 2018, Vol. 38
2018, Vol. 38文章信息
- 杨立荣, 张治礼, 云勇, 严武平, 陈宣, 张浪, 郑道君, 陈加利.
- YANG Lirong, ZHANG Zhili, YUN Yong, YAN Wuping, CHEN Xuan, ZHANG Lang, ZHENG Daojun, CHEN Jiali.
- 濒危植物海南龙血树的种群结构与动态
- The population structure and dynamics of Dracaena cambodiana, an endangered tree on Hainan Island
- 生态学报. 2018, 38(8): 2802-2815
- Acta Ecologica Sinica. 2018, 38(8): 2802-2815
- http://dx.doi.org/10.5846/stxb201708071415
-
文章历史
- 收稿日期: 2017-08-07
- 网络出版日期: 2018-01-08





2. 海南省农业科学院农业环境与土壤研究所, 海口 571100;
3. 海南大学, 海口 570228;
4. 海南省热带特种经济植物种质资源创新利用重点实验室, 海口 571100
2. Agricultural Environment And Soil Research Institute in Hainan Academy of Agricultural Sciences, Haikou 571100, China;
3. Hainan University, Haikou 570228, China;
4. The Key Laboratory of Tropic Special Economic Plant Innovation and Utilization, Hainan Province, Haikou 571100, China
种群通常指某一特定时间和区域内同一物种构成的群体, 它们共享同一基因库或存在潜在随机交配能力[1]。种群结构是种群的最基本特征, 一直以来都是种群生态学研究的核心内容之一, 通常通过分析种群的年龄结构、生命表和存活曲线等方法来获得[2-3]。种群结构既能反映现在的种群状态, 即现在种群内不同大小个体数量的分布和配置状况, 也能反映植物与环境之间的适合度, 分析过去种群结构与受干扰情况, 预测未来的种群动态和演变趋势[4-6]。因此, 研究种群的结构及其动态, 有助于人们更好地认识种群的生态特性、稳定性及发展动态等, 对濒危种群的更新复壮及其所在群落和生态系统的恢复具有重要意义。
海南龙血树(Dracaena cambodiana)(小花龙血树、柬埔寨龙血树), 属百合科龙血树属的单子叶植物[7-8], 国家二级重点保护植物和稀有濒危保护植物[9-10]。海南龙血树是一种生长缓慢, 耐干旱的喜阳植物, 目前分布于海南岛西南部的内陆山区及南部沿海地区, 生长于裸露花岗岩和石灰岩的石缝残积土中或紧贴石壁生长于砂壤土中, 属于典型的岩石伴生植物[11-12]。由于海南龙血树具有较高的药用价值和观赏价值[13-15], 野生资源受到掠夺性采挖, 野生资源已十分匮乏, 种群处于濒危状态。目前关于海南龙血树的研究主要集中在组织快繁、栽培技术、以血竭为基础的基础性研究和开发利用以及分子生物学等方面[16-19]。此外, 郑道君等人对海南龙血树野生资源分布、种群更新情况及种子萌发进行了研究[12, 20-21]。近年来, 海南龙血树的种群结构和分布格局也有报道, 但周文嵩等人仅对昌江地区的海南龙血树种群结构进行了研究[22], 李伟杰等人涉及的种群虽多, 但仅对海南龙血树种群的年龄结构和分布格局进行了研究, 并未对海南龙血树种群的静态生命表、数量动态特征和生存状况进行分析[23]。本文拟以处于不同代表性生境的海南龙血树种群为对象, 对它们的年龄结构、数量动态特征与生存状况进行定量研究, 预测种群未来的动态趋势, 以期从种群生态学方面为海南龙血树种群的保护和恢复提供依据。
1 研究方法 1.1 野外调查在查阅文献资料的基础上, 采用踏查法对海南龙血树种群进行实地调查。海南龙血树主要分布在石灰岩和花岗岩两种生境中, 因此, 本文对两种生境的11个种群进行了调查。其中, 石灰岩生境调查了东方市俄贤岭的2个种群; 花岗岩生境调查了分布于海边的三亚南山和亚龙湾种群、万宁乐南山种群、昌江昌化岭种群, 以及分布于非海边的三架岭种群、东方黑岭种群和乐东尖峰岭的3个种群(表 1)。各种群的地理位置如图 1所示。采用路线调查法调查种群内的海南龙血树个体, 记录个体的株型、基径、树高及冠幅。
| 种群 Population |
海拔/m Altitude |
东经 Latitude |
北纬 Longitude |
个体数量/株 Individual |
分布点 Location |
生境类型 Habitat type |
| NS | 15 | 109°09′5.50″ | 18°18′3.08″ | 83 | 三亚市南山 | 花岗岩 |
| YL | 59 | 109°39′16.50″ | 18°14′21.78″ | 19 | 三亚市亚龙湾 | 花岗岩 |
| JF | 865 | 108°50′34.51″ | 18°45′38.86″ | 40 | 乐东县尖峰岭 | 花岗岩 |
| JFH | 808 | 108°50′15.14″ | 18°45′49.37″ | 20 | 乐东县尖峰岭 | 花岗岩 |
| JFM | 219 | 108°59′53.16″ | 18°44′37.76″ | 20 | 乐东县尖峰岭 | 花岗岩 |
| CH | 170 | 108°42′21.74″ | 19°20′8.64″ | 40 | 昌江县昌化大岭 | 花岗岩 |
| SJL | 95 | 108°50′59.06″ | 19°21′59.77″ | 21 | 昌江县三架岭 | 花岗岩 |
| LN | 44 | 110°24′37.47″ | 18°40′43.99″ | 26 | 万宁市乐南山 | 花岗岩 |
| HL | 194 | 108°48′45.49″ | 18°46′30.09″ | 57 | 东方市黑岭 | 花岗岩 |
| EXL | 980 | 109°06′40.31″ | 19°00′45.09″ | 40 | 东方市俄贤岭 | 石灰岩 |
| EX | 230 | 109°04′11.90″ | 18°59′58.00″ | 55 | 东方市俄贤岭 | 石灰岩 |
