 2018, Vol. 38
2018, Vol. 38文章信息
- 曾文豪, 石慰, 唐一思, 郑维艳, 曹坤芳.
- ZENG Wenhao, SHI Wei, TANG Yisi, ZHENG Weiyan, CAO Kunfang.
- 广西地区喀斯特与非喀斯特山地森林树木物种多样性及系统发育结构比较
- Comparison of the species diversity and phylogenetic structure of tree communities in karst and non-karst mountains in Guangxi
- 生态学报. 2018, 38(24): 8708-8716
- Acta Ecologica Sinica. 2018, 38(24): 8708-8716
- http://dx.doi.org/10.5846/stxb201808021643
-
文章历史
- 收稿日期: 2018-08-02
- 修订日期: 2018-10-12





2. 广西大学, 亚热带农业生物资源保护与利用国家重点实验室, 南宁 530004
2. State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Guangxi University, Nanning 530004, China
生物多样性的形成和维持机制一直是生态学研究的核心问题之一[1-3]。很长时间里, 生态学家基于环境因素对群落多样性分布格局的形成进行了大量的探索, 取得许多重要的发现。生物的代谢和生长离不开温度, 物种的分布范围受环境最低温的限制[4];一般情况下, 物种多样性随温度的升高而升高[5]。生态学代谢理论认为, 较高的环境温度能够促进物种的新陈代谢, 进而提高地区的物种丰富度[6]。土壤是植物生长的基础, 土壤的质量与物种丰富度的关系密切相关[7-8]。任学敏等在秦岭太白山巴山冷杉(Abies fargesii)-糙皮桦(Betula utilis)混交林的调查中发现物种丰富度随土壤pH值升高而增加, 随岩石盖度升高而减小[9]。Dornbush等研究发现高草草原土壤厚度的增加有利于提高物种丰富度[10]。海拔作为环境因子的一个集合体, 它综合反映着环境温度、降雨、光照和土壤等梯度的变化, 这些因子影响群落物种组成以及系统发育结构。卢孟孟等对云南哀牢山亚热带森林的研究中发现群落系统发育结构随着海拔梯度的升高由聚集逐渐变为发散或聚集程度降低[11]。Culmsee在研究马来西亚亚热带森林时发现, 从低海拔到高海拔系统发育结构由发散逐渐转向聚集[12]。虽然针对不同区域的研究呈现的结果有所差异, 但是这些研究足以说明环境因子对物种的形成及群落的系统发育结构产生影响。目前, 群落系统发育学被广泛用于群落的构建机制和进化历史研究, 在一个群落的演化过程中, 如果群落的构建是以环境选择为主要机制, 其系统发育结构表现为聚集;如果是以物种竞争为主要构建机制, 群落系统发育结构表现为离散;如果是以随机过程为主要构建机制, 则群落的系统发育结构呈随机状态[13-14]。
我国的亚热带地区受海洋季风影响, 光热水资源丰富、土壤肥沃, 地带性植被为常绿阔叶林, 其生态系统生产力高[15]。广西处于低纬度地区, 自北向南占据着亚热带(包括中亚热带和南亚热带)及逐渐向热带过渡的区域, 依次分布着中亚热带常绿阔叶林带、南亚热带常绿季雨林带、北热带季节性雨林带三个植被带;由东向西降水减少, 广西西部地区为湿润的气候区向半湿润气候区的过渡区[16]。
喀斯特地貌以碳酸盐为主的岩溶构造, 多表现为峰林、峰丛、洼地、峡谷等地貌特征;具有土层浅薄偏碱性、富钙镁, 岩石裸露, 土被不连续, 土壤蓄水能力低等特点[17-18]。复杂独特的喀斯特地质气候条件构成了许多的适合不同植物生存的小生境, 决定了其植物群落的特殊性[19-20]。通过对喀斯特森林和非喀斯特森林树木物种多样性的研究, 揭示两种生境森林群落的形成机制的差别, 对森林资源的保护和利用提供有价值参考[21]。独特的喀斯特生境几十年来吸引了众多学者对其森林的研究, 但大多数都是针对个别保护区开展的研究, 比较少的从较大区域尺度上开展对喀斯特和非喀斯特森林群落构建机制、系统发育多样性之间差异的探讨[22-24]。本研究以广西地区喀斯特与非喀斯特山地森林为研究对象, 结合系统发育学的在生态学中的应用探讨下列两个科学问题:(1)海拔≤1000 m喀斯特和非喀斯特森林以及海拔>1000 m的非喀斯特森林之间乔木植物科组成的差异。(2)这三类生境的森林物种丰富度和系统发育多样性的差异及其与生长地点年平均温度的关系, 讨论不同生境山地森林群落的主要构建机制。
