 2018, Vol. 38
2018, Vol. 38文章信息
- 乔佳伦, 孙太福, 齐利平, 蔡永华, 郑程莉, 程建国, 付文龙, 孟秀祥.
- QIAO Jialun, SUN Taifu, QI Liping, CAI Yonghua, ZHENG Chengli, CHENG Jianguo, FU Wenlong, MENG Xiuxiang.
- 圈养林麝(Moschus berezovskii)个性及与麝香分泌和繁殖成效的关系
- Studies on the personality and relationships of musk secretion and reproduction success in captive Forest musk deer
- 生态学报. 2018, 38(23): 8306-8313
- Acta Ecologica Sinica. 2018, 38(23): 8306-8313
- http://dx.doi.org/10.5846/stxb201805201110
-
文章历史
- 收稿日期: 2018-05-20
- 修订日期: 2018-09-07





2. 中国人民大学环境学院, 北京 100872;
3. 四川养麝研究所, 都江堰 611830
2. School of Environment and Natural Resources, Renmin University of China, Beijing 100872, China;
3. Sichuan Institute of Musk Deer Breeding, Dujiangyan 611830, China
麝类动物(Moschus spp)是珍稀资源动物[1]。因生境丧失、过度利用等原因, 野生麝已极度濒危, 在保育过程中, 出现了怀孕率、成活率低、流产率高等问题, 导致麝的繁殖性能逐年下降, 种群增长速度缓慢[2]。行为研究是迁地保育的重要基础, 目前为止, 学者们基于行为对麝香分泌和繁殖成效进行了多方面的研究, 如母幼接触对幼仔成活率的影响[3], 泌香与种群增长和种群结构的关系、年龄组对泌香的影响[4], 林麝(Moschus berezovskii)的社会行为、饲养方式等对后代的影响[5-6]。此外, 孟秀祥等[7]采用行为量化参数对圈养马麝(Moschus sifanicus)的繁殖性能进行了判别。
近年来, 动物个性已经成为一个新的研究热点, 涉及多个物种类群与学科, 其研究具有重要的生态和进化意义, 为探究次优行为、物种入侵、生活史权衡等提供了新的视角[8-9]。但至今国内外均无从动物个性的角度来探究林麝泌香与繁殖的报道。动物个性是个体表现出的跨时间和情境的稳定行为趋势[10-12], 大量研究表明, 动物个性与其繁殖成效、生活史策略等相关联。如, 探索性强的一种水石龙子(Eulamprus heatwolei)的子代具有较高的平均体重[13], 侵略性强的斑马鱼(Barchydanio reriovar)具有较高的交配成功率[14], 胆大的三刺鱼(Gasterosteus aculeatus)能获得更适宜的繁殖位点[15]。个性作为贯穿动物生活史的统一体, 更关注于个体存在的稳定性差异, 为探究和解决林麝泌香、分娩等问题提供了新的视角[16], 对动物的迁地保育、福利评价等具有重要的参考价值。因此本研究意在建立圈养林麝的个性维度, 并探究个性维度与繁殖成效(如产仔量、仔麝死亡率等)和麝香分泌之间存在的内在关联, 为进一步探明林麝生物学和生态学规律奠定基础, 为迁地保育和饲养管理提供参考。
1 研究方法 1.1 研究地区与对象本研究于四川养麝研究所下属的马尔康林麝繁育场(以下简称麝场)进行。马尔康麝场位于川西北阿坝州, 地处高原, 空气稀薄, 辐射强烈, 年日照时数2214 h, 日照率50%, 气候高寒, 年均温为5—9℃, 昼夜温差大。年均降雨760 mm, 多夜雨。马尔康的森林资源十分丰富, 是国家重要木材生产基地; 动物资源也十分丰富, 大型草食性动物主要有羚牛(Budorcas taxicolor)、藏原羚(Procapra picticaudata)等[17]。
四川马尔康麝场现有9个饲养区, 每个饲养区中由5—8个并排圈舍组成, 每个圈舍由4—7个小舍(每个小舍面积2—4 m2, 混凝土地面基底)及一活动场(面积100 m2)组成, 活动场中央位置是水泥凉棚(6 m2左右), 凉棚四周设置2个饮水槽和2个叶槽。活动场为硬化的泥质基底, 有天然杂草植被和榆树等落叶乔木。每日2次于相对固定时间(夏季:06:00和19:00;冬季:08:00和17:00)投放树叶、水等辅料。其他时间除打扫圈舍外不再进入圈内, 保证林麝不受外界干扰。