| NS:三亚市南山居群, Nanshan population in Sanya City; YL:三亚市亚龙湾居群, Yalongwan population in Sanya City; JF:乐东县尖峰岭引水头居群, Yinshuitou population on Jianfengling in Ledong County; JFH:乐东县尖峰岭回头湾居群, Huitouwan population on Jianfengling in Ledong County; JFM:乐东县尖峰岭叉河口居群, Chahekou population on Jianfengling in Ledong County; CH:昌江县昌化大岭居群, Changhuadaling population in Changjiang County; SJL:昌江县三架岭居群, Sanjialing population in Changjiang County; LN:万宁市乐南山居群, Lenanshan population in Wanning City; HL:东方市黑岭居群, Heiling population in Dongfang City; EXL:东方市俄贤岭居群, Exianling population in Dongfang City; EX:东方市俄贤岭南浪村居群, Nanlang population on Exianling in Dongfang City | ||||||

|
| 图 1 海南龙血树种群的地理位置 Fig. 1 Populations′ location of Dracaena cambodiana NS:三亚市南山居群, Nanshan population in Sanya City; YL:三亚市亚龙湾居群, Yalongwan population in Sanya City; JF:乐东县尖峰岭引水头居群, Yinshuitou population on Jianfengling in Ledong County; JFH:乐东县尖峰岭回头湾居群, Huitouwan population on Jianfengling in Ledong County; JFM:乐东县尖峰岭叉河口居群, Chahekou population on Jianfengling in Ledong County; CH:昌江县昌化大岭居群, Changhuadaling population in Changjiang County; SJL:昌江县三架岭居群, Sanjialing population in Changjiang County; LN:万宁市乐南山居群, Lenanshan population in Wanning City; HL:东方市黑岭居群, Heiling population in Dongfang City; EXL:东方市俄贤岭居群, Exianling population in Dongfang City; EX:东方市俄贤岭南浪村居群, Nanlang population on Exianling in Dongfang City |
在同一生境下同一树种的龄级和径级对环境的反应规律具有一致性[24]。因此, 采用空间推时间的方法, 用径级代替龄级分析海南龙血树种群的结构[2, 25]。根据生活史特征和野外调查数据的特点, 将海南龙血树种群划分为13个径级, 每个径级对应相应的龄级。基径(以下用D表示)≤3cm为第Ⅰ龄级, 3cm < D≤10cm为第Ⅱ龄级, D>10cm的海南龙血树个体以10cm作为级差进行龄级划分, 100cm < D≤150cm为第Ⅻ龄级, D>150cm为第ⅩⅢ龄级。为了方便分析海南龙血树各种群的年龄结构, 根据野外调查情况和海南龙血树的生长特点, 将Ⅰ、Ⅱ龄级(D≤10cm)的个体划分为幼龄个体, 将Ⅲ至Ⅵ龄级(10cm < D≤50cm)划分为中龄个体, Ⅶ至Ⅺ龄级(50cm < D≤100cm)划分为成龄个体, 将Ⅻ、ⅩⅢ龄级(D>100cm)划分为老龄个体。
1.2.2 种群年龄结构及动态采用陈晓德种群结构动态量化分析方法[26], 推导海南龙血树种群年龄结构的动态指数, 对种群动态(V)进行定量描述, Vn、Vpi、V′pi、V"pi取正、负、零值的意义分别反映种群或相邻年龄级个体数量的增长、衰退、稳定的动态关系[2, 25]。
种群龄级间动态指数(Vn)为:
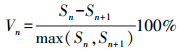
|
式中:Sn与Sn+1分别为第n和第n+1年龄级的个体数, -1≤Vn≤1。
种群年龄结构的数量变化动态指数(Vpi)为:
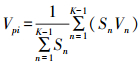
|
式中:K为种群年龄级数量, 此值越大, 说明种群的增长趋势越大。
考虑种群年龄级数量(K)及种群年龄级个体数(Sn)因素的种群结构动态指数(V′pi)为:
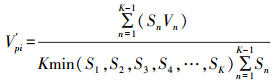
|
此值越大, 说明种群抗干扰能力越大, 种群稳定性越好。
1.2.3 静态生命表和存活曲线根据特定时间生命表的编制方法[27], 编制海南龙血树种群的静态生命表。静态生命表包含:x为年龄级; Ax为x龄级内现有个体数; 由于静态生命表是用同一时期收集到的种群中的所有个体的径阶编制而成, 它只反映了多个世代重叠的年龄动态历程中的一个特定时间, 而不是对种群全部生活史的追踪[28-29], 因此, 生命表的编制中会出现死亡率为负值的情况, 为此, 本文采取文献中的匀滑技术对数据Ax进行处理[27, 30-31], ax即为Ax匀滑修正后的数据; lx为x龄级开始时标准化存活个体数; dx为从x到x+1龄级间隔期内标准化死亡个体数; qx为从x到x+1龄级间隔期间的死亡率; Lx为从从x到x+1龄级间隔期间还存活的个体数; Tx为从x龄级到超过x龄级的个体总数; ex为进入x龄级个体的生命期望或平均期望寿命; Kx为消失率。通过实测Ax值, 计算以上各项的值, 它们的计算公式分别为:
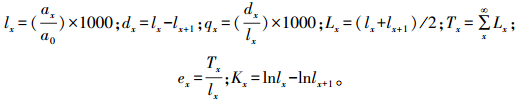
|
种群生存分析采用种群生存率函数(S(i)); 累计死亡率函数(F(i)); 死亡率密度函数(f(ti))和危险率函数(λ(ti))[30-31]。计算公式如下:
S(i)=S1×S2×S3×…×Si; F(i)=1-Si; 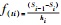
分析结果表明, 海南龙血树种群的年龄结构与其所处的生境关系密切。NS、YL、CH和LN种群分布在海边的花岗岩生境, 它们的特点是:种群中均存在一定比例的中龄、成龄和老龄个体。三亚NS种群和YL种群的年龄结构中均缺少幼龄个体, 幼龄个体占种群个体总数的比例分别为0和5%;中、成龄个体所占的比例较高, 分别为61%和90%;老龄个体占有一定的比例, 分别为39%和5%。由于幼龄个体的极度缺乏, NS和YL种群结构均属于衰退型。其中, NS种群幼龄个体缺失, 且拥有更高比例的老龄个体。因此, NS种群的衰退程度比YL种群更大。位于万宁乐南山的LN种群幼龄、中龄、成龄和老龄个体所占的比例分别为27%、46%、19%和8%, 属于稳定型种群。同样, 位于昌化大岭的CH种群幼、中、成、老龄的个体数量占种群个体总数量的比例分别为30%、25%、25%和20%, 其种群年龄结构也属于稳定型种群。相比之下, CH种群幼、中、成、老龄的个体数量分布较LN种群均匀, 种群的稳定性更好些, 而LN种群虽有老龄个体, 但其老龄个体比例仅为8%, 偏向于增长型(图 2)。

|