1 研究区概况广西地处华南, 104°26′—112°04′E, 20°54′—26°24′N, 陆地总面积23.7万km2, 其中喀斯特分布面积占广西总面积35%, 主要分布于桂西南、桂西北、桂中和桂东北(图 1)。广西地区水热同期且丰沛, 各地年日照时数1169—2219 h, 各地年均气温16.5—23.1℃, 累年极端最高温33.7—42.2℃, 累年极端最低气温为-8.4—2.9℃, 日均温≥10℃的积温在4995—8328℃之间。常年降水量1086.3—2754.5 mm, 季节分配不均[16]。

|
| 图 1 样地分布图 Fig. 1 The distribution map of community sample plots (Partially overlapping) (部分样地重叠) |
本研究的山地森林群落数据主要来源于《广西植被志要》[25], 将收集到的样地分为三类生境, 一是海拔≤1000 m非喀斯特样地, 二是海拔>1000 m非喀斯特样地, 三是海拔≤1000 m喀斯特样地。各个生境样地信息如表 1, 样地地理分布如图 1。从国家气象网收集到的广西地区90个气象站30年气象数据, 建立年平均温度与经度(G)、纬度(L)和海拔高度(H)的回归模型(年平均温=72.794-0.736×L-0.31×G-0.005×H, R2=0.938, P < 0.001), 以此为基础, 结合每个样地点的经度、纬度和海拔计算出每个样地点的年平均温度[26]。
| 生境 Habitat |
样方数量/个 Number of plot |
样方海拔范围 Elevation range/m |
样方面积 Area/m2 |
样方累计面积 Cumulative area/m2 |
样方为400 m2的比例 Proportion of plots with 400 m2/% |
| 海拔≤ 1000 m喀斯特Karst with an elevation of ≤ 1000 m | 80 | 130—1000 | 300—1200 | 38000 | 57.5 |
| 海拔≤ 1000 m非喀斯特Non-karst with an elevation of ≤ 1000 m | 225 | 10—1000 | 300—1000 | 108100 | 57.3 |
| 海拔>1000 m非喀斯特Non-karst with an elevation of > 1000 m | 60 | 1300—2020 | 300—1000 | 31900 | 46.7 |
考虑到部分样方面积的不一致性, 采用Gleason指数来表示物种多样性的测度[27], 比较和分析广西地区喀斯特和非喀斯特森林树木物种多样性的差异。采用Faith系统发育多样性指数PD(phylogenetic diversity)来分析群落系统发育多样性[28], 使用协方差分析的方法检验不同生境群落Gleason物种多样性指数与系统发育多样性线性回归线的差异。Gleason指数公式如下:
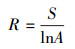
|
式中, A表示所研究的面积(m2);S表示面积A内的种数(个)。
2.2.2 系统发育树的构建和系统发育指数的选择将收集到的样地树木的拉丁名名录输入到Phylomatic平台[29](http://phylodiversity.net/phylomatic/), 利用Phylomatic中数据库构建物种系统发育树, 该平台以被子植物分类系统APG Ⅲ为基础, 整合Zanne的进化树信息, 输出带枝长的系统进化树[30]。净亲缘关系指数(net relatedness index, NRI)和最近分类群指数(nearest taxon index, NTI)被广泛用于研究群落的系统发育结构[13, 31]。当NRI、NTI>0, 说明群落物种组成在系统发育结构上聚集, 群落由亲缘关系比较近的物种组成, 生境过滤是群落构建的主要因素;当NRI、NTI < 0, 说明群落系统发育结构发散, 群落由亲缘关系较远的种构成, 竞争排斥是群落构建的主要控制因素;当NRI、NTI=0, 说明群落的系统发育结构呈随机状态, 环境因素和生物因素共同维持群落生物多样性[13]。使用t-检验分析不同生境间指数是否存在显著性差异。NRI和NTI计算公式如下:

|
式中, MPD表示物种间的平均成对系统发育距离, MPDsample表示实际观察到的MPD值, MPDrandsample代表随机产生的999个随机组合的群落的MPD值, SD(MPDrandsample)为这些随机值的标准差。MNTD表示平均最近种间系统发育距离, MNTDsample表示群落内亲缘关系最近的物种之间的平均系统发育距离, MNTDrandsample表示随机产生的999个随机组合的群落的MNTD值, SD(MNTDrandsample)为这些随机值的标准差。