本研究涉及样本172头, 其中雄麝113头, 雌麝59头; 将林麝按年龄划分年龄组为亚成体林麝(1—2岁, 73头)、成体林麝(3—7岁, 68头)和老年林麝(≥8岁, 31头)。
1.2 数据收集本研究参照黇鹿(Dama dama)的个性量表[18]以及云豹(Neofelis nebulosa)的个性分类和定义[19], 结合已报道的麝类动物的行为模式[20], 构建圈养林麝的个性分类, 其定义参见表 1。建立林麝个性评估标尺(标尺为7 cm长, 最小刻度为1 mm), 个性评估人员(熟悉林麝个体的饲养员)在标尺上对林麝的个性得分进行打点, 依据打点在标尺上进行读数、判分, 得分越高则相应个性越强。
| 个性 Personality | 定义 Explanation and definition |
| 稳定性Stability | 林麝在平时行活动中表现稳定, 其行为在圈养环境和其他林麝个体前不会轻易改变 |
| 自我指向Self-pointing | 林麝较长时间地表达求适及刻板等发出动物和指向动物均为个体本身的行为, 行为比较单一、重复, 而且不易停止 |
| 鲁莽性Recklessness | 林麝相对轻率, 不关心自身行为的后果 |
| 社会性(合群性)Sociality | 林麝在圈养活动中更加倾向于对其他个体展现友好行为, 进行交互活动, 独处时间较少 |
| 社会性胆小Social timidity | 动物显得比较缺乏自信、易于被恐吓, 在新的社群环境中不积极、害怕 |
| 非社会性胆小 Non-social timidity |
动物显得比较缺乏自信、易于被恐吓, 在新的圈舍等物理环境中不积极 |
| 易受伤害性Vulnerability | 林麝容易因自身不当的行为或者受到其他个体的攻击而导致的可能是生理上或者心理上的伤害 |
| 欺凌弱小Bullying | 容易对年龄较小、身体较弱或新进入社群的个体释放侵犯行为 |
| 侵略性Aggressiveness | 在日常活动中, 林麝容易对其他个体展现示警、喷气、攻击、取代等攻击性行为 |
| 友善性Friendliness | 林麝与其他个体社会关系密切, 相互间舔理等亲和行为较多, 会展现共同摄食等行为 |
| 冲动性Impulsiveness | 没有任何预兆的情况下林麝展现突然的行为, 持续时间短, 行为表达比较难以预测 |
| 镇静性Calm | 林麝较少受情绪影响, 显得镇定、自信和平静 |
| 暴躁Irritability | 较多时间林麝处于不好的状态, 面对外界环境, 容易由平静的状态产生愤怒的情绪 |
| 挑衅性Impulse | 林麝对高序位等级的个体发动挑衅性行为, 并伴随其率先释放的行为, 受到这些个体的威胁 |
| 保护性Protection | 个体花费较多时间关注社群及共同相处的其他个体, 会较多释放警戒行为减少同群个体受伤害的可能性 |
| 反复无常Unsteady | 个体的行为或状态不稳定、展现重复、不连贯、不确定的的行为, 如往返走 |
| 同人友好性Human friendliness | 经常寻求同人的亲和接触, 很少对人主动释放侵犯行为 |
采用Cronbach′s α系数和因子分析对量表进行信度和效度分析, 以验证量表设计合理性。结合因子分析与聚类分析对全部个性项目进行降维, 建立林麝的个性维度, 并采用Pearson相关系数探究圈养林麝个性维度与麝香分泌和繁殖成效的关系。
个性维度表达式为:
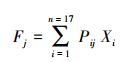
|
式中, Fj为第j个个性维度, Xi为第i个个性项目的评分, Pi为第i个个性项目的因子载荷值。本研究所有数据处理均在Excel 2010与SPSS 20中完成。
2 结果 2.1 马尔康圈养林麝个性维度对量表进行信度分析, Cronbach′s α系数为0.648, 说明量表内部各项目一致性较高。进行KMO和Bartlett检验, Bartlett球度检验的P为0.000, KMO值为0.838, 根据KMO度量标准可知, 量表的17个个性项目适合进行因子分析。因子分析结果显示, 当特征根大于1时, 有5个主成分, 能够解释70.77%的个性项目, 各项目载荷均在0.4以上(表 2), 表明量表具有良好的结构效度, 可以比较全面的反映整体。
| 序号 Number |
构成变量 Variance |
因子载荷Factor loadings | 得分均值 Scores mean |
||||
| 侵略性 Aggressiveness |
活跃性 Activity |
刻板性 Stereotype |