| 图 2 海南龙血树种群的年龄结构 Fig. 2 Age structure of Dracaena cambodiana populations Ⅰ: D(个体基径, basal diameter of individuals)≤3cm; Ⅱ: 3cm < D≤10cm; Ⅲ: 10cm < D≤20cm; Ⅳ: 20cm < D≤30cm; Ⅴ: 30cm < D≤40cm; Ⅵ: 40cm < D≤50cm; Ⅶ: 50cm < D≤60cm; Ⅷ: 60cm < D≤70cm; Ⅸ: 70cm < D≤80cm; Ⅹ: 80cm < D≤90cm; Ⅺ: 90cm < D≤100cm; Ⅻ: 100cm < D≤150cm; ⅩⅢ: D>150cm |
分布于花岗岩山间斜坡上的种群包括尖峰岭地区的JF、JFH、JFM和HL种群, 以及昌江三架岭的SJL种群, 它们的年龄结构无老龄个体, 成龄的个体极少, 种群中个体集中分布在幼、中龄, 种群为增长型。其中, JF种群的幼、中龄比例为53%和42%, 成龄的个体仅为5%;HL种群幼、中龄比例为33%和60%, 成龄的个体仅为7%;JFH和JFM幼、中龄比例均为100%。
位于石灰岩俄贤岭地区的EXL和EX种群与花岗岩非海边种群的年龄结构类似, 无老龄个体, 种群中个体集中分布在幼、中龄, 种群也属于增长型。但它们没有成龄个体, 中龄个体也较少, 种群个体主要集中在幼龄阶段, 如, EXL的幼龄个体占95%, 中龄仅有5%;EX幼龄个体占67%。
2.2 种群年龄结构的动态变化分析基于种群动态量化分析方法, 对海南龙血树11个种群相邻龄级间个体数量变化动态及各种群整体数量变化动态进行分析(表 2, 表 3)。结果显示, 海南龙血树各种群龄级间的个体数量均有明显的波动过程, 各种群的结构并不稳定, 11个种群在龄级间多出现了负增长动态, 出现龄级间衰退现象的次数在1/4以上, 最高达到3/5(JFH)。Ⅰ-Ⅳ龄级在大部分种群中存在, 但是10个种群(NS不存在Ⅰ和Ⅱ龄级)中, 有6个种群在Ⅰ和Ⅱ龄级之间出现衰退现象, 表明Ⅰ向Ⅱ龄级时有严重的环境筛, 导致Ⅰ向Ⅱ龄级过渡时极易出现高死亡率; Ⅲ和Ⅳ龄级之间的衰退现象较少, 9个种群(EX和EXL没有Ⅲ龄级)中仅2个种群出现负增长, 表明Ⅲ向Ⅳ龄级过渡时环境条件较为合适, 种群稳定。此外, 有6个种群存在Ⅵ向Ⅶ龄级过渡, 其中有5个种群的年龄结构动态指数等于或大于零, 仅NS的为负数, 且仅为11.11%, 表明这个阶段也是较为稳定的。
| 龄级/种群 Age class/Population |
NS | LN | YL | CH | JF | JFH | JFM | HL | SJL | EX | EXL |
| Ⅰ | — | 25.00 | -100.00 | 50.00 | 50.00 | -100.00 | -100.00 | -64.29 | 40.00 | -31.82 | -91.43 |
| Ⅱ | — | -57.14 | -50.00 | 75.00 | 42.86 | -16.67 | 27.27 | 42.86 | 33.33 | 18.18 | 94.29 |
| Ⅲ | 50.00 | 100.00 | -33.33 | 100.00 | 0.00 | 50.00 | 87.50 | -20.00 | 100.00 | — | — |
| Ⅳ | 0.00 | -100.00 | 33.33 | -100.00 | -50.00 | -25.00 | — | 20.00 | — | — | — |
| Ⅴ | -75.00 | -75.00 | 0.00 | -50.00 | 87.50 | 50.00 | — | 0.00 | -100.00 | — | — |
| Ⅵ | -11.11 | 25.00 | 0.00 | 66.67 | 100.00 | — | — | 87.50 | — | — | — |
| Ⅶ | 33.33 | 66.67 | 0.00 | -33.33 | -100.00 | — | — | -66.67 | — | — | — |
| Ⅷ | -45.45 | 100.00 | 50.00 | 66.67 | 100.00 | — | — | — | — | — | — |
| Ⅸ | 72.73 | -100.00 | 0.00 | -66.67 | -100.00 | — | — | — | — | — | — |
| Ⅹ | -50.00 | 100.00 | -50.00 | 66.67 | — | — | — | — | — | — | — |
| Ⅺ | -68.42 | -100.00 | 50.00 | -83.33 | — | — | — | — | — | — | — |
| Ⅻ | 31.58 | 0.00 | — | 66.67 | — | — | — | — | — | — | — |
| ⅩⅢ | — | — | — | — | — | — | — | — | — | — | — |
| Ⅰ: D(个体基径, basal diameter of individuals)≤3cm; Ⅱ: 3cm < D≤10cm; Ⅲ: 10cm < D≤20cm; Ⅳ: 20cm < D≤30cm; Ⅴ: 30cm < D≤40cm; Ⅵ: 40cm < D≤50cm; Ⅶ: 50cm < D≤60cm; Ⅷ: 60cm < D≤70cm; Ⅸ: 70cm < D≤80cm; Ⅹ: 80cm < D≤90cm; Ⅺ: 90cm < D≤100cm; Ⅻ: 100cm < D≤150cm; ⅩⅢ: D>150cm; “—”代表空值, “—” means null values | |||||||||||
| 种群/动态指数 Population/Dynamic index |
Vpi | V′pi |
| NS | 11.38 | 0.45 |
| CH | 42.98 | 3.31 |
| LN | 42.14 | 3.24 |
| YL | 7.41 | 0.67 |
| JF | 45 | 4.5 |
| JFH | 18.98 | 1.58 |
| JFM | 52.63 | 13.16 |
| HL | 17.63 | 2.52 |
| SJL | 20.00 | 3.33 |
| EXL | 84.89 | 14.15 |
| EX | 46.57 | 1.04 |
| Vpi:种群年龄结构的数量变化动态指数, dynamic index of quantitative change in population age structure; V′pi:考虑种群年龄级数量及种群年龄级个体数因素的种群结构动态指数, population structure dynamic index considering population age class number and individual number in each population age class | ||
