 2019, Vol. 39
2019, Vol. 39文章信息
- 于成龙, 刘丹
- YU Chenglong, LIU Dan
- 基于MODIS的东北地区自然植被生产力对干旱的响应
- Response in productivity of natural vegetation to drought in northeast China based on MODIS
- 生态学报. 2019, 39(11): 3978-3990
- Acta Ecologica Sinica. 2019, 39(11): 3978-3990
- http://dx.doi.org/10.5846/stxb201803300655
-
文章历史
- 收稿日期: 2018-03-30
- 网络出版日期: 2019-03-21





2. 黑龙江省气象院士工作站, 哈尔滨 150030;
3. 黑龙江省气象科学研究所, 哈尔滨 150030
2. Meteorological Academician Workstation of Heilongjiang Province, Harbin 150030, China;
3. Heilongjiang Province Institute of Meteorological Sciences, Harbin 150030, China
干旱已成为全球陆地生态系统面临的普遍问题[1], 对全球碳、水循环产生重大影响, 进而影响人类的生产和生活[2-3]。东北地区干湿分异典型、植被分布独特、自然地理区域相对完整, 已成为全球变化和生态地理过程研究的热点地区, 该区域生长季干旱的空间分布特征为西重东轻[4], 在近50年中半干旱边界总体呈向东和向南扩展的趋势[5], 而且在未来30年干旱还有可能逐步严重[6-7]。
遥感和地理信息技术的发展提高了区域或全球尺度干旱事件的监测效率, 一些表征干旱的指数也应运而生, 如TVDI(温度植被干旱指数, Temperature-Vegetation Dryness Index)、VCI(植被条件指数, Vegetation Condition index)[8]、TCI(温度条件指数, Temperature Condition Index)[9]、VHI(植被健康指数, Vegetation Health Indices)[10]、DSI(干旱敏感性指数, Drought Severity Index)[11]、SPI(标准化降水指数, Standardized Precipitation Index)[12]、PDSI(帕尔默干旱指数, Palmer Drought Severity Index)[13]和后来改进的scPDSI(改进帕默尔干旱指数, Self-calibrating Palmer Drought Severity Index)[13]等, 在不同尺度和地域监测干旱均取得了较好效果。目前有许多证据表明, 陆地生态系统在受到干旱胁迫时会表现出植物光合作用下降、呼吸代谢降低等适应性状[14-16], 但植物的水分利用效率(Water Use Efficiency, WUE)[11]、碳利用效率(Carbon Use Efficiency, CUE)[17]、土壤微生物活动[18-19]的表现却不尽相同, 而且区域间的植被类型和环境因素差异较大, 结论也会出现差异[20]。
过去的10年是自19世纪80年代以来最温暖的10年[21], 为回答在近几十年中最暖10年间东北地区自然植被对干旱如何响应, 本研究以东北地区自然植被为研究对象, 基于MODIS数据产品、降水数据和scPDSI开展以下研究:①了解2002—2013年东北地区年降水量和干旱的时空分布特征;②分析自然植被NPP、LAI和CUE的时空变化;③明确自然植被LAI、NPP和CUE在持续干旱事件中的变化特征;④探寻独立干旱事件对自然植被LAI、NPP和CUE影响的时间效应。研究结果阐明了东北地区自然植被生态系统在干旱条件下的适应能力, 为了解气候变化背景下自然植被碳水利用提供基础, 进而为区域自然植被可持续发展政策的制定提供参考。
1 研究区概况东北地区包括黑龙江省、吉林省、辽宁省和内蒙古自治区东部的赤峰市、通辽市、呼伦贝尔市及兴安盟[22], 位于111.15°—135.09°E, 37.95°—53.56°N之间, 土地面积78.7万km2, 境内有大兴安岭、小兴安岭、长白山系和东北平原(包括松嫩平原、辽河平原和三江平原), 海拔高度在2—2667 m之间。该地区属温带季风气候, 冬季寒冷干燥, 夏季高温多雨, 近30年(1981—2010年)年平均气温-4.08—11.34℃, 年降水量199.53—1170.60 mm, 日平均日照时数5.26—9.21 h。该区植被属欧亚森林、草原植物亚区和中国-日本森林植物亚区[23]。
2 数据来源与处理 2.1 地理信息数据数字高程模型(DEM)为SRTM地形产品V4.1版本数据, 空间分辨率为90 m, 数据来源于中国科学院计算机网络信息中心国际科学数据镜像网站(http://www.gscloud.cn)。
研究所需的地区级和省级行政区划数据来自于中国气象局下发的1:25万基础地理信息, 对数据进行拓补检查。
2.2 遥感数据除MOD16A2数据产品(数据来源于美国蒙大拿州立大学Numerical Terradynamic Simulation Group, NTSG)外, 本研究所用遥感数据均来源于美国NASA LPDAAC(The Land Processes Distributed Active Archive Center)EOS数据中心。全部遥感数据均应用MODIS Reprojection Tool software(MRT)V4.0进行影像的拼接、重采样(统一空间分辨率为500 m)和投影转换(由Sinusoidal投影转换为经纬度投影)等预处理, 时间范围为2002—2013年, 空间范围为东北地区。
2.2.1 MCD12Q1数据产品本研究基于MODIS的MCD12Q1数据产品(土地类型覆盖产品)的“IGBP全球植被分类方案”的分类结果提取研究区自然植被分布范围, 该数据原始空间分辨率为500 m, 由于该数据产品在相似地类的区分中仍然存在困难[24], 因此根据研究需要, 将MCD12Q1数据分类结果合并为森林、灌木林地、草地、湿地和非自然植被(包括城市用地、农田、水体和雪地), 又因植被类型的改变会在很大程度上改变植被对气候变化的响应程度, 因此本研究只保留了2002—2013年间植被类型没有变化的自然生态系统作为研究对象, 研究对象分布如图 1。由于研究区灌木林和湿地所占面积比例过小(≤0.01%), 所以只研究森林和草地对干旱的响应。

|