胆怯型 Timidity |
社会性 Sociability |
|||
| 1 | 同人友好 Human friendliness |
-0.810 | -0.010 | -0.179 | -0.179 | 0.097 | 4.36±0.28 |
| 2 | 侵略性Aggressiveness | 0.685 | 0.520 | 0.162 | -0.184 | -0.031 | 3.59±0.12 |
| 3 | 欺凌弱小Bullying | 0.670 | 0.441 | 0.235 | -0.106 | -0.014 | 3.60±0.12 |
| 4 | 暴躁Irritability | 0.670 | 0.565 | 0.214 | -0.045 | -0.054 | 3.60±0.12 |
| 5 | 易受伤害的 Vulnerability |
0.607 | -0.006 | 0.476 | -0.130 | 0.047 | 3.08±0.12 |
| 6 | 挑衅性Impulse | 0.135 | 0.842 | 0.134 | -0.008 | 0.011 | 3.81±0.11 |
| 7 | 冲动性Impulsiveness | 0.385 | 0.649 | 0.301 | 0.022 | -0.074 | 3.76±0.12 |
| 8 | 鲁莽性Recklessness | 0.019 | 0.613 | 0.402 | 0.308 | 0.032 | 4.02±0.11 |
| 9 | 自我指向Self-pointing | 0.282 | 0.192 | 0.746 | -0.007 | 0.067 | 3.43±0.13 |
| 10 | 反复无常Unsteady | 0.243 | 0.282 | 0.711 | -0.168 | 0.028 | 3.06±0.11 |
| 11 | 镇静性Calm | -0.125 | -0.438 | -0.642 | -0.082 | 0.163 | 4.14±0.20 |
| 12 | 稳定性Stability | -0.207 | -0.079 | -0.640 | 0.001 | 0.449 | 4.21±0.12 |
| 13 | 非社会性胆小 Non-social timidity |
0.000 | 0.003 | -0.016 | 0.939 | -0.021 | 4.31±0.14 |
| 14 | 社会性胆小 Social timidity |
-0.049 | 0.073 | -0.069 | 0.928 | 0.084 | 3.85±0.15 |
| 15 | 友善性Friendliness | -0.158 | -0.031 | 0.050 | 0.021 | 0.871 | 4.21±0.12 |
| 16 | 保护性Protection | -0.007 | 0.148 | -0.118 | -0.130 | 0.730 | 3.95±0.25 |
| 17 | 社会性(合群)Sociality | 0.090 | -0.154 | -0.029 | 0.180 | 0.671 | 4.31±0.14 |
| 特征根Eigen value | 5.871 | 2.089 | 1.922 | 1.100 | 1.049 | ||
| 方差贡献率 Variance/% |
34.538 | 12.288 | 11.306 | 6.470 | 6.169 | ||
| 累计贡献率/% Total variance |
34.538 | 46.826 | 58.132 | 64.602 | 70.771 | ||
| 数据为平均值±标准误 | |||||||
依据因子分析结果, 将林麝个性分为5个维度:第一主成分包含侵略性, 欺凌弱小等项目, 反映了林麝对其他个体的侵犯性行为的释放强度, 命名为“侵犯性”维度。第二成分中包括冲动, 鲁莽等, 命名为“活跃性”维度。第三主成分包括自我指向、反复无常等, 反映了林麝行为的单一性和刻板性, 命名为“刻板性”维度。第四主成分中包括非社会性胆小、社会性胆小, 命名为“胆怯性”维度。第五主成分包含友善性, 保护性等项目, 反映了林麝在群体中的社会关系, 命名为“社会性”维度。其表达式分别为:

|
利用聚类分析对17个个性项目进行聚类, 为与主成分分析结果相比较, 在10.00处进行切分, 可将全部个性项目划分为五大类(图 1), 而其中胆怯性、社会性、活跃性三大类别与主成分分析的结果完全一致, 其他个性稍有区别, 但整体结果基本一致, 进一步验证了5大个性维度划分的合理性。

|