种群总体动态变化指数分析结果表明, 11个种群均为增长型种群, 种群总体动态变化指数Vpi均大于0。在这11个种群中, EXL种群的Vpi值最大, 达到84.89%, 显示出最高的增长趋势, 同时V′pi值最大, 为14.15%, 种群抗干扰能力强, 种群稳定性好。其次, Vpi值大于40%以上的种群依次为JFM、EX、JF、CH和LN种群, 也显示出较高的增长趋势, 其中, JFM的Vpi值为52.63%, 是EXL种群Vpi值的0.62倍, 但它的V′pi值是13.16%, 抗干扰能力和稳定性近接EXL种群; EX种群虽然具较高的Vpi值, 但其抗干扰能力和稳定性较差, V′pi值为1.04%, 仅略高于NS。SJL、JFH和HL种群增长趋势较低, Vpi值分别为20%、18.98%和17.63%, 其中SJL的Vpi值为CH的0.47倍, 但V′pi值为3.33%, 略高于CH。NS和YL种群总体动态变化指数Vpi值虽然大于0, 但仅为11.38%和7.4%;V′pi值也极低, 仅为0.45%和0.67%, 种群波动大, 种群稳定性差, 种群抗干扰能力极低; 而且种群结构中幼龄个体极度缺乏, 成、老龄个体比例高。因此, NS和YL应属衰退型种群。
2.3 静态生命表及存活曲线海南龙血树各种群的静态生命表如表 4所示, 以龄级为x轴, 以lnlx、qx、kx为y轴绘制各种群的存活曲线、死亡率曲线和消失率曲线如图 3和图 4所示。种群存活曲线是对种群的死亡过程的直观描述, 由于匀滑处理会掩盖种群数量波动中的某些生态现象[32], 因此, 本文将匀滑处理前后的种群存活曲线进行对比。
| 种群 Population |
龄级 Age class |
Ax | ax | lx | lnlx | dx | qx | Lx | Tx | ex | Kx |
| NS | Ⅰ | 0 | 15 | 1000 | 6.91 | 133 | 0.13 | 933 | 5300 | 5.30 | 0.14 |
| Ⅱ | 0 | 13 | 867 | 6.76 | 133 | 0.15 | 800 | 4467 | 5.15 | 0.17 | |
| Ⅲ | 4 | 11 | 733 | 6.60 | 133 | 0.18 | 667 | 3700 | 5.05 | 0.20 | |
| Ⅳ | 2 | 9 | 600 | 6.40 | 67 | 0.11 | 567 | 3033 | 5.06 | 0.12 | |
| Ⅴ | 2 | 8 | 533 | 6.28 | 133 | 0.25 | 467 | 2467 | 4.63 | 0.29 | |
| Ⅵ | 8 | 6 | 400 | 5.99 | 0 | 0.00 | 400 | 2000 | 5.00 | 0.00 | |
| Ⅶ | 9 | 6 | 400 | 5.99 | 67 | 0.17 | 367 | 1600 | 4.00 | 0.18 | |
| Ⅷ | 6 | 5 | 333 | 5.81 | 0 | 0.00 | 333 | 1233 | 3.70 | 0.00 | |
| Ⅸ | 11 | 5 | 333 | 5.81 | 67 | 0.20 | 300 | 900 | 2.70 | 0.22 | |
| Ⅹ | 3 | 4 | 267 | 5.59 | 0 | 0.00 | 267 | 600 | 2.25 | 0.00 | |
| Ⅺ | 6 | 4 | 267 | 5.59 | 133 | 0.50 | 200 | 333 | 1.25 | 0.69 | |
| Ⅻ | 19 | 3 | 133 | 4.89 | 67 | 0.50 | 100 | 133 | 1.00 | 0.69 | |
| ⅩⅢ | 13 | 1 | 67 | 4.20 | 67 | 1.00 | 33 | 33 | 0.50 | 4.20 | |
| JF | Ⅰ | 14 | 14 | 1000 | 6.91 | 500 | 0.50 | 750 | 2714 | 2.71 | 0.69 |
| Ⅱ | 7 | 7 | 500 | 6.21 | 71 | 0.14 | 464 | 1964 | 3.93 | 0.15 | |
| Ⅲ | 4 | 6 | 429 | 6.06 | 71 | 0.17 | 393 | 1500 | 3.50 | 0.18 | |
| Ⅳ | 4 | 5 | 357 | 5.88 | 71 | 0.20 | 321 | 1107 | 3.10 | 0.22 | |
| Ⅴ | 8 | 4 | 286 | 5.65 | 71 | 0.25 | 250 | 786 | 2.75 | 0.29 | |
| Ⅵ | 1 | 3 | 214 | 5.37 | 71 | 0.33 | 179 | 536 | 2.50 | 0.41 | |
| Ⅶ | 0 | 2 | 143 | 4.96 | 0 | 0.00 | 143 | 357 | 2.50 | 0.00 | |
| Ⅷ | 1 | 2 | 143 | 4.96 | 71 | 0.50 | 107 | 214 | 1.50 | 0.69 | |
| Ⅸ | 0 | 1 | 71 | 4.27 | 0 | 0.00 | 71 | 107 | 1.50 | 0.00 | |
| Ⅹ | 1 | 1 | 71 | 4.27 | 71 | 1.00 | 36 | 36 | 0.50 | 4.27 | |
| JFH | Ⅰ | 0 | 8 | 1000 | 6.91 | 250 | 0.25 | 875 | 2188 | 2.19 | 0.29 |
| Ⅱ | 5 | 6 | 750 | 6.62 | 250 | 0.33 | 625 | 1375 | 1.83 | 0.41 | |
| Ⅲ | 6 | 4 | 500 | 6.21 | 250 | 0.50 | 375 | 750 | 1.50 | 0.69 | |
| Ⅳ | 3 | 2 | 250 | 5.52 | 125 | 0.50 | 188 | 375 | 1.50 | 0.69 | |
| Ⅴ | 4 | 1 | 125 | 4.83 | 0 | 0.00 | 125 | 188 | 1.50 | 0.00 | |
| Ⅵ | 2 | 1 | 125 | 4.83 | 125 | 1.00 | 63 | 63 | 0.50 | 4.83 | |
| JFM | Ⅰ | 0 | 12 | 1000 | 6.91 | 333 | 0.33 | 833 | 833 | 0.83 | 0.41 |
| Ⅱ | 11 | 8 | 667 | 6.50 | 250 | 0.38 | 542 | 542 | 0.81 | 0.47 | |
| Ⅲ | 8 | 5 | 417 | 6.03 | 333 | 0.80 | 250 | 250 | 0.60 | 1.61 | |
| Ⅳ | 1 | 1 | 83 | 4.42 | 83 | 1.00 | 42 | 42 | 0.50 | 4.42 | |
| CH | Ⅰ | 8 | 8 | 1000 | 6.91 | 125 | 0.13 | 938 | 5500 | 5.50 | 0.13 |
| Ⅱ | 4 | 7 | 875 | 6.77 | 125 | 0.14 | 813 | 4563 | 5.21 | 0.15 | |
| Ⅲ | 1 | 6 | 750 | 6.62 | 125 | 0.17 | 688 | 3750 | 5.00 | 0.18 | |