| 图 1 东北地区自然植被分布示意图 Fig. 1 Natural vegetation distribution in Northeast China 图中白色部分为非植被区域和2000—2013年土地覆盖类型有变化的区域, 面积由大到小顺序排列为草地(占东北地区总面积的20.45%)、森林(18.73%)、灌木林(0.01%)和湿地(<0.01%) |
叶面积指数(Leaf Area Index, LAI)是植被长势的重要表征指标[25], 本研究选用MOD15A2H数据产品(MODIS/Terra Leaf Area Index/FPAR)分析植被长势对气候变化的响应, 该数据为8 d合成产品, 原始空间分辨率为500 m, LAI为无量纲指数, 数据合并了部分光合有效辐射(Fraction of Photosynthetically Active Radiation, 简称FPAR)和LAI。
2.2.3 MOD17A2H数据产品植被总初级生产力(Gross Primary Productivity, GPP)是单位时间内植物通过光合作用途径所固定的有机碳量, 是地表碳循环的重要组成部分[26], 本研究选用MOD17A2H数据产品(MODIS/Terra Gross Primary Productivity)用于计算植物碳利用效率(CUE), 该数据为8 d合成产品, 原始空间分辨率为500 m, 单位为kgC/m2。
2.2.4 MOD17A3H数据产品植被净初级生产力(Net Primary Productivity, NPP)是陆地生态系统质量的表征指标[27], 本研究选用MOD17A3H数据产品(MODIS/Terra Net Primary Production)分析生态系统碳分配变化的时空特征, 该数据为年合成产品, 原始空间分辨率为500 m, 单位为kg·C/m2。该数据已经通过设立在不同气候区和生物带的涡度相关通量塔数据的检验[28-30], 因此本研究不再对MOD17A3H数据产品的准确性进行验证。
2.3 气象数据 2.3.1 scPDSI数据为分析2002—2013年研究区干湿变化及其对生态系统的影响, 本研究选用全球月scPDSI(Self-calibrating Palmer Drought Severity Index)数据产品, 该数据来源于英国东英格利亚大学(University of East Anglia, UEA)气候研究中心(Climatic Research Unit)编辑的降水和干旱数据集(http://www.cru.uea.ac.uk/), 数据的空间分辨率为50 km, 无量纲, 数据范围为[-5.0, 5.0], 根据研究需要把该数据空间分辨率重采样为500 m。另外, 本研究参考王兆礼等[31]的scPDSI干湿等级划分标准, 根据研究需要把干湿等级进行合并, 得到如下划分方式:scPDSI划分为干旱(scPDSI<-1)正常(-1.0≤scPDSI<1.0)、湿润(scPDSI≥1.0)。
2.3.2 降水数据黑龙江省、吉林省和辽宁省的降水数据来源于各自省份气象局的2002—2013年的共194个气象台站逐日整编资料;内蒙古自治区的降水数据来源于中国气象数据网(http://data.cma.cn/)的“中国地面国际交换站气候资料日值数据集”, 该数据集由各省上报的全国地面月报信息化文件根据《全国地面气候资料(1961—1990)统计方法》及《地面气象观测规范》有关规定整编统计而得, 可下载的内蒙古自治区内的气象观测站有14个。本研究在ARCGIS 10.0软件下利用IDW方法对研究区降水数据进行插值处理, 得到空间分辨率为500 m的逐年月降水量合计值。
2.4 碳利用效率碳利用效率(Carbon Use Efficiency, CUE)反映了生态系统中将大气中CO2转化为生物量的能力, 无量纲, 表达式为:
表达式为:CUE=NPP/GPP
式中, NPP:净初级生产力, GPP:总初级生产力(Gross Primary Productivity)。
3 结果与分析 3.1 东北地区年降水量分布特征东北地区年降水量在120—1132 mm之间, 分布上大体表现为东部高于西部, 其中东南部在1000—1132 mm之间, 中东部在600—1100 mm之间, 西部地区年降水量在600 mm以下(图 2)。降水量分布的像元密度图能够反映一个地区不同降水量所占的面积分布情况, 东北地区年降水量主要集中在200—600 mm之间, 所在区域面积占东北地区总面积的87.00%, 其中年降水量在420—450 mm对应的像元密度最高, 所在区域面积占13.11%;分析森林所在地像元密度分布曲线可见, 森林主要分布在降水量为288—1130 mm之间的区域, 曲线呈现2个峰值, 对应的降水量分别为425—465 mm和536—594 mm之间;草地主要分布在降水量为120—448 mm之间的区域, 像元密度分布曲线也呈现2个峰值, 对应的降水量分别为240—257 mm之间和323—354 mm之间(图 2)。

|
| 图 2 2002—2013年东北地区年降水量分布图 Fig. 2 Scattergram of annual precipitation in Northeast China from 2002 to 2013 |
2002—2013年间, 研究区每年都有发生干旱的区域(图 3), 其中2002年、2007年和2008年发生干旱的面积较大, 分别占研究区总面积的81.73%、77.86%和84.85%。

|
| 图 3 2002—2013年东北地区干旱面积和频次分布 Fig. 3 Distribution of drought area and frequency in Northeast China from 2002 to 2013 |
分析干旱发生频次的空间分布可知(图 3), 内蒙古草原和松嫩平原大部地区发生干旱的频次较高, 在6—11次之间;大兴安岭和长白山北部发生干旱的频次较低, 为0—2次;其他区域发生干旱的频次在3—5次之间。
3.3 自然植被LAI、NPP和CUE的时空变化分析图 4可见, 森林的NPP年平均值明显高于草地, 但森林或草地各自的NPP年平均值的年际差异较小, 森林年平均NPP在412.54—491.76之间, 草地年平均NPP在169.15—259.07之间。从空间分布看, 东北地区自然植被NPP年平均值出现较明显的三级分布, 其中长白山脉和小兴安岭南部地区的NPP较大, 在550.00—1799.97之间;大兴安岭和小兴安岭大部分地区的NPP居中, 在300.00—600.00之间;中西部草原区域的NPP最小, 在100.00—350.00之间。

|
| 图 4 东北地区自然植被LAI、NPP和CUE时空变化 Fig. 4 Spatial distribution and interannual Changes of LAI, NPP and CUE of the nature vegetation in Northeast China |