| 图 1 圈养林麝个性评分的聚类分析 Fig. 1 Clustering analysis of captive Forest musk deer′s personalities |
如表 3所示, 社会性维度和雄性产香量存在显著正相关(r=0.234, P= 0.023 < 0.05, N=95)。与活跃性、刻板性、胆怯性维度相比, 侵略性维度和产香量之间有较强的负相关关系, 但结果不显著(r=-0.186, P=0.071>0.05, N=95)。
| 相关性 Correlation |
侵略性 Aggressiveness |
活跃性 Activity |
刻板性 Stereotype |
胆怯性 Timidity | 社会性 Sociability |
|
| 产香Musk secretion | r | -0.186 | -0.037 | -0.095 | -0.040 | 0.234* |
| P | 0.071 | 0.723 | 0.362 | 0.704 | 0.023 | |
| N | 95 | 95 | 95 | 95 | 95 | |
| *:P < 0.05;**:P < 0.01 | ||||||
如表 4所示, 对雌性林麝的分娩时间和五大个性维度进行Pearson相关分析, 结果显示侵略性(r=0.463, P=0.003, N=39)、活跃性(r=0.440, P=0.005, N=39)、刻板性(r=0.595, P=0.000, N=39)与分娩时间存在显著正相关关系, 即个性越强, 分娩时间越晚。胆怯性和社会性维度相关性均不显著。
| 相关性 Correlation |
侵略性 Aggressiveness |
活跃性 Activity |
刻板性 Stereotype |
胆怯性 Timidity | 社会性 Sociability |
|
| 分娩时间 | r | 0.463** | 0.440** | 0.595** | 0.050 | -0.053 |
| Delivery time | P | 0.003 | 0.005 | 0.000 | 0.762 | 0.750 |
| N | 39 | 39 | 39 | 39 | 39 | |
| *:P < 0.05;**:P < 0.01 | ||||||
相关性分析结果显示, 雌麝的个性“刻板性”与产仔数呈显著负相关(r=-0.341, P=0.034 < 0.05), 侵犯性与产仔数呈正相关, 结果不显著(r=0.063, P=0.704>0.05)。而活跃性(r=-0.076, P=0.646>0.05)、胆怯性(r=-0.054, P=0.744>0.05)和社会性维度(r=-0.119, P=0.471>0.05)与产仔数呈负相关, 但不显著。
五大个性维度与仔麝死亡数均无显著相关性。侵犯性与产仔死亡数呈正相关, 结果不显著(r=0.063, P=0.704>0.05)。活跃性(r=-0.035, P=0.834>0.05)、刻板性(r=-0.113, P=0.492>0.05)、胆怯性(r=-0.302, P=0.061>0.05)和社会性(r=-0.035, P=0.831>0.05)与仔麝死亡数呈负相关, 均不显著。
3 讨论 3.1 动物个性研究的方法论行为编码法(The coding technique)是动物个性研究最常用的方法, 它包括基于情景的自然观察法和试验测评法[21-22]。此外, 当研究对象为动物园或圈养个体时, 基于量表的个性评价法在动物个性研究方面也具有重要实践价值[23]。
本研究对象为圈养林麝, 由于圈养条件和物种自身特点的限制, 难以进行新奇物、镜子测试等行为实验。本研究采用了个性的量表评价法, 以饲养员为获取数据的对象, 进行林麝个性测评。参考前人研究, 编制出适合圈养林麝的含有17个项目的个性量表。并根据饲养员对个性项目的打分计测数据, 采用统计学方法进行信度和效度分析, 以验证量表设计的合理性。最后, 通过因子分析和聚类分析来进行个性维度划分。
此类研究方法体系可以在较短的时间内获得一组具有长时期代表性的数据(评价者对动物个体熟悉, 具有丰富的经验), 可为其他动物个性研究提供重要参考。需要注意的是个性评价法主观性较大, 要求一个大的样本量且评价者需对动物个体较为熟悉。
3.2 圈养林麝的个性维度本研究采用个性评价法, 通过具有丰富管理经验的饲养员来获取数据, 保证了数据的可靠性。对所有项目进行内部信度分析, Cronbach′s α系数为0.648, 此外, 因子分析结果显示, 各项目载荷均在0.4以上, 表明量表具有良好的结构效度[24]。当特征根大于1时, 提取5个主成分, 能够解释70.77%的个性项目, 因此将圈养林麝17个项目划分为5大个性维度:侵犯性, 活跃性, 刻板性, 胆怯性和社会性。通过聚类分析, 进一步验证了个性维度划分的合理性。