3 结果 3.1 喀斯特和非喀斯特森林科与物种数的统计将样地群落按照前面所述3种不同生境统计群落的科组成及其物种数情况。发现所研究群落样地共有树木936个种, 按照中国植物志分类系统, 他们隶属于98科361个属, 其中从科的组成上看广西地区海拔≤1000 m的非喀斯特群落中共有87个科, 占所有研究群落植物科的88.8%, 其中含有4种以上的有樟科、壳斗科、茜草科、桑科、山矾科等44科, 共计709个种, 占所研究群落所有物种种数的75.7%。海拔>1000 m非喀斯特森林共有61科, 占所研究群落所有树木科的62.2%, 其中含4种以上的有樟科、壳斗科、山茶科、杜鹃花科等20科, 共计360个种, 占所研究群落所有物种种数的38.5%。喀斯特森林共有70科, 占所研究群落所有植物科的71.4%, 其中含有4种以上的有大戟科、樟科、豆科、桑科、茜草科等26科, 共计308个种, 占所研究群落所有物种种数的32.9%(图 2)。

|
| 图 2 广西海拔≤1000 m非喀斯特、海拔>1000 m非喀斯特和海拔≤1000 m喀斯特天然林科与物种数组成及其所占比例 Fig. 2 Size of families (%) containing over 4 species of 3 habitats: non-karst with elevation of ≤1000 m, non-karst with elevation of >1000 m and karst with elevation of ≤1000 m 科前面数字代表他们所包含的物种数 |
3种生境比较来看, 海拔≤1000 m非喀斯特森林含有4种以上的植物科比海拔>1000 m的非喀斯特中山森林多了24科, 其中樟科、壳斗科、山茶科、杜鹃花科等都是两种生境的重要组成科。大戟科为喀斯特森林含有种数最多的科, 其中出现在非喀斯特森林中的山茶科和杜鹃花科等没有在喀斯特森林出现。
3.2 喀斯特和非喀斯特森林物种丰富度及系统发育多样性海拔>1000 m非喀斯特中山生境的Gleason指数的平均值显著高于另外两种生境的森林, 海拔≤1000 m非喀斯特和喀斯特两种生境的Gleason指数没有显著差异。其中, 喀斯特森林样地分布范围比海拔≤1000 m非喀斯特森林要窄, 前者主要分布于年平均温18—22℃的生境之间, 后者主要分布于年平均温15—23℃的生境之间, 海拔大于1000 m非喀斯特森林样地分布范围的年平均温比低海拔两种生境都要低, 且其Gleason指数与年平均温表现为一定的反相关(图 3)。

|
| 图 3 三种生境森林样地Gleason多样性指数与年平均温散点图 Fig. 3 Lack of correlation between Gleason diversity index and mean annual temperature in the forest communities in all of the three habitats 黒点R2=0.2%, P=0.74;红点R2=2.3%, P=0.02;绿点R2=20.1%, P=0.0003。小图展示三种生境的平均多样指数和标准差, 不同字母表示差异显著(P < 0.01) |
3种生境的森林群落系统发育多样性PD指数与Gleason指数都表现为显著的线性正相关关系。协方差分析结果表明PD指数与Gleason指数的关系的回归线斜率在3种生境森林之间的没有表现出显著差异, 截距表现出显著差异(P < 0.05)。在相同的Gleason指数情况下, 海拔≤1000 m喀斯特森林样地系统发育多样性比海拔≤1000 m非喀斯特森林样地离散程度小, 海拔≤1000 m非喀斯特森林样地系统发育多样性PD指数比>1000 m的要高(图 4)。

|
| 图 4 三种生境森林样地系统发育多样性PD指数与Gleason相关散点图 Fig. 4 The correlation between Gleason diversity index and phylogenetic diversity index in the forest communities in all of the three habitats 黒点R2=90%, P < 0.0001;红点R2=95%, P < 0.0001;绿点R2=94%, P < 0.0001 |
从净亲缘关系指数NRI来看, 除了小部分样地表现为系统发育发散外, 3种生境其他大部分样地均表现为系统发育聚集。喀斯特森林和中山地带的非喀斯特森林绝大部分的样地表现为系统发育聚集, 而低海拔非喀斯特森林有约25.7%的样地表现为系统发育离散(图 5)。

|
| 图 5 净亲缘关系指数(NRI)、最近分类群指数(NTI)与年平均温度和海拔的变化 Fig. 5 Variation in NRI and NTI with (A and B) mean annual temperature (MAT) and elevation A图:黒点P=0.47;红点R2=5%, P=0.0007;绿点P=0.99; B图:黒点P=0.08;红点R2=5.7%, P=0.0003;绿点P=0.26 |