| Ⅳ | 0 | 5 | 625 | 6.44 | 125 | 0.20 | 563 | 3063 | 4.90 | 0.22 | |
| Ⅴ | 3 | 4 | 500 | 6.21 | 0 | 0.00 | 500 | 2500 | 5.00 | 0.00 | |
| Ⅵ | 6 | 4 | 500 | 6.21 | 125 | 0.25 | 438 | 2000 | 4.00 | 0.29 | |
| Ⅶ | 2 | 3 | 375 | 5.93 | 0 | 0.00 | 375 | 1563 | 4.17 | 0.00 | |
| Ⅷ | 3 | 3 | 375 | 5.93 | 125 | 0.33 | 313 | 1188 | 3.17 | 0.41 | |
| Ⅸ | 1 | 2 | 250 | 5.52 | 0 | 0.00 | 250 | 875 | 3.50 | 0.00 | |
| Ⅹ | 3 | 2 | 250 | 5.52 | 0 | 0.00 | 250 | 625 | 2.50 | 0.00 | |
| Ⅺ | 1 | 2 | 250 | 5.52 | 125 | 0.50 | 188 | 375 | 1.50 | 0.69 | |
| Ⅻ | 6 | 1 | 125 | 4.83 | 0 | 0.00 | 125 | 188 | 1.50 | 0.00 | |
| ⅩⅢ | 2 | 1 | 125 | 4.83 | 125 | 1.00 | 63 | 63 | 0.50 | 4.83 | |
| SJL | Ⅰ | 10 | 10 | 1000 | 6.91 | 400 | 0.40 | 800 | 1900 | 1.90 | 0.51 |
| Ⅱ | 6 | 6 | 600 | 6.40 | 200 | 0.33 | 500 | 1100 | 1.83 | 0.41 | |
| Ⅲ | 4 | 4 | 400 | 5.99 | 200 | 0.50 | 300 | 600 | 1.50 | 0.69 | |
| Ⅳ | 0 | 2 | 200 | 5.30 | 100 | 0.50 | 150 | 300 | 1.50 | 0.69 | |
| Ⅴ | 0 | 1 | 100 | 4.61 | 0 | 0.00 | 100 | 150 | 1.50 | 0.00 | |
| Ⅵ | 1 | 1 | 100 | 4.61 | 100 | 1.00 | 50 | 50 | 0.50 | 4.61 | |
| EXL | Ⅰ | 3 | 30 | 1000 | 6.91 | 567 | 0.57 | 717 | 967 | 0.97 | 0.84 |
| Ⅱ | 35 | 13 | 433 | 6.07 | 400 | 0.92 | 233 | 250 | 0.58 | 2.56 | |
| Ⅲ | 2 | 1 | 33 | 3.51 | 33 | 1.00 | 17 | 17 | 0.50 | 3.51 | |
| EX | Ⅰ | 15 | 22 | 1000 | 6.91 | 182 | 0.18 | 909 | 1955 | 1.95 | 0.20 |
| Ⅱ | 22 | 18 | 818 | 6.71 | 182 | 0.22 | 727 | 1045 | 1.28 | 0.25 | |
| Ⅲ | 18 | 14 | 636 | 6.46 | 636 | 1.00 | 318 | 318 | 0.50 | 6.46 | |
| LN | Ⅰ | 4 | 7 | 1000 | 6.91 | 143 | 0.14 | 929 | 5929 | 5.93 | 0.15 |
| Ⅱ | 3 | 6 | 857 | 6.75 | 143 | 0.17 | 786 | 5000 | 5.83 | 0.18 | |
| Ⅲ | 7 | 5 | 714 | 6.57 | 0 | 0.00 | 714 | 4214 | 5.90 | 0.00 | |
| Ⅳ | 0 | 5 | 714 | 6.57 | 143 | 0.20 | 643 | 3500 | 4.90 | 0.22 | |
| Ⅴ | 1 | 4 | 571 | 6.35 | 0 | 0.00 | 571 | 2857 | 5.00 | 0.00 | |
| Ⅵ | 4 | 4 | 571 | 6.35 | 143 | 0.25 | 500 | 2286 | 4.00 | 0.29 | |
| Ⅶ | 3 | 3 | 429 | 6.06 | 0 | 0.00 | 429 | 1786 | 4.17 | 0.00 | |
| Ⅷ | 1 | 3 | 429 | 6.06 | 143 | 0.33 | 357 | 1357 | 3.17 | 0.41 | |
| Ⅸ | 0 | 2 | 286 | 5.65 | 0 | 0.00 | 286 | 1000 | 3.50 | 0.00 | |
| Ⅹ | 1 | 2 | 286 | 5.65 | 0 | 0.00 | 286 | 714 | 2.50 | 0.00 | |
| Ⅺ | 0 | 2 | 286 | 5.65 | 143 | 0.50 | 214 | 429 | 1.50 | 0.69 | |
| Ⅻ | 1 | 1 | 143 | 4.96 | 0 | 0.00 | 143 | 214 | 1.50 | 0.00 | |
| ⅩⅢ | 1 | 1 | 143 | 4.96 | 143 | 1.00 | 71 | 71 | 0.50 | 4.96 | |
| YL | Ⅰ | 0 | 4 | 1000 | 6.91 | 250 | 0.25 | 875 | 5625 | 5.63 | 0.29 |
| Ⅱ | 1 | 3 | 750 | 6.62 | 0 | 0.00 | 750 | 4875 | 6.50 | 0.00 | |
| Ⅲ | 2 | 3 | 750 | 6.62 | 0 | 0.00 | 750 | 4125 | 5.50 | 0.00 | |
| Ⅳ | 3 | 3 | 750 | 6.62 | 250 | 0.33 | 625 | 3375 | 4.50 | 0.41 | |
| Ⅴ | 2 | 2 | 500 | 6.21 | 0 | 0.00 | 500 | 2750 | 5.50 | 0.00 | |
| Ⅵ | 2 | 2 | 500 | 6.21 | 0 | 0.00 | 500 | 2250 | 4.50 | 0.00 | |
| Ⅶ | 2 | 2 | 500 | 6.21 | 0 | 0.00 | 500 | 1750 | 3.50 | 0.00 | |
| Ⅷ | 2 | 2 | 500 | 6.21 | 250 | 0.50 | 375 | 1250 | 2.50 | 0.69 | |
| Ⅸ | 1 | 1 | 250 | 5.52 | 0 | 0.00 | 250 | 875 | 3.50 | 0.00 | |
| Ⅹ | 1 | 1 | 250 | 5.52 | 0 | 0.00 | 250 | 625 | 2.50 | 0.00 | |