大部分关于林麝泌香、产仔等研究, 均侧重于分析性别、年龄、体重等离散变量的差异, 而动物个性作为贯穿个体生活史的统一体, 体现了个体之间存在的稳定性差异, 为林麝的相关研究提供了新的视角。研究显示, 林麝的侵犯性维度贡献率最高, 一定程度上反应了林麝群体间的冲突情况。一个重要原因是圈养条件的限制, 如范围限制、统一喂食等。野外林麝独居性极强, 领域较大, 而在圈养条件下, 被迫与其他个体进行食物、空间等资源竞争, 因此更易展现威胁、取代、逃离等侵犯性行为。有研究报道, 不同种类以及同种动物在不同环境下都存在个性维度的差异, 本研究发现圈养林麝展现出刻板性维度, 是由于其不能尽快适应所处的非自然环境, 更不能有效的选择和改变环境以及环境因子的类型和作用方式, 其行为模式经常被环境刺激及相关的学习过程所修饰, 而导致动物行为中出现了食异物、狂奔、往返走等行为, 展现了刻板性[25]。
本研究建立的圈养林麝的个性量表与维度, 具有重要的实践价值。如:圈养动物刻板行为的发育和表达对动物的繁殖成效及生产力均会施加负面影响[26], 本研究可以帮助饲养员进行科学的主观评价, 评估动物的刻板程度, 发现刻板行为发育的规律特点, 以便更及时的对圈养林麝采取有效管理措施; 对林麝个体的侵略性进行判别, 指导圈舍的合理分配, 避免个别侵犯性强的个体对其他林麝造成伤害。对胆怯性个体进行分圈、治疗和取香等过程中, 采用适当的方式以避免对林麝造成伤害, 防止成年林麝狂奔误撞仔麝。
3.3 雄性林麝个性维度与泌香的关系本研究进行了个性维度与泌香量的相关性分析, 发现社会性维度和产香量存在显著正相关(r=0.234, P=0.023 < 0.05, N=95)。该维度包含友善性、保护性等项目, 反映了与其他个体的接触程度, 社会性高的个会展现出更多的亲和性接触、互饰、共同摄食等亲和行为。麝香是雄麝特有的麝香腺囊分泌物, 是对麝群行使化学通讯机能的一种信息素[27], 促进麝香分泌的激素为雄性激素, 且泌香盛期结束即为交配期。本研究结果表明, 泌香量越高, 其社会性越强, 因此本研究推测泌香增加了同其他个体的信息交流与接触, 可能对于求偶交配过程具有一定作用, 意味着个体的交配成功率与社会性强度有密切关系, 需要进一步探究。
3.4 雌性林麝个性维度与繁殖成效的关系为探究个性维度与林麝繁殖成效的关系, 本研究从分娩时间、产仔数、仔麝死亡数3个方面进行探索分析。其中分娩时间格局对仔麝成活率有显著影响, 分娩时间越晚的个体其仔麝成活率较低[28]。孙军平等[29]探究过年龄等生理因素和圈舍条件等环境因素对圈养林麝分娩定时的效应, 而本研究从个性角度进行探究, 发现侵略性、活跃性、刻板性均与分娩时间存在显著正相关关系, 即个性越强, 分娩时间越晚。一个重要原因为动物个体的体质状况和能量储备是有限的, 分娩需要投资一定的时间和能量, 而侵犯性、活跃性、刻板性越强, 意味着其投入分娩的能量相对较低, 因此导致其繁殖成效下降。另外, 研究发现产仔数与刻板性呈显著负相关, 进一步表明刻板性强的个体对繁殖成效会有一个负面的效应, 这与前人研究结果一致[30]。圈养动物刻板行为的发育和表达对动物的繁殖成效及生产力均会施加负面效应, 侵犯性和活跃性虽然与分娩定时相关, 但却与产仔数无显著相关性, 是因为侵略性、活跃性强的个体可能将能量投资于争夺摄食序位、空间等资源, 牺牲了最佳产仔时间, 却为产仔做好充足准备, 这体现了动物个体繁殖策略的选择与权衡, 在某一情境中虽表现出次优行为, 但相对于动物整个生活史而言, 可能是一种最优选择。圈养动物的刻板行为是在胁迫环境下的表达, 其模式固定且反复, 无明显行为目的和功能, 因此会影响动物个体合理的时间和能量分配。另外, 本研究结果显示五大个性维度与仔麝死亡数均无显著相关性, 一个重要原因可能是圈养环境下仔麝死亡多与疾病相关, 这可能与圈养环境、气候等相关性较大, 而与个体本身的个性维度相关性较小。
致谢: 感谢四川养麝研究所所属马尔康林麝繁育中心对本研究的支持。| [1] | Meng X X, Feng J C, Yun M Y, Wang B, Cody N. Relationships between musk extraction, social rank and tail-rubbing in male Alpine musk deer, Moschus sifanicus. Biologia, 2011, 66(5): 928–932. |