分析图 4可见, 与NPP的年际分布规律相似, 森林的LAI年平均值明显高于草地, 但森林或草地的LAI年平均值的年际差异较小, 森林年平均LAI在1.61—1.80之间, 草地年平均LAI在0.31—0.42之间。从LAI的空间分布看, 东北地区自然植被LAI年平均值的空间分布差异较大(范围在0.01—3.04之间), 表现出明显的植被分异特征:大兴安岭、小兴安岭和长白山脉植被的LAI较高(该区域主要植被类型为森林), 其中海拔低于800 m的植被LAI在2.50—3.04之间, 海拔高于800 m的植被的LAI在1.48—2.60之间;东北地区中西部地区植被的LAI较低(该区域主要的植被类型为草地), 其中中部地区植被的LAI在0.34—1.49之间, 西部地区植被的LAI在0.01—0.36之间。
分析图 4可见, 草地的CUE年平均值明显高于森林, 但森林和草地各自的CUE年平均值的年际差异较小, 森林年平均CUE在0.50—0.58之间, 草地年平均CUE在0.61—0.64之间。从空间分布看, 东北地区自然植被CUE年平均值也存明显的空间分异, 但与LAI分布不同, 西部草原区的CUE最大, 在0.70—1.10之间;中部草原区的CUE居中, 在0.50—0.80之间, 大兴安岭、小兴安岭和长白山脉绝大多数植被的CUE小于0.60。
3.4 自然植被对独立干旱事件的响应为研究干旱事件对森林和草地的后续影响, 本研究仍然以0.2°×0.2°为间隔提取2002—2013年各像元的NPP、LAI和CUE, 对参与分析的像元进行空间定位, 对比分析干旱年与干旱前的水分正常年森林和草地的NPP、LAI和CUE的关系, 以及干旱年后连续4年水分正常年与干旱前的水分正常年的森林和草地NPP、LAI和CUE的关系。
3.4.1 森林对独立干旱事件的响应应用独立样本T检验方法, 借助SPSS 19.0, 分别检验干旱年、干旱年后连续4年与干旱前的水分正常年森林的NPP、LAI和CUE平均值之间的差异, 结果如表 1, 为表述简洁, 把干旱年森林NPP平均值记为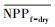
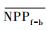

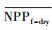
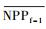
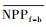
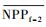
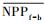
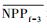
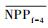
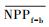
| 指数 Index |
干旱年 Drought year |
干旱后第1个水分正常年 First normal year of moisture after drought |
干旱后第2个水分正常年 Second normal year of moisture after drought |
干旱后第3个水分正常年 The third normal year of moisture after drought |
干旱后第4个水分正常年 The fourth normal year of moisture after drought |
| ΔNPP | -20.34 | -9.70 | 2.02 | -68.47** | -44.47** |
| ΔLAI | -0.01 | 0.03 | 0.13** | 0.07 | 0.10** |
| ΔCUE | 0.00 | 0.00 | -0.02* | -0.04** | -0.02 |
| *为两组对比数据均值存在显著差异, **为存在极显著差异; ΔNPP:植被净初级生产力之差;ΔLAI:叶面积指数之差;ΔCUE:碳利用效率之差; NPP:植被净初级生产力, Net Primary Productivity;LAI:叶面积指数, Leaf Area Index;CUE:碳利用效率, Carbon Use Efficiency | |||||
为进一步探究干旱事件对森林NPP、LAI和CUE的后续影响, 本研究把干旱年前的水分条件正常年森林的NPP记为NPPBaseline, 把干旱年后连续n个水分正常年森林NPP与对应像元的NPPBaseline之差记为ΔNPPn, 分析ΔNPPn和NPPBaseline的关系, LAI和CUE命名方式和研究方法同NPP。
图 5描述了干旱年前后森林NPP、LAI和CUE的变化, 分析可见, 干旱后第1和第2个水分正常年有50%左右的NPP较干旱前高, 并且ΔNPPn和NPPBaseline存在显著的线性负相关关系(sig.<0.001);分析图 5中LAI变化情况可见, 干旱事件后4年ΔLAIn和LAIBaseline存在显著线性负相关关系, 而且ΔLAIn≥0的像元比例在逐年增加;干旱对森林CUE影响的规律性较差, 只有在干旱年发生后第4个水分正常年ΔCUEn和CUEBaseline才出现显著的负相关关系。

|
| 图 5 独立干旱年前后森林NPP、LAI和CUE的变化 Fig. 5 Changes of NPP, Lai and cue in forest before and after drought year NPPBaselin:干旱年前的水分条件正常年的NPP;LAIBaseline:干旱年前的水分条件正常年的LAI;CUEBaseline:干旱年前的水分条件正常年的CUE |
把干旱年草地NPP平均值记为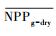

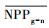

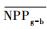
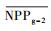
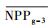
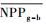
| 指数 Index |
干旱当年 Drought year |
干旱后1年 One year after the drought |
干旱后2年 Two years after the drought |
| ΔNPP | -22.34** | 63.14** | 37.77** |
