:10.1126/science.199.4335.1302 |
| [25] |
Collins S L, Glenn S M, Gibson D J. Experimental analysis of intermediate disturbance and initial floristic composition:decoupling cause and effect. Ecology, 1995, 76(2): 486-492. DOI:10.2307/1941207 |
| [26] |
Mackey R L, Currie D J. The diversity-disturbance relationship:is it generally strong and peaked?. Ecology, 2001, 82(12): 3479-3492. |
| [27] |
Cannon C H, Peart D R, Leighton M. Tree species diversity in commercially logged bornean rainforest. Science, 1998, 281(5381): 1366-1368. DOI:10.1126/science.281.5381.1366 |
| [28] |
Thomas S C, Halpern C B, Falk D A, Liguori D A, Austin K A. Plant diversity in managed forests:understory responses to thinning and fertilization. Ecological Applications, 1999, 9(3): 864-879. DOI:10.1890/1051-0761(1999)009[0864:PDIMFU]2.0.CO;2 |
| [29] |
张建宇, 王文杰, 杜红居, 仲召亮, 肖路, 周伟, 张波, 王洪元. 大兴安岭呼中地区3种林分的群落特征、物种多样性差异及其耦合关系. 生态学报, 2018, 38(13): 4684-4693. |
| [30] |
汪媛燕, 王立, 满多清. 民勤绿洲荒漠过渡带群落特征及其物种多样性研究. 四川农业大学学报, 2014, 32(4): 355-361. DOI:10.3969/j.issn.1000-2650.2014.04.001 |
| [31] |
王磊, 高贤明, 孙书存. 岷江上游人工油松林群落空间结构:物种丰富度和盖度. 林业科学, 2004, 40(6): 8-12. DOI:10.3321/j.issn:1001-7488.2004.06.002 |
| [32] |
张佳, 李生宇, 靳正忠, 雷加强. 防护林下草本植物层片物种多样性与环境因子的关系. 干旱区研究, 2011, 28(1): 118-125. |