从最近分类群指数NTI来看, 3种生境的大部分样地都表现为群落系统发育聚集(图 5)。但是与系统发育结构NRI不同的是两种非喀斯特生境的森林群落有部分样地的系统发育结构(NTI)聚集程度比喀斯特森林的要大。
4 讨论 4.1 喀斯特与非喀斯特森林群落物种丰富度和系统发育多样性差异海拔>1000 m非喀斯特森林的Gleason指数总体上都要比海拔≤1000 m非喀斯特和喀斯特两种森林的Gleason指数稍高, 同时海拔≤1000 m和>1000 m两种非喀斯特生境的森林含有4种以上的植物科组成上有所不同, 并且前者含有4种以上的科比后者要多, 造成这差异的原因可能有:第一, 在一定海拔范围内, 海拔>1000 m的环境因子并不能对物种的分布造成限制影响, 何艳华等在对山西省五鹿山森林物种多样性海拔格局研究中发现在一定范围内物种丰富度随着海拔的升高而逐渐增加, 在海拔1500 m左右达到最大[32]。卢孟孟等在对云南哀牢山亚热带森林的调查同样呈现在一定海拔范围内物种丰富度随着海拔的升高而逐渐增加的结果, 并在海拔1700 m左右达到最大[11]。朱彪等在对广西猫儿山的森林调查中发现类似的结果, 物种丰富度在海拔1350 m以下变化不明显, 但在1350 m以上随着海拔的升高物种丰富度呈下降趋势[33]。在我们的研究中, 海拔大于1000 m生境的样地中, 海拔范围在1000 m至1500 m的样地占了72.1%, 海拔范围在1000 m至1700 m的样地占了91.8%。第二, 由于较低海拔范围内, 受人为干扰较大, 降水较少, 土壤元素含量较低, 可能会导致物种丰富度较低。随着海拔的升高, 降水增加, 气温虽然降低但是不太寒冷, 土壤中养分含量有所增加, 导致适合生存的物种增多[32]。海拔>1000 m非喀斯特森林样地Gleason指数与年均温呈反相关的关系, 这与前人对物种丰富度在一定范围内随温度的升高而升高的基本认识相勃[34], 造成这一结果的原因可能与中山地区一些特殊的小生境有关。海拔≤1000 m非喀斯特森林和喀斯特森林在科属组成有所不同, 前者种数最多的科为樟科, 后者为大戟科, 其中出现在非喀斯特森林的山茶科和杜鹃花科等都没有在喀斯特森林中出现。造成这种差异的原因可能是由于喀斯特特殊的地质和地形所致, 喀斯特基质漏水, 土层薄, 岩石裸露, 容易缺水, 并且土壤富积钙, 中性或弱碱性, 使得嫌钙植物不能生长, 而一些喜酸土、耐酸土植物也不能在喀斯特生境分布。
协方差分析表明三种生境森林群落PD指数与Gleason指数的回归线的截距显著差异。总体上看来Gleason指数相同的情况下海拔≤1000 m喀斯特森林样地系统发育多样性PD指数比海拔≤1000 m非喀斯特生境的森林样地离散程度小, 海拔≤1000 m非喀斯特森林样地系统发育多样性PD指数比>1000 m的要高, 推测海拔≤1000 m非喀斯特森林生境比海拔>1000 m的非喀斯特生境更适合不同亲缘关系的树种生存。
4.2 三种生境森林群落系统发育结构从净亲缘关系指数(NRI)和最近分类群指数(NTI)来看, 3种生境的大部分样地都倾向于系统发育聚集。这说明生境过滤对于这3种生境的群落构建起主要作用, 使得发育关系相近物种得以生存, 从而表现出系统发育聚集[35]。但是, 无论从NRI指数还是NTI指数来看, 海拔≤1000 m喀斯特(NRI 3.8%, NTI 15%), 海拔>1000 m非喀斯特(NRI 11.5%, NTI 8.2%)两种生境森林群落发生系统发育发散的比例都比海拔≤1000 m非喀斯特(NRI 25.7%, NTI 20.4%)低, 揭示生境过滤在这两个生境中表现的更为明显。从净亲缘关系指数(NRI)来看, 海拔≤1000 m非喀斯特森林样地呈现系统发育聚集比例(74.3%)比海拔>1000 m非喀斯特(88.5%)生境要小, 这与前人的研究结果类似, 之前的研究表明, 系统发育聚集程度随着海拔的升高而逐渐加强或表现为单峰模式[36];而喀斯特森林的群落系统发育结构聚集程度要比其他两种生境强(96.3%), 这可能与喀斯特生境选择压力大相关。从最近分类群指数(NTI)来看, 得出的结果与净亲缘关系指数(NRI)有所差别, 其中非喀斯特海拔>1000 m的系统发育聚集程度(91.8%)要比另外两种生境要强。在前人的研究中也有类似的结果, 随着海拔的升高, 温度、水分和土壤等都发生变化, 系统发育结构逐渐聚集[37]。