| ΔLAI | -0.05** | 0.06 | 0.06 |
| ΔCUE | 0 | 0.02 | 0.01 |
应用与图 5相同的方法探究干旱事件发生前后草地NPP、LAI和CUE的变化特征发现, 干旱事件发生后1年和2年的ΔNPPn和NPPBaseline、ΔLAIn和LAIBaseline、ΔCUEn和CUEBaseline均没有显著的相关关系(sig>0.05), 由此进一步印证了干旱事件仅在当年对草地产生显著影响。
3.5 自然植被对持续干旱事件的响应 3.5.1 森林对持续干旱事件的响应为探究森林在发生持续干旱事件时NPP、LAI和CUE的变化规律, 以0.2°×0.2°为间隔提取2002—2013年各像元的NPP、LAI和CUE, 对参与分析的像元进行空间定位, 计算各指数在水分正常年、第1个干旱年、第2个干旱年、……、第5个干旱年中的平均值, 并应用独立样本T检验方法, 借助SPSS 19.0检验干旱年各指数平均值与水分正常年平均值之间的差异, 结果如表 3, 分析可见, 随着干旱的持续, NPP呈现显著线性下降趋势(y =-6.55x+544.80, R2=0.762, sig.=0.023), 而且第3个干旱年和第4个干旱年的NPP均值显著低于水分正常年的NPP均值, 第5个干旱年更是出现了极显著差异;LAI和CUE虽然也存在下降趋势, 但下降趋势并不显著(线性拟合显著性检验sig.分别为0.096和0.796), 另外第4个干旱年和第5干旱年LAI均值显著低于水分正常年LAI均值;干旱年CUE均值与水分正常年没有显著差异。由此可见, 持续干旱从总体上能够显著降低研究区森林的NPP和LAI, 但对CUE的影响并不显著。
| 指数 Index |
水分条件正常 Normal water condition |
第1个干旱年 Drought of the first year |
第2个干旱年 Drought of the second year |
第3个干旱年 Drought of the third year |
第4个干旱年 Drought of the fourth year |
第5个干旱年Drought of the fifth year |
| NPP | 543.38 | 523.04 | 530.55 | 511.46* | 519.11* | 503.70** |
| LAI | 1.71 | 1.70 | 1.76 | 1.66 | 1.60* | 1.63* |
| CUE | 0.59 | 0.59 | 0.56 | 0.58 | 0.59 | 0.58 |
本研究通过画散点图和做线性趋势分析的方法, 从像元水平进一步探究森林NPP、LAI和CUE在持续干旱年与水分正常年之间的关系。其中水分正常年像元对应的NPP、LAI和CUE分别记为NPPBaseline、LAIBaseline和CUEBaseline;某像元第1个干旱年的NPP记为NPPDry1, 第2个记为NPPDry2, ……, 第n个记为NPPDryn;NPPDryn与NPPBaseline之差记为ΔNPPDryn, LAI和CUE的命名方式同NPP。
图 6显示了连续发生干旱5年的森林NPP、LAI和CUE与水分正常年之间的关系, 分析可见, 干旱会使部分森林的NPP有所增加, 而且随着干旱的持续, NPP增加的像元比例有扩大的趋势, 但ΔNPPDryn与NPPBaseline之间并没有显著线性关系;ΔLAIDryn>0的比例没有明显的变化趋势, 但在第1、2、4、5个干旱年ΔLAIDryn与LAIBaseline之间存在显著线性负相关关系(sig<0.001);ΔCUEDryn>0的比例变化较大, 在研究时段内未见明显的变化规律, 而且ΔCUEDry与CUEBaseline之间并没有显著线性关系。由此可见, 研究区部分森林的NPP、LAI和CUE对持续干旱均具有一定的适用能力, 其中LAIBaseline较低的森林, 在持续干旱时ΔLAIDryn增加的可能性越大, 但本研究并未发现NPP和CUE存在相似的变化规律。

|
| 图 6 1—5年持续干旱年森林NPP、LAI和CUE的变化 Fig. 6 Changes of NPP, LAI and CUE in forest during 1—5 years of persistent drought NPPDryn:第n个干旱年NPP与水分正常年NPP之差;LAIDryn:第n个干旱年LAI与水分正常年LAI之差;CUEDryn:第n个干旱年CUE与水分正常年CUE之差 |
分析持续干旱年草地NPP、LAI和CUE平均值的变化可见(表 4), 5个连续干旱年草地NPP平均值显著低于水分正常年NPP的平均值, 但并没有随着连续干旱年的发生而出现明显的变化趋势;干旱年的LAI平均值也低于水分正常年, 而且除第3个干旱年的LAI平均值与水分正常年LAI平均值无显著差异外, 其他干旱年均出现显著差异;对于CUE来说, 每个干旱年CUE的平均值与水分正常年的CUE平均值均未出现显著差异。由此可见, 持续干旱总体上显著降低了草地的NPP和LAI, 但对草地CUE总体上并未产生显著影响。
| 指数 Index |
水分条件正常 Normal water condition |
第1年干旱 Drought of the first year |
第2年干旱 Drought of the second year |
第3年干旱 Drought of the third year |
第4年干旱 Drought of the fourth year |
第5年干旱 Drought of the fifth year |
| NPP | 217.69 | 195.35** | 177.08** | 195.59** | 197.66** | 187.88** |
| LAI | 0.42 | 0.37** | 0.35** | 0.40 | 0.36** | 0.35** |
| CUE | 0.62 | 0.62 | 0.61 | 0.59 | 0.62 | 0.61 |
从像元水平上分析(图 7), 与森林相似, 草地在持续干旱条件下也表现出部分NPP增加的现象, 在连续干旱4年过程中NPP增加的草地像元比例有扩大的趋势, 但到第5年比例出现明显减小, 由于研究数据时长的限制, 未获得更长时间序列持续干旱数据, 因此不能判断这种减小的趋势是否会持续。另外, 除持续第3年干旱时ΔNPPDryn与NPPBaseline没有显著的变化趋势外, 其他干旱的年份中ΔNPPDryn与NPPBaseline均存在显著的线性负相关关系(sig<0.001);在每个干旱年份中均有部分草地的LAI和CUE会增加, 但随着干旱事件的持续, 增加的比例没有明显的线性规律, 另外, 多数年份的ΔLAIDryn与LAIBaseline、ΔCUEDryn与CUEBaseline也存在显著的线性负相关关系(sig<0.001)。由此可见, 研究区部分草地对持续干旱具有一定的适用能力, 而且NPPBaseline、LAIBaseline和CUEBaseline较低的草地, 在持续干旱时ΔNPPDryn、ΔLAIDryn和ΔCUEDryn增加的可能性越大。