| [2] | Zhou Y J, Meng X X, Feng J C, Yang Q S, Feng Z J, Xia L, Bartoš L. Review of the distribution, status and conservation of musk deer in China. Folia Zoologica, 2004, 53(2): 129–140. |
| [3] | 杜卫国, 盛和林. 林麝哺乳期的时间分配和行为研究. 兽类学报, 1998, 18(1): 22–27. |
| [4] | 康发功, 盛岩, 马泠桃, 孟秀祥. 兴隆山自然保护区驯养马麝的麝香分泌及与其种群动态和年龄结构的关系. 生态学报, 2015, 35(15): 4993–4999. |
| [5] | 徐正强, 徐宏发. 圈养雌麝社会行为及其对后代的影响. 兽类学报, 2002, 22(2): 155-158, 86-86. DOI:10.3969/j.issn.1000-1050.2002.02.010 |
| [6] | 徐正强, 徐宏发. 饲养林麝的种群特征和幼麝的存活研究. 兽类学报, 2003, 23(1): 17–20. DOI:10.3969/j.issn.1000-1050.2003.01.004 |
| [7] | 孟秀祥, 杨奇森, 冯祚建, 冯金朝, 周宜君, 徐宏发. 甘肃兴隆山保护区圈养雄性马麝繁殖性能的行为判别. 动物学研究, 2007, 28(2): 149–154. DOI:10.3321/j.issn:0254-5853.2007.02.006 |
| [8] | Sih A, Bell A, Johnson J C. Behavioral syndromes:an ecological and evolutionary overview. Trends in Ecology & Evolution, 2004, 19(7): 372–378. |
| [9] | Bell A M. Animal personalities. Nature, 2007, 447(7144): 539–540. |
| [10] | Caspi A, Roberts B W, Shiner R L. Personality development:stability and change. Annual Review of Psychology, 2005, 56: 453–484. DOI:10.1146/annurev.psych.55.090902.141913 |
| [11] | Stamps J, Groothuis T G G. The development of animal personality:relevance, concepts and perspectives. Biological Reviews, 2010, 85(2): 301–325. DOI:10.1111/brv.2010.85.issue-2 |
| [12] | Carere C, Locurto C. Interaction between animal personality and animal cognition. Current Zoology, 2011, 57(4): 491–498. DOI:10.1093/czoolo/57.4.491 |
| [13] | Stapley J, Keogh J S. Behavioral syndromes influence mating systems: floater pairs of a lizard have heavier offspring. Behavioral Ecology, 2005, 16(3): 514-520. |
| [14] | Ariyomo T O, Watt P J. The effect of variation in boldness and aggressiveness on the reproductive success of zebrafish. Animal Behaviour, 2012, 83(1): 41-46. |
| [15] | Huntingford F A. The relationship between anti-predator behaviour and aggression among conspecifics in the three-spined stickleback, Gasterosteus Aculeatus. Animal Behaviour, 1976, 24(2): 245-260. |