| Ⅺ | 2 | 1 | 250 | 5.52 | 0 | 0.00 | 250 | 375 | 1.50 | 0.00 | |
| Ⅻ | 1 | 1 | 250 | 5.52 | 250 | 1.00 | 125 | 125 | 0.50 | 5.52 | |
| HL | Ⅰ | 5 | 16 | 1000 | 6.91 | 125 | 0.13 | 938 | 4094 | 4.09 | 0.13 |
| Ⅱ | 14 | 14 | 875 | 6.77 | 125 | 0.14 | 813 | 3156 | 3.61 | 0.15 | |
| Ⅲ | 8 | 12 | 750 | 6.62 | 125 | 0.17 | 688 | 2344 | 3.13 | 0.18 | |
| Ⅳ | 10 | 10 | 625 | 6.44 | 125 | 0.20 | 563 | 1656 | 2.65 | 0.22 | |
| Ⅴ | 8 | 8 | 500 | 6.21 | 63 | 0.13 | 469 | 1094 | 2.19 | 0.13 | |
| Ⅵ | 8 | 7 | 438 | 6.08 | 125 | 0.29 | 375 | 625 | 1.43 | 0.34 | |
| Ⅶ | 1 | 5 | 313 | 5.74 | 125 | 0.40 | 250 | 250 | 0.80 | 0.51 | |
| Ⅷ | 3 | 3 | 188 | 5.23 | 188 | 1.00 | 94 | 94 | 0.50 | 5.23 | |
| Ax:x龄级内现有的个体数, individual number of age class x; ax:Ax匀滑修正后的数据, the revised data of Ax; lx:x龄级开始时标准化存活个体数, the standardized number of surviving individuals of age class x; lnlx: lx的自然对数, the natural logarithm of lx; dx:从x到x+1龄级间隔期内标准化死亡个体数, the standardized number of death individuals from age class x to age class x+1; qx:从x到x+1龄级间隔期间的死亡率, mortality from age class xto age class x+1; Lx:从x到x+1龄级间隔期间还存活的个体数, the number of surviving individuals from age class x to age class x+1; Tx:从x龄级到超过x龄级的个体总数, the total individual number of age class xand age classes older than x; ex:进入x龄级个体的生命期望, life expectancy of individuals in the age class x; Kx:消失率, disappearance rate | |||||||||||

|
| 图 3 海南龙血树种群的存活曲线 Fig. 3 Survival curve of Dracaena cambodiana populations |

|
| 图 4 海南龙血树种群的死亡率(qx)和消失率(Kx) Fig. 4 Mortality and killing power of Dracaena cambodiana populations |
从存活曲线看, 11个海南龙血树种群均属于Deevey-Ⅰ型, 呈现凸型, 生命期望值ex随着龄级的增长而呈现不断下降的趋势, 表明海南龙血树的生存能力随着年龄的增加而下降, 而且其生命期望值通常在老龄阶段的下降趋势更显著, 种群幼龄和中幼龄个体的生命期望通常相差不大, 说明海南龙血树幼龄及中幼龄个体的生存能力较好, 接近老龄时个体生存能力较差。图 4也显示海南龙血树种群幼龄个体的存活率高于中龄个体和老龄个体, 幼龄个体的死亡率和消失率小于中、老龄个体(图 3)。
2.4 种群生存分析NS、YL、CH、EX、LN和HL种群累计死亡率的起始值较低, 函数值≤0.2;JF、JFH、JFM和SJL种群累计死亡率的起始值较高, 函数值在0.2—0.4之间; EXL种群累计死亡率的起始值最高, 函数值为0.67, 表明EXL种群所处的生境对龄级Ⅰ个体的环境筛作用较强。NS、JF、JFH、JFM、SJL、EXL种群的累计死亡曲线呈现出前期变化幅度大, 中后期变化幅度变小的特点; YL的累计死亡曲线呈现出波动性变化, Ⅰ到Ⅱ龄级、Ⅳ到Ⅵ龄级、Ⅷ到Ⅹ龄级的死亡累计率无变化, 其余各龄级间的变化幅度基本一致; CH、EX、LN、HL种群的累计死亡曲线呈现出早中晚龄期变化幅度基本一致的特点(图 5)。

|
| 图 5 海南龙血树种群的生存率(S(i))和累计死亡率(F(i)) Fig. 5 Survival rate and accumulated mortality rate curve of Dracaena cambodiana populations |
各种群的死亡密度函数曲线和危险率动态基本相似, 在Ⅰ龄级向Ⅱ龄级过渡过程中变化幅度较大, 后面各龄级间的变化趋势趋于平稳, 除EXL种群外, 其他种群的危险率函数呈现先减小后增大的趋势, 说明海南龙血树种群在整个生活史中中期稳定, 早期和晚期较为脆弱(图 6)。

|
| 图 6 海南龙血树种群的死亡密度函数(F(ti))和危险率(λ(ti)) Fig. 6 Morality density and hazard rate curve of Dracaena cambodiana populations |
植物种群龄级结构是其生物学特征与环境因素共同作用的结果[33-34]。李伟杰等人[23]对昌江昌化岭、三亚大小洞天、万宁东澳镇和东方江边乡及天安乡海南龙血树种群的年龄结构进行分析后认为这些种群的结构均为衰退型; 周文嵩等人[22]对昌江地区的海南龙血树种群年龄结构进行分析后认为昌江地区的海南龙血树种群也为衰退型。本文对海南龙血树不同生境下11个种群的年龄结构及种群结构动态指数分析的综合结果显示, 海南龙血树种群结构与种群所处的生境关系密切, 位于海边的NS和YL种群结构为衰退型, CH、LN种群为稳定型, 而位于非海边生境的其他7个种群为增长型。这一结果与以上两人的研究结果有一定差异。造成这种差异的原因可能是龄级划分标准、调查研究区域和研究方法的不同。由于海南龙血树种群中的很多个体都没有明显的主干, 无法获得个体胸径, 因此本研究中未用胸径, 而是采用基径划分海南龙血树的个体年龄级别, 而且将D≤10cm的个体划分为幼龄个体, 这一划分标准可能将更多的海南龙血树个体划分成了幼龄个体。本研究调查的区域更广, 处于尖峰岭保护区内的种群有4个, 在海拔800m以上的种群有2个, 它们不易受到人为干扰和破坏, 跟其他种群相比, 种群中的幼龄个体更多。本研究在尖峰岭JF种群中发现了较多数量的实生小苗, 这是在其他种群中没有的现象。此外, 本研究用年龄结构和种群结构动态指数两种方法来综合分析海南龙血树种群的结构, 比仅用单一的年龄结构分析可能更准确。