|
| 图 7 1—5年持续干旱年草地NPP、LAI和CUE的变化 Fig. 7 Changes of NPP, LAI and CUE in grasslands during 1—5 years of persistent drought |
本研究结果表明, 森林年平均CUE在0.50—0.58之间, 草地年平均CUE在0.61—0.64之间, 与许多研究报道CUE的范围(0.20—0.80之间)相一致[32], 同时草地的CUE年平均值明显高于森林的结论, 也与Choudhury[33]的结论一致。另外, 本研究中发现草地与森林的CUE不同, 即使同为草地, 由于所处地理位置不同, CUE的年平均值也会存在较大差异, 这个结论更偏向于支持CUE并非恒定的观点[34-36]。
在研究中发现, 干旱从总体上降低了东北地区自然植被的NPP、LAI和CUE, 这一结论与多位研究者的结论是一致的[37-39], 但干旱持续时间和植被类型的差异, 使NPP、LAI和CUE的变化也不尽相同。在研究森林对独立干旱年的响应时发现, 森林在干旱年过后的第2个水分正常年LAI平均值较干旱前有显著提高、CUE平均值出现显著降低的现象, NPP平均值在干旱后第3个和第4个水分正常年出现显著降低的现象, 可见独立干旱事件对森林造成的影响至少可以持续到干旱过后第4年, 而独立的干旱事件对草地产生的影响效应在水分条件恢复后随即消逝, 这与Wu等[40]认为干旱对森林生长的遗产效应可以长达4年、草地不超过1年的研究结论是一致的。另外值得注意的是, 森林在经历了独立干旱年后, ΔLAIn和LAIBaseline均存在显著线性负相关关系, 而且ΔLAIn≥0的像元比例在逐年增加, 原因可能是干旱发生时森林通过稀疏的叶面积或矮小的枝干以适应水分匮缺的环境[41-42], 但水分恢复正常后LAI会出现反弹效应, 使LAI比干旱前更高, 这一现象在其他地区的研究中未见报道。
同时研究发现, 森林在第3个持续干旱年的NPP平均值出现显著下降的现象, 在第4个持续干旱年LAI平均值出现显著下降的现象, 草地的NPP平均值和LAI平均值都是在第1个干旱年便出现显著下降的情况, 这种反应时间的差别可能与森林和草地根系深度有关, 森林主要利用深层土壤的水分, 而基于scPDSI划分的标准反映的是浅层土壤的旱涝程度, 初期浅层土壤的水分匮缺可能对深层土壤水分不会产生过多影响, 只能影响树木的浅层根系对水分的吸收, 因此虽然干旱初期森林的NPP平均值出现下降, 但与水分正常年平均值并没有显著差异, 持续干旱导致深层土壤水分得不到有效补充, 树木深层根系便会出现吸水困难的现象, 由此森林NPP平均值显著低于水分正常年的NPP平均值。而草地的根系较浅, 浅层土壤水分条件的变化便会对其产生显著影响。
本研究的数据基础是MODIS数据产品, 除其固有的空间分辨率的限制外, 由于本研究的时空局限性还在于:缺乏对MODIS数据产品有效的地面验证工作;只区分了森林和草地, 忽略了在研究过程中森林和草地内部种的变化;由于干旱发生的时间不同, 对植被生理过程的影响也存在差异, 但本研究只考虑了年度水分的变化, 没有区分干旱发生的时间。尽管存在以上的限制, 但本研究还是敏感地捕捉到了无论是独立干旱还是持续干旱, 东北地区部分植被的NPP、LAI和CUE在干旱条件下均有增加的现象, 表明从生态系统水平上东北地区森林和草地均具有一定的抗旱性。
5 结论基于2001—2013年降水数据、scPDSI数据和MODIS的土地类型覆盖、LAI、GPP、NPP数据产品, 分析东北地区降水和干旱的发生规律, 研究干旱对东北地区森林和草地的LAI、NPP和CUE对干旱的响应关系, 结果表明:
① 东北地区年降水量在120—1132 mm之间, 大体分布为东高西低, 森林主要分布在降水量为288—1130 mm之间的区域, 草地主要分布在降水量为120—448 mm之间的区域。
② 东北地区自然植被所在区域每年都有干旱发生, 内蒙古草原和松嫩平原大部地区发生干旱的频次较高。
③ 2002—2013年平均水平上看, 森林的NPP年平均值(412.54—491.76之间)和LAI年平均值(1.61—1.80之间)明显高于草地(NPP年平均值在169.15—259.07之间, LAI在0.31—0.42之间), 森林的CUE年平均值(0.50—0.58之间)略低于草地(0.61—0.64之间)。
④ 森林和草地均通过降低NPP和LAI来应对独立干旱事件, 其中森林在干旱年过后第3年NPP会显著下降, LAI在干旱年后第2年比干旱前显著升高, 而且这种变化效应至少持续到干旱年过后第4年;草地仅在独立干旱年当年有显著变化。
⑤ 随着干旱的持续, 森林NPP增加的比例有扩大趋势, LAIBaseline较低的森林在持续干旱时ΔLAIDryn增加的可能性越大;草地对持续干旱也具有一定的适用能力, 而且NPPBaseline、LAIBaseline和CUEBaseline较低的草地, 在持续干旱时ΔNPPDryn、ΔLAIDryn和ΔCUEDryn增加的可能性越大。
⑥ 无论是独立干旱事件还是持续干旱, 森林或草地的CUE变化很少达到显著性水平, 变化规律的显著性也低于NPP和LAI。
| [1] |
Houghton J T, Fihlo L G M, Callander B A, Harris N, Kattenberg A, Maskell K. Climate Change 1995:The Science of Climate Change:Contribution of Working Group I to the Second Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 1996.
|
| [2] |
Van Der Molen M K, Dolman A J, Ciais P, Eglin T, Gobron N, Law B E, Meir P, Peters W, Phillips O L, Reichstein M, Chen T, Dekker S C, Doubková M, Friedl M A, Jung M, Van Den Hurk B J J M, De Jeu R A M, Kruijt B, Ohta T, Rebel K T, Plummer S, Seneviratne S I, Sitch S, Teuling A J, Van Der Werf G R, Wang G. Drought and ecosystem carbon cycling. Agricultural and Forest Meteorology, 2011, 151(7): 765-773. DOI:10.1016/j.agrformet.2011.01.018 |
