 2018, Vol. 38
2018, Vol. 38文章信息
- 罗云建, 张小全, 朱建华, 张治军, 车通.
- LUO Yunjian, ZHANG Xiaoquan, ZHU Jianhua, ZHANG Zhijun, CHE Tong.
- 关帝山林区退化灌木林转变为华北落叶松林对生态系统碳储量的影响
- Changes in ecosystem carbon storage following conversion of degraded shrubland to larch (Larix principis-rupprechtii Mayr.) in the Guandi Mountain Forest Region, northern China
- 生态学报. 2018, 38(23): 8354-8362
- Acta Ecologica Sinica. 2018, 38(23): 8354-8362
- http://dx.doi.org/10.5846/stxb201801250204
-
文章历史
- 收稿日期: 2018-01-25
- 网络出版日期: 2018-09-26





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;
3. 大自然保护协会中国部, 北京 100600;
4. 中国林业科学研究院森林生态环境与保护研究所, 北京 100091;
5. 国家林业局昆明勘察设计院, 昆明 650216
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. The Nature Conservancy China Program, Beijing 100600, China;
4. Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China;
5. Kunming Institute of Forest Exploration and Design, State Forestry Administration, Kunming 650216, China
灌木林是一种以灌木为主体的植被类型, 具有生命力强、区系组成复杂、生活型多样、生态适应性广等特点, 其在群落演替、生物固碳等方面扮演着不可替代的角色[1]。作为世界上灌木林分布最广泛的国家之一, 我国灌木林面积达1.8×108 hm2, 约占全国陆地面积的18.5%[2]。由于自然和人为因素的影响, 大面积的灌木林表现出郁闭度低、群落结构趋于简单、生态功能(如涵养水源、保持水土)降低等一系列退化特征[3]。随着我国多项林业生态工程(如京津风沙源治理工程、三北及长江流域等重点防护林体系建设工程)的逐步开展, 宜林地大幅减少, 自然条件更加严峻, 造林绿化推进的难度也越来越大。因此, 在适宜种植乔木的地区(如年均降水量大于400—500 mm[4]), 将退化灌木林转变为乔木林被认为是一种可行的植被恢复方式[3, 5-7]。
目前, 人们认为造林/再造林、森林管理、减少毁林等林业活动是减少温室气体排放、稳定大气CO2浓度的有效措施, 其中造林/再造林是最为重要的一种[8]。关于造林/再造林的固碳效应, 大量的案例研究和整合分析(meta-analysis)表明, 固碳效应由前期土地利用方式、气候、土壤类型、林龄、树种、管理措施等因素共同控制[9-13]。但也存在一些不足:①侧重非林地(如耕地、草地、裸地)转变为林地、天然乔木林转变为人工林等的固碳效应, 相对缺少对灌木林转变为乔木林的探讨;②主要关注森林的某一碳库, 特别是土壤碳库, 很少从整个生态系统开展;③以前期土地利用方式为对照, 常研究造林/再造林之后某特定时间的固碳功能, 而对固碳功能的动态了解不多。
关帝山林区是华北山地暖温带植被的典型代表, 拥有保存良好的寒温性针叶林、温性针叶林、温性阔叶林、温性灌木林、温性草丛等植被。由于频繁的人为干扰和破坏, 较大面积的灌木林次生林出现了生长缓慢、群落结构失调、生态功能退化等问题。华北落叶松(Larix principis-rupprechtii Mayr.)是我国暖温带亚高山森林类型的主要构成树种, 也是华北亚高山地区、黄河流域山区、辽河上游山区等地区林分更新和荒山造林的重要树种[14]。当地人们为提升森林的生态功能(如涵养水源、保持水土), 将一部分退化灌木次生林转变为华北落叶松林。现有研究主要关注华北落叶松林自身的生物量、生产力、元素循环、碳储量等方面, 尚不清楚这种转变的固碳效应如何。因此, 本文选取由退化灌木次生林转变而成的不同林龄华北落叶松林, 并以相邻的退化灌木次生林为对照, 探究这种转变对生态系统碳储量及其组分的影响, 为我国开展造林/再造林、林业碳汇项目等工作提供科学依据和数据支撑。
1 材料与方法 1.1 研究区概况关帝山林区(111°18′—112°18′ E, 37°20′—38°20′ N), 位于山西西部的吕梁山脉中段, 为方山、娄烦两县之界山, 主峰孝文山海拔2831 m, 相对高差约2000 m, 平均坡度25°左右。地处暖温带大陆性季风气候区, 年均气温3—4℃, 年均降水量600—800 mm, 无霜期100—130 d。成土母岩以花岗岩、片麻岩、石英岩和角闪岩为主;土壤属于山地土壤类型, 从低到高垂直带谱为山地褐土、山地淋溶褐土、山地棕壤和亚高山草甸土。