| [16] | Wilson D S, Clark A B, Coleman K, Dearstyne T. Shyness and boldness in humans and other animals. Trends in Ecology & Evolution, 1994, 9(11): 442–446. |
| [17] | 蔡永华, 孙军平, 杨营, 王静, 付文龙, 朱萍, 郑程莉, 李宁, 石磊, 孟秀祥. 迁地保育林麝体况及影响因素. 生态学报, 2017, 37(5): 1617–1622. |
| [18] | Bergvall U A, Schäpers A, Kjellander P, Weiss A. Personality and foraging decisions in fallow deer, Dama dama. Animal Behaviour, 2011, 81(1): 101–112. |
| [19] | Wielebnowski N C. Behavioral differences as predictors of breeding status in captive cheetahs. Zoo Biology, 1999, 18(4): 335–349. DOI:10.1002/(SICI)1098-2361(1999)18:4<>1.0.CO;2-# |
| [20] | 孟秀祥, 冯金朝, 周宜君, 杨奇森, 冯祚建, 薛程, 徐宏发. 麝类行为谱的初步建构及行为型的描述性定义. 四川动物, 2007, 26(1): 46–50. DOI:10.3969/j.issn.1000-7083.2007.01.011 |
| [21] | Gosling S D. From mice to men:what can we learn about personality from animal research?. Psychological Bulletin, 2001, 127(1): 45–86. DOI:10.1037/0033-2909.127.1.45 |
| [22] | Watters J V, Powell D M. Measuring animal personality for use in population management in zoos:suggested methods and rationale. Zoo Biology, 2012, 31(1): 1–12. DOI:10.1002/zoo.20379 |
| [23] | Freeman E W, Schulte B A, Brown J L. Using behavioral observations and keeper questionnaires to assess social relationships among captive female African elephants. Zoo Biology, 2010, 29(2): 140–153. |
| [24] | 李灿, 辛玲. 调查问卷的信度与效度的评价方法研究. 中国卫生统计, 2008, 25(5): 541–544. DOI:10.3969/j.issn.1002-3674.2008.05.034 |
| [25] | 孟秀祥, 杨奇森, 冯祚建, 徐宏发, 冯金朝, 周宜君. 圈养马麝的刻板行为. 东北林业大学学报, 2007, 35(1): 47-48, 57-57. DOI:10.3969/j.issn.1000-5382.2007.01.016 |
| [26] | 孟秀祥, 贡保草, 薛达元, 冯金朝, 杨奇森, 冯祚建. 圈养马麝刻板行为表达频次及影响因素. 生态学报, 2011, 31(22): 6896–6901. |
| [27] | 毕书增, 张治国, 贾林征, 关强, 宋建民. 麝泌香盛期后麝香腺囊的显微与超微结构和麝香分泌研究. 兽类学报, 1987, 7(2): 96–99. |
| [28] | 洪沂生, 李复东, 邓文明, 李时万. 麝泌香与雄性激素的关系. 中药材科技, 1981(1): 19–22. |
| [29] | 孙军平, 蔡永华, 杨营, 王静, 付文龙, 程建国, 孟秀祥. 驯养林麝(Moschus berezovskii)的繁殖节律:分娩定时及同步化. 生态学报, 2017, 37(5): 1611–1616. |
| [30] | 孟秀祥, 杨奇森, 冯祚建, 施哲梅, 蒋应文. 马麝(Moschus sifanicus)的分娩时间格局. 四川动物, 2005, 24(1): 30–35. DOI:10.3969/j.issn.1000-7083.2005.01.008 |