| [3] |
Jia B H, Wang Y Y, Xie Z H. Responses of the terrestrial carbon cycle to drought over China:Modeling sensitivities of the interactive nitrogen and dynamic vegetation. Ecological Modelling, 2018, 368: 52-68. DOI:10.1016/j.ecolmodel.2017.11.009 |
| [4] |
袭祝香, 杨雪艳, 刘实, 纪玲玲. 东北地区夏季干旱风险评估与区划. 地理科学, 2016, 33(6): 735-740. |
| [5] |
赵东升, 吴绍洪. 气候变化情景下中国自然生态系统脆弱性研究. 地理学报, 2013, 68(5): 602-610. |
| [6] |
卢洪健, 莫兴国, 孟德娟, 刘苏峡. 气候变化背景下东北地区气象干旱的时空演变特征. 地理科学, 2015, 35(8): 1051-1059. |
| [7] |
马建勇, 许吟隆. 东北地区作物生长季干旱时空分布特征及其环流背景. 中国农业气象, 2013, 34(1): 81-87. DOI:10.3969/j.issn.1000-6362.2013.01.012 |
| [8] |
Kogan F N. Remote sensing of weather impacts on vegetation in non-homogeneous areas. International Journal of Remote Sensing, 1990, 11(8): 1405-1419. DOI:10.1080/01431169008955102 |
| [9] |
Kogan F N. Application of vegetation index and brightness temperature for drought detection. Advances in Space Research, 1995, 15(11): 91-100. DOI:10.1016/0273-1177(95)00079-T |
| [10] |
Sholihah R I, Trisasongko B H, Shiddiq D, Iman L O S, Kusdaryanto S, Manijo, Panuju D R. Identification of agricultural drought extent based on vegetation health indices of Landsat data:case of Subang and Karawang, Indonesia. Procedia Environmental Sciences, 2016, 33: 14-20. DOI:10.1016/j.proenv.2016.03.051 |
| [11] |
Gang C C, Wang Z Q, Chen Y Z, Yang Y, Li J L, Cheng J M, Qi J G, Odeh I. Drought-induced dynamics of carbon and water use efficiency of global grasslands from 2000 to 2011. Ecological Indicators, 2016, 67: 788-797. DOI:10.1016/j.ecolind.2016.03.049 |
| [12] |
Zarch M A A, Sivakumar B, Sharma A. Droughts in a warming climate:a global assessment of Standardized precipitation index (SPI) and Reconnaissance drought index (RDI). Journal of Hydrology, 2015, 526: 183-195. DOI:10.1016/j.jhydrol.2014.09.071 |
| [13] |
Ficklin D L, Letsinger S L, Gholizadeh H, Maxwell J T. Incorporation of the Penman-Monteith potential evapotranspiration method into a Palmer Drought Severity Index tool. Computers & Geosciences, 2015, 85: 136-141. |
| [14] |
Metcalfe D B, Meir P, Aragão L E O C, Lobo-do-Vale R, Galbraith D, Fisher R A, Chaves M M, Maroco J P, Da Costa A C L, De Almeida S S, Braga A P, Gonçalves P H L, De Athaydes J, Da Costa M, Portela T T B, De Oliveira A A R, Malhi Y, Williams M. Shifts in plant respiration and carbon use efficiency at a large-scale drought experiment in the eastern Amazon. New Phytologist, 2010, 187(3): 608-621. DOI:10.1111/j.1469-8137.2010.03319.x |
| [15] |
Drake J E, Power S A, Duursma R A, Medlyn B E, Aspinwall M J, Choat B, Creek D, Eamus D, Maier C, Pfautsch S, Smith R A, Tjoelker M G, Tissue D T. Stomatal and non-stomatal limitations of photosynthesis for four tree species under drought:a comparison of model formulations. Agricultural and Forest Meteorology, 2017, 247: 454-466. DOI:10.1016/j.agrformet.2017.08.026 |