4.3 结语本研究通过分析广西地区喀斯特和非喀斯特山地森林群落的物种多样性和系统发育结构发现, 海拔>1000 m非喀斯特生境物种丰富度比另外两种生境要高;海拔≤1000 m的喀斯特和非喀斯特两种生境物种丰富度没有显著差异, 说明喀斯特的生境虽比非喀斯特生境严峻, 但喀斯特形成许多水热条件差异的小生境使不同类型的物种得以生长, 从而保持喀斯特生境也具有较高的物种丰富度。同时, 在3种生境中大部分样地的森林群落都表现出系统发育聚集的现象, 较少样地表现出发散或随机的现象, 说明生境过滤是广西地区天然林群落构建的主要驱动因素。研究结果揭示中山地带的森林树木物种多样性与低海拔森林相似, 喀斯特森林具有许多特有植物, 表明它们与低海拔非喀斯特生境的森林一样具有高的生物资源保护价值。
| [1] | Francis A P, Currie D J. A globally consistent richness-climate relationship for angiosperms. The American Naturalist, 2003, 161(4): 523–536. DOI:10.1086/368223 |
| [2] | Tilman D. Niche tradeoffs, neutrality, and community structure:a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 10854–10861. DOI:10.1073/pnas.0403458101 |
| [3] | Kim D, Yu K B. A conceptual model of coastal dune ecology synthesizing spatial gradients of vegetation, soil, and geomorphology. Plant Ecology, 2009, 202(1): 135–148. DOI:10.1007/s11258-008-9456-4 |
| [4] | Wright D H. Species-energy theory:an extension of species-area theory. Oikos, 1983, 41(3): 496–506. DOI:10.2307/3544109 |
| [5] | Austin M P, Pausas J G, Nicholls A O. Patterns of tree species richness in relation to environment in southeastern New South Wales, Australia. Australian Journal of Ecology, 1996, 21(2): 154–164. DOI:10.1111/aec.1996.21.issue-2 |
| [6] | Stegen J C, Enquist B J, Ferriere R. Advancing the metabolic theory of biodiversity. Ecology Letters, 2009, 12(10): 1001–1015. DOI:10.1111/ele.2009.12.issue-10 |
| [7] | 尚文艳, 吴钢, 付晓, 刘阳. 陆地植物群落物种多样性维持机制. 应用生态学报, 2005, 16(3): 573–578. DOI:10.3321/j.issn:1001-9332.2005.03.036 |
| [8] | Jones M M, Tuomisto H, Borcard D, Legendre P, Clark D B, Olivas P C. Explaining variation in tropical plant community composition:influence of environmental and spatial data quality. Oecologia, 2008, 155(3): 593–604. DOI:10.1007/s00442-007-0923-8 |
| [9] | 任学敏, 杨改河, 王得祥, 秦晓威, 刘振学, 赵双喜, 白宇. 环境因子对巴山冷杉-糙皮桦混交林物种分布及多样性的影响. 生态学报, 2012, 32(2): 605–613. |
| [10] | Dornbush M E, Wilsey B J. Experimental manipulation of soil depth alters species richness and co-occurrence in restored tallgrass prairie. Journal of Ecology, 2010, 98(1): 117–125. DOI:10.1111/jec.2009.98.issue-1 |
| [11] | 卢孟孟, 黄小翠, 慈秀芹, 杨国平, 李捷. 沿海拔梯度变化的哀牢山亚热带森林群落系统发育结构. 生物多样性, 2014, 22(4): 438–448. |