缺乏可更新的幼龄个体将导致种群趋于衰退[35]。海南龙血树的自然更新方式有种子更新、根蘖更新和桩蘖更新3种方式, 其中后两种属于无性繁殖; 目前的研究均显示海南龙血树种群中极少见到实生苗, 种群更新不良, 其更新方式以无性繁殖为主[20, 23]。本研究结果与前人的基本一致, 在调查到的11个海南龙血树种群中, 仅在JF、LN和SJL种群记录到2年生左右的实生苗, 其他种群中极少见到实生苗; 在CH种群中发现的幼苗均为根蘖苗或桩蘖苗, 是被砍伐后残留的海南龙血树根或桩上萌生的幼苗。海南龙血树种子萌发和幼苗生长对生境的依赖性较高, 需要一定的遮荫环境, 但遮荫度较好的NS和YL种群中没有发现幼苗的存在, 且幼龄个体都极度缺乏, 导致NS和YL种群成为衰退种群。在调查NS和YL种群时正值海南龙血树的结果期, 但调查过程中仅发现已枯萎的花序, 并未见到其果实, 因此推测是否是因为种群个体无法结实才导致NS和YL种群缺少幼苗和幼龄个体而成为衰退种群。这一推测还有待于繁殖生态学方面的研究来验证。JF、JFM、JFH、HL、EXL、EX和SJL种群仅有前3龄级的个体或缺少Ⅶ龄级以后的中老龄个体, 说明这7个种群的形成时间较短, 是正在发展的种群。
3.2 海南龙血树种群的动态趋势生命表是判断植物种群发展趋势的重要指标, 通过建立种群静态生命表、绘制存活曲线及生存函数曲线可以对海南龙血树种群动态进行分析。种群静态生命表和绘制存活曲线分析结果显示11个海南龙血树种群的存活曲线均属于Deevey-Ⅰ型, 海南龙血树幼龄个体的生存能力较好, 存活率高于中龄个体和老龄个体, 老龄个体生存能力较差; 但生存函数分析的结果显示海南龙血树种群生活史早期和晚期脆弱, 中期稳定, 这说明海南龙血树种群所处的生境更适合成年海南龙血树的生存, 虽然幼龄树自身的生存能力较强, 但现有的生境对幼龄个体的环境筛作用较强, 对幼龄树的生存不利, 这一点与郑道君等人[21]的研究结果一致。
植物种群的发展是一个漫长而复杂的过程, 涉及到物种内和物种间的竞争, 还涉及到物种与无机环境之间的适应过程。海南龙血树在种群发展的漫长过程中, 适应了严酷的岩石生境, 对种群分布及种群结构与动态的综合分析后可以看出, 位于海边生境的海南龙血树种群是古老种群, 处于稳定发展中或正趋于衰退; 而位于非海边的种群则是年轻种群, 是增长型种群。由此推断海南龙血树种群正在由海边的岩石生境向非海边的岩石生境转移, 这一推测还需要分子生物学和生物地理学方面的研究结果来验证。
3.3 海南龙血树种群的保护与恢复措施根据种群聚集程度的不同, 物种可分为补偿物种、中性物种和退偿物种三个类别, 补偿物种不易灭绝, 退偿物种容易灭绝, 当退偿物种的种群数量小于该物种的最小存活种群K0时, 该物种趋于灭绝[36]。海南龙血树在石灰岩生境中为集群分布, 在花岗岩生境中为均匀分布, 已被列入中国珍稀濒危保护植物名录, 应属于退偿物种。人类对海南龙血树野生资源的掠夺性采挖和对其生境的破坏造成海南龙血树野生资源的分布范围缩减, 分布点数量变少。从目前的文献和本研究的调查结果来看, 野生海南龙血树种群的分布点不会超过20个, 而且多数种群内的个体数量很少, 在100株以下; 目前种群估测数量能在500株以上的仅有三亚的大小洞天、昌江的昌化大岭和万宁的大洲岛, 其中大洲岛的海南龙血树种群是目前发现的保存最为完好的种群, 具有重要的科学研究价值。
综上所述, 为了防止海南龙血树这一物种的灭绝, 海南龙血树种群的保护工作刻不容缓。对濒危植物的保护目前主要有三种方法:就地保护、迁地保护和利用现代生物技术的保护[37]。近年来, 随着对极小种群野生植物拯救保护工程的进行, 一种新的野生植物保护方法“近地保护”被提了出来, 即在物种现有分布区(点)范围内的附近, 选择与其相同气候和相似的生境建立人工保护点[38]。此方法经中科院西双版纳热带植物园的比较论证, 认为它对稀有、濒危植物的保护, 大大优于通常的迁地保护, 是一种行之有效的新保护方法[39]。在就地保护中, 对于生活史中存在薄弱环节的濒危物种种群, 可以采用人工辅助措施, 如采用实验室辅助种子萌发, 待幼苗长至一定大小时, 再移植回原种群[40]。
根据本文的分析结果, 对海南龙血树种群的保护应采取多种保护方法相结合的方式。对于缺少幼龄个体的种群可以采用人工辅助措施获得该种群的实生幼苗, 然后将幼苗移植回该种群, 如NS、YL和CH种群可以采取此方式。对于处于易受人为干扰生境下的种群, 可以根据种群的情况选择迁地保护或近原地保护方式保护这类种群的遗传多样性, 如LN、EX、SJL种群可以采取此方式。而对于已在保护区内海南龙血树种群, 如EXL、JF、JFH、JFM、HL种群, 可以根据种群的实际情况, 利用人工辅助措施促进种群的天然更新能力, 如扰动土壤以促进种子落入土壤、伐除母树和幼苗周围的乔灌木、增加土壤养分等, 可以促进母树的结实, 提高幼苗的生长量, 增加幼苗的密度。加强对海南龙血树遗传多样性的研究, 明确其濒危的内在原因对海南龙血树种群的保护极其重要。建立异地种质资源圃来保存现存海南龙血树的核心种质, 以防现存种群因人为或自然原因而突然消失从而造成海南龙血树遗传多样性进一步丢失。
| [1] | 姜汉侨, 段昌群, 杨树华, 王崇云, 苏文华. 植物生态学.第二版. 北京: 高等教育出版社, 2010. |
| [2] | 韩路, 王家强, 王海珍, 宇振荣. 塔里木河上游胡杨种群结构与动态. 生态学报, 2014, 34(16): 4640–4651. |
| [3] | 汪洋, 冷艳芝, 苏长江, 宋丛文, 程德华, 操英南, 张敏, 付翠林. 恩施天然红椿种群结构及空间分布格局. 浙江农林大学学报, 2016, 33(1): 17–25. |
| [4] | 刘普幸. 疏勒河中下游绿洲胡杨种群结构与动态研究. 自然资源学报, 2011, 26(3): 429–439. |
| [5] | Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park, Chile. Vegetatio, 1992, 98(1): 13–22. DOI:10.1007/BF00031633 |
| [6] | Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercus garryana) in British Columbia, Canada. Forest Ecology and Management, 2000, 137(1/3): 209–219. |
| [7] | 广东省植物研究所. 海南植物志(第四卷). 北京: 科学出版社, 1977: 155. |
| [8] | Chen X Q, Turland N J. Dracaena Vandelli ex Linnaeus//Wu Z Y, Raven P H. Flora of China, Vol. 24. Beijing: Science Press and St. Louis: Missouri Botanical Garden Press, 2000: 215-217. |
| [9] | 中国科学院植物研究所. 中国数字植物标本馆. http://www.nhpe.org/cvh/. [2017-08-12] |