| [16] |
Gleason S M, Wiggans D R, Bliss C A, Comas L H, Cooper M, DeJonge K C, Young J S, Zhang H H. Coordinated decline in photosynthesis and hydraulic conductance during drought stress in Zea mays. Flora, 2017, 227: 1-9. DOI:10.1016/j.flora.2016.11.017 |
| [17] |
Manzoni S, Taylor P, Richter A, Porporato A, Agren G I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytologist, 2012, 196(1): 79-91. DOI:10.1111/j.1469-8137.2012.04225.x |
| [18] |
Fuchslueger L, Bahn M, Hasibeder R, Kienzl S, Fritz K, Schmitt M, Watzka M, Richter A. Drought history affects grassland plant and microbial carbon turnover during and after a subsequent drought event. Journal of Ecology, 2016, 104(5): 1453-1465. DOI:10.1111/1365-2745.12593 |
| [19] |
Malone S L, Starr G, Staudhammer C L, Ryan M G. Effects of simulated drought on the carbon balance of Everglades short-hydroperiod marsh. Global Change Biology, 2013, 19(8): 2511-2523. DOI:10.1111/gcb.12211 |
| [20] |
王兆礼, 李军, 黄泽勤, 钟睿达, 陈佳颖, 邱卓辉. 基于改进帕默尔干旱指数的中国气象干旱时空演变分析. 农业工程学报, 2016, 32(2): 161-168. |
| [21] |
Zhao M S, Running S W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science, 2010, 329(5994): 940-943. DOI:10.1126/science.1192666 |
| [22] |
王士君, 宋飏. 中国东北地区城市地理基本框架. 地理学报, 2006, 61(6): 574-584. DOI:10.3321/j.issn:0375-5444.2006.06.002 |
| [23] |
徐文铎. 中国东北主要植被类型的分布与气候的关系. 植物生态学与地植物学学报, 1986, 10(4): 254-263. |
| [24] |
Friedl M A, Sulla-Menashe D, Tan B, Schneider A, Ramankutty N, Sibley A, Huang X M. MODIS Collection 5 global land cover:Algorithm refinements and characterization of new datasets. Remote sensing of Environment, 2010, 114(1): 168-182. |
| [25] |
Iio A, Hikosaka K, Anten N P R, Nakagawa Y, Ito A. Global dependence of field-observed leaf area index in woody species on climate:a systematic review. Global Ecology and Biogeography, 2014, 23(3): 274-285. DOI:10.1111/geb.12133 |
| [26] |
Croft H, Chen J M, Froelich N J, Chen B, Staebler R M. Seasonal controls of canopy chlorophyll content on forest carbon uptake:implications for GPP modeling. Journal of Geophysical Research, 2015, 120(8): 1576-1586. |
| [27] |
Murray-Tortarolo G, Friedlingstein P, Sitch S, Seneviratne S I, Fletcher I, Mueller B, Greve P, Anav A, Liu Y, Ahlström A, Huntingford C, Levis S, Levy P, Lomas M, Poulter B, Viovy N, Zaehle S, Zeng N. The dry season intensity as a key driver of NPP trends. Geophysical Research Letters, 2016, 43(6): 2632-2639. DOI:10.1002/grl.v43.6 |
| [28] |
Turner D P, Ritts W D, Cohen W B, Gower S T, Running S W, Zhao M S, Costa M H, Kirschbaum A A, Ham J M, Saleska S R, Ahl D E. Evaluation of MODIS NPP and GPP products across multiple biomes. Remote Sensing of Environment, 2006, 102(3/4): 282-292. |
| [29] |
Cracknell A P, Kanniah K D, Tan K P, Wang L. Towards the development of a regional version of MOD17 for the determination of gross and net primary productivity of oil palm trees. International Journal of Remote Sensing, 2015, 36(1): 262-289. DOI:10.1080/01431161.2014.995278 |