| [12] | Culmsee H, Leuschner C. Consistent patterns of elevational change in tree taxonomic and phylogenetic diversity across Malesian mountain forests. Journal of Biogeography, 2013, 40(10): 1997–2010. |
| [13] | Webb C O, Ackerly D D, McPeek M A, Donoghue M J. Phylogenies and community ecology. Annual Review of Ecology and Systematics, 2002, 33: 475–505. DOI:10.1146/annurev.ecolsys.33.010802.150448 |
| [14] | Jetz W, Fine P V A. Global gradients in vertebrate diversity predicted by historical area-productivity dynamics and contemporary environment. PLoS Biology, 2012, 10(3): e1001292. DOI:10.1371/journal.pbio.1001292 |
| [15] | 刘翥, 杨玉盛, 朱锦懋, 谢锦升, 司友涛. 中亚热带森林转换对土壤可溶性有机质数量与光谱学特征的影响. 生态学报, 2015, 35(19): 6288–6297. |
| [16] | 广西大百科全书编纂委员会. 广西大百科全书. 北京: 中国大百科全书出版社, 2008. |
| [17] | 曹建华, 袁道先, 潘根兴. 岩溶生态系统中的土壤. 地球科学进展, 2003, 18(1): 37–44. DOI:10.3321/j.issn:1001-8166.2003.01.006 |
| [18] | 刘方, 王世杰, 罗海波, 刘元生, 刘鸿雁. 喀斯特森林生态系统的小生境及其土壤异质性. 土壤学报, 2008, 45(6): 1055–1062. DOI:10.3321/j.issn:0564-3929.2008.06.007 |
| [19] | 周游游, 黎树式, 黄天放. 我国喀斯特森林生态系统的特征及其保护利用——以西南地区茂兰、木论、弄岗典型喀斯特森林区为例. 广西师范学院学报(自然科学版), 2003(3): 1–7. DOI:10.3969/j.issn.1002-8743.2003.03.001 |
| [20] | 郭柯, 刘长成, 董鸣. 我国西南喀斯特植物生态适应性与石漠化治理. 植物生态学报, 2011, 35(10): 991–999. |
| [21] | Moritz C. Strategies to protect biological diversity and the evolutionary processes that sustain it. Systematic Biology, 2002, 51(2): 238–254. DOI:10.1080/10635150252899752 |
| [22] | 杜虎, 宋同清, 曾馥平, 王克林, 彭晚霞, 付威波, 李莎莎. 喀斯特峰丛洼地不同植被类型碳格局变化及影响因子. 生态学报, 2015, 35(14): 4658–4667. |
| [23] | Zhang Z H, Hu G, Zhu J D, Luo D H, Ni J. Spatial patterns and interspecific associations of dominant tree species in two old-growth karst forests, SW China. Ecological Research, 2010, 25(6): 1151–1160. DOI:10.1007/s11284-010-0740-0 |
| [24] | Xiang W S, Ding T, Lü S H, Li X K. A comparison of tree species diversity in two subtropical forests, Guangxi, Southwest China. Journal of Resources and Ecology, 2015, 6(4): 208–216. DOI:10.5814/j.issn.1674-764X.2015.04.003 |
| [25] | 王献溥, 郭柯, 温远光. 《广西植被志要》(上、下册). 生命世界, 2015(5): 2–2. |