| [10] | 国家环境保护局, 中国科学院植物研究所. 中国珍稀濒危保护植物名录(第一册). 北京: 科学出版社, 1987: 1–38. |
| [11] | 罗赫, 黄世满. 海南龙血古树优势群丛的调查. 热带林业, 2007, 35(4): 50-51, 41-41. |
| [12] | 郑道君, 云勇, 吴宇佳, 李海文, 张治礼. 海南龙血树野生资源分布及其与水热关系的分析. 热带亚热带植物学报, 2012, 20(4): 326–332. |
| [13] | 郑道君, 谢良商, 王盈, 张治礼, 张文. 中国血竭基源植物的研究与利用. 中国野生植物资源, 2009, 28(6): 15–20. |
| [14] | 梅文莉, 戴好富, 吴娇, 庄令, 洪葵. 海南龙血树抗肿瘤新用途研究. 中药材, 2005, 28(10): 871–873. |
| [15] | 杨先会, 邓世明, 范丽霞. 海南龙血树植物资源的开发利用. 海南大学学报:自然科学版, 2004, 22(3): 270–272. |
| [16] | 杨本鹏, 张树珍, 蔡文伟, 宋启示, 王兴红, 王东, 杨学, 顾丽红, 冯翠莲, 王俊刚, 罗遵喜. 海南龙血树组织培养过程中血竭形成的诱导. 热带作物学报, 2009, 30(2): 181–185. |
| [17] | Luo Y, Dai H F, Wang H, Mei W L. Chemical constituents from dragon's blood of Dracaena cambodiana. Chinese Journal of Natural Medicines, 2011, 9(2): 112–114. |
| [18] | 罗冠勇, 钟云芳, 宋希强, 戴好富, 杨冬华. 海南龙血树基于愈伤组织诱导途径的组培快繁技术体系优化. 热带作物学报, 2012, 33(10): 1824–1828. |
| [19] | Zheng D J, Xie L S, Zhu J H, Zhang Z L. Low genetic diversity and local adaptive divergence of Dracaena cambodiana (Liliaceae) populations associated with historical population bottlenecks and natural selection:an endangered long-lived tree endemic to Hainan Island, China. Plant Biology, 2012, 14(5): 828–838. DOI:10.1111/plb.2012.14.issue-5 |
| [20] | 郑道君, 李海文, 云勇, 张浪, 张治礼. 海南龙血树种群生境及自然更新能力调查. 热带亚热带植物学报, 2010, 18(6): 627–632. |
| [21] | 郑道君, 吴宇佳, 云勇, 姜殿强, 陈宣, 张治礼. 濒危植物海南龙血树种子萌发及其环境适应性分析. 热带亚热带植物学报, 2016, 24(1): 71–79. |
| [22] | 周文嵩, 冯丹丹, 李东海, 陶楚, 万春红, 邢莎莎, 杨小波. 海南昌江县濒危药用植物的种群动态. 热带生物学报, 2014, 5(4): 392–399. |
| [23] | 李伟杰, 肖艳, 杨海建, 陈沂章, 曾琳, 何明军, 郑希龙. 海南龙血树种群的年龄结构及分布格局. 氨基酸和生物资源, 2017, 39(1): 17–24. |
| [24] | 张维, 李海燕, 赖晓辉, 杨允菲. 新疆天山峡谷不同坡向野核桃种群分布格局. 应用生态学报, 2016, 27(10): 3105–3113. |
| [25] | 李肇晨, 陈永富, 洪小江, 韩文涛, 李小成. 海南霸王岭陆均松种群年龄结构与点格局分布特征. 生态学杂志, 2015, 34(6): 1507–1515. |
| [26] | 陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214–217. |
| [27] | 江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992. |
| [28] | Silvertown J W. 植物种群生态学导论. 祝宁, 王义弘, 陈文斌, 译. 哈尔滨: 东北林业大学出版社, 1987. |
| [29] | 吴承祯, 洪伟. 杉木数量经营学引论. 北京: 中国林业出版社, 2000. |
| [30] | 毕晓丽, 洪伟, 吴承祯, 闫淑君, 蓝斌. 黄山松种群统计分析. 林业科学, 2002, 38(1): 61–67. |
| [31] | 苏建荣, 刘万德, 缪迎春, 李帅锋, 郎学东. 云南红豆杉种群生物学.第三十一版. 北京: 科学出版社, 2016. |
| [32] | 李晓笑, 王清春, 崔国发, 杨传东. 濒危植物梵净山冷杉野生种群结构及动态特征. 西北植物学报, 2011, 31(7): 1479–1486. |
| [33] | Svensson J S, Jeglum J K. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline. Forest Ecology and Management, 2001, 151(1/3): 67–79. |
| [34] | 王金山, 哈力克·玉米提, Cyffka B, 吾斯曼·库尔班, 彭树宏. 塔里木河下游胡杨林胸径结构及林木分布特征. 植物学报, 2008, 25(6): 728–733. |
| [35] | 解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 2014, 34(15): 4272–4279. |
| [36] | 李晓波, 王克, 范猛, 李宪高. 最小存活种群的确定与生物多样性保护. 东北师大学报:自然科学版, 2001, 33(3): 86–90. |
| [37] | 吴成贡, 蒋昌顺. 濒危植物保护生物学技术研究进展. 华南热带农业大学学报, 2006, 12(3): 49–51. |
| [38] | 国家林业局野生动植物保护与自然保护区管理司, 云南省林业厅, 中国科学院昆明植物研究所. 云南省极小种群野生植物保护实践与探索. 昆明: 云南科技出版社, 2013: 1–100. |
| [39] | 许再富, 高江云, 李保贵, 周惠芳. 国家重点保护植物"迁地"与"近地"保护有效性的比较研究//中国植物园(第十五期). 西安: 中国植物学会, 2011: 6–15. |
| [40] | 胡世俊. 生境片断化与隔离对濒危植物缙云卫矛(Euonymus chloranthoides Yang)种群特征的影响[D]. 重庆: 西南大学, 2007 http://cdmd.cnki.com.cn/article/cdmd-10635-2007130951.htm |