| [30] |
Malone S L, Tulbure M G, Pérez-Luque A J, Assal T J, Bremer L L, Drucker D P, Hillis V, Varela S, Goulden M L. Drought resistance across California ecosystems:evaluating changes in carbon dynamics using satellite imagery. Ecosphere, 2016, 7(11): e01561. DOI:10.1002/ecs2.1561 |
| [31] |
王兆礼, 黄泽勤, 李军, 钟睿达, 黄文炜. 基于SPEI和NDVI的中国流域尺度气象干旱及植被分布时空演变. 农业工程学报, 2016, 32(14): 177-186. DOI:10.11975/j.issn.1002-6819.2016.14.024 |
| [32] |
DeLucia E H, Drake J E, Thomas R B, Gonzalez-MELER M. Forest carbon use efficiency:is respiration a constant fraction of gross primary production?. Global Change Biology, 2007, 13(6): 1157-1167. DOI:10.1111/gcb.2007.13.issue-6 |
| [33] |
Choudhury B J. Carbon use efficiency, and net primary productivity of terrestrial vegetation. Advances in Space Research, 2000, 26(7): 1105-1108. DOI:10.1016/S0273-1177(99)01126-6 |
| [34] |
Rowland L, Hill T C, Stahl C, Siebicke L, Burban B, Zaragoza-Castel J, Ponton S, Bonal D, Meir P, Williams M. Evidence for strong seasonality in the carbon storage and carbon use efficiency of an Amazonian forest. Global Change Biology, 2014, 20(3): 979-991. DOI:10.1111/gcb.2014.20.issue-3 |
| [35] |
Zhang Y J, Yu G R, Yang J, Wimberly M C, Zhang X Z, Tao J, Jiang Y B, Zhu J T. Climate-driven global changes in carbon use efficiency. Global Ecology and Biogeography, 2014, 23(2): 144-155. DOI:10.1111/geb.2014.23.issue-2 |
| [36] |
Yang Y, Wang Z Q, Li J L, Gang C C, Zhang Y Z, Odeh I, Qi J G. Assessing the spatiotemporal dynamic of global grassland carbon use efficiency in response to climate change from 2000 to 2013. Acta Oecologica, 2017, 81: 22-31. DOI:10.1016/j.actao.2017.04.004 |
| [37] |
Jassal R S, Black T A, Cai T B, Morgenstern K, Li Z, Gaumont-Guay D, Nesic Z. Components of ecosystem respiration and an estimate of net primary productivity of an intermediate-aged Douglas-fir stand. Agricultural and Forest Meteorology, 2007, 144(1/2): 44-57. |
| [38] |
Zhang Y J, Xu M, Chen H, Adams J. Global pattern of NPP to GPP ratio derived from MODIS data:effects of ecosystem type, geographical location and climate. Global Ecology and Biogeography, 2009, 18(3): 280-290. DOI:10.1111/geb.2009.18.issue-3 |
| [39] |
Helman D, Lensky I M, Yakir D, Osem Y. Forests growing under dry conditions have higher hydrological resilience to drought than do more humid forests. Global Change Biology, 2017, 23(7): 2801-2817. DOI:10.1111/gcb.2017.23.issue-7 |
| [40] |
Wu X C, Liu H Y, Li X Y, Ciais P, Babst F, Guo W C, Zhang C C, Magliulo V, Pavelka M, Liu S M, Huang Y M, Wang P, Shi C M, Ma Y J. Differentiating drought legacy effects on vegetation growth over the temperate Northern Hemisphere. Global Change Biology, 2018, 24(1): 504-516. |
| [41] |
刘长成, 刘玉国, 郭柯. 四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性. 植物生态学报, 2011, 35(10): 1070-1082. |
| [42] |
朱万泽. 森林碳利用效率研究进展. 植物生态学报, 2013, 37(11): 1043-1058. |