| [26] | 张新时. 西藏阿里植物群落的间接梯度分析、数量分类与环境解释. 植物生态学与地植物学学报, 1991, 15(2): 101–113. |
| [27] | Gleason H A. The individualistic concept of species diversity.A critique and alternative parameters. Ecology, 1926, 52: 577–586. |
| [28] | Faith D P. Conservation evaluation and phylogenetic diversity. Biological Conservation, 1992, 61(1): 1–10. |
| [29] | Webb C O, Donoghue M J. Phylomatic:tree assembly for applied phylogenetics. Molecular Ecology Notes, 2005, 5(1): 181–183. DOI:10.1111/men.2005.5.issue-1 |
| [30] | Zanne A E, Tank D C, Cornwell W K, Eastman J M, Smith S A, FitzJohn R G, McGlinn D J, O'Meara B C, Moles A T, Reich P B, Royer D L, Soltis D E, Stevens P F, Westoby M, Wright I J, Aarssen L, Bertin R I, Calaminus A, Govaerts R, Hemmings F, Leishman M R, Oleksyn J, Soltis P S, Swenson N G, Warman L, Beaulieu J M. Corrigendum:three keys to the radiation of angiosperms into freezing environments. Nature, 2014, 514(7522): 394. |
| [31] | Swenson N G, Enquist B J, Pither J, Thompson J, Zimmerman J K. The problem and promise of scale dependency in community phylogenetics. Ecology, 2006, 87(10): 2418–2424. DOI:10.1890/0012-9658(2006)87[2418:TPAPOS]2.0.CO;2 |
| [32] | 何艳华, 闫明, 张钦弟, 张直峰, 苗艳明, 毕润成. 五鹿山国家级自然保护区物种多样性海拔格局. 生态学报, 2013, 33(8): 2452–2462. |
| [33] | 朱彪, 陈安平, 刘增力, 李光照, 方精云. 广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局. 生物多样性, 2004, 12(1): 44–52. DOI:10.3321/j.issn:1005-0094.2004.01.006 |
| [34] | Kreft H, Jetz W. Global patterns and determinants of vascular plant diversity. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(14): 5925–5930. DOI:10.1073/pnas.0608361104 |
| [35] | Mo X X, Shi L L, Zhang Y J, Zhu H, Slik J W F. Change in phylogenetic community structure during succession of traditionally managed tropical rainforest in Southwest China. PLoS One, 2013, 8(7): e71464. DOI:10.1371/journal.pone.0071464 |
| [36] | 牛跃, 刘洋, 张一平, 高进波. 哀牢山植物多样性垂直分布格局. 西部林业科学, 2008, 37(4): 27–34. DOI:10.3969/j.issn.1672-8246.2008.04.005 |
| [37] | Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220–227. DOI:10.1038/35012228 |