1.2 研究方法根据造林记录档案及实地踏查, 筛选出立地条件(海拔、坡度和坡向)、造林方式(如林地清理、造林密度)、管理措施(如修剪、施肥)等方面基本一致但不同造林时间的地段, 以期尽量降低非林龄因素的影响。于2006年生长季(7—8月), 选择不同林龄(10、18、23、27年和35年, 涵盖幼龄、中龄和近熟3个生长阶段)的华北落叶松人工林以及立地条件基本一致、相邻的退化灌木林, 共设置25块20 m × 20 m的调查样地(表 1)。各林龄华北落叶松林均是由退化灌木次生林转变而来;退化灌木次生林的优势种主要为虎榛子(Ostryopsis davidiana Decaisne)、黄刺玫(Rosa xanthina Lindl.)、三裂绣线菊(Spiraea trilobata L.)、沙棘(Hippophae rhamnoides L.)等。
| 基本情况 Characteristics |
林龄Stand age/a | |||||
| CK | 10 | 18 | 23 | 27 | 35 | |
| 样地数Plot number | 7 | 3 | 4 | 4 | 4 | 3 |
| 海拔Altitude/m | 1921±39 | 1902±13 | 1908±140 | 1935±146 | 1915±25 | 1943±79 |
| 坡度Slope/(°) | 30±2 | 28±3 | 31±3 | 33±3 | 31±3 | 33±4 |
| 坡向Aspect | N, NE | N | N, NE | N, NE | N, NE | N, NE |
| 郁闭度Canopy density/% | 24±9 | 43±6 | 70±14 | 73±10 | 70±8 | 67±6 |
| 平均胸径Mean DBH/cm | ― | 5.0±0.8 | 10.2±0.4 | 11.8±0.4 | 13.7±0.6 | 17.0±0.3 |
| 平均树高Mean height/m | 1.6±0.8 | 4.5±0.4 | 9.1±0.4 | 10.8±0.6 | 11.3±0.7 | 14.8±0.6 |
| 林分密度Stand density/(株/hm2) | 4100±1695 | 2231±301 | 1833±126 | 1950±197 | 1669±188 | 1583±260 |
| 人类干扰Human disturbance | Int | Ext | Ext | Ext | Ext | Ext |
| *数据以“平均值±标准偏差”呈现;CK表示灌木林, 其为华北落叶松(Larix principis-rupprechtii Mayr.)人工林的对照;N和NE分别表示北坡和东北坡;Int和Ext分别表示强烈干扰和粗放管理 | ||||||
对每块样地进行每木检尺, 测定胸径和树高, 计算平均胸径和树高, 依此选取3—4株标准木。将标准木沿地表伐倒, 地上部分采用Monsi分层切割法, 以每段1 m进行分割;地下部分采用全挖法。野外称取各器官(树干、树枝、树叶和树根)的鲜质量, 同时随机取各器官的样品200 g以上带回实验室, 85℃恒温下烘干至恒重, 测定各器官的含水率和含碳量, 计算器官生物量(即烘干质量)。基于57株标准木的胸径(D, cm)、树高(H, m)、器官生物量(W, kg)等实测数据, 利用两种具有代表性的模型W=a·Db和W=a·(D2H)b[15], 建立器官生物量模型(表 2), 选择拟合效果较优者, 用于估算样地内每株华北落叶松的生物量, 从而推算乔木单位面积的生物量(Mg/hm2)。
| 器官 Tree component |
模型形式 Model form |
a (S.E.) | b (S.E.) | R2 | S.E.E | P-value |
| 树干Stem | W=a·Db | 0.052 (0.008) | 2.489 (0.062) | 0.967 | 0.224 | < 0.001 |
| W=a·(D2H)b | 0.049 (0.007) | 0.854 (0.019) | 0.973 | 0.201 | < 0.001 | |
| 树枝Branch | W=a·Db | 0.215 (0.039) | 1.291 (0.076) | 0.840 | 0.275 | < 0.001 |
| W=a·(D2H)b | 0.210 (0.038) | 0.442 (0.026) | 0.841 | 0.274 | < 0.001 | |
| 树叶Needle | W=a·Db | 0.297 (0.073) | 0.852 (0.103) | 0.553 | 0.373 | < 0.001 |
| W=a·(D2H)b | 0.296 (0.074) | 0.290 (0.036) | 0.547 | 0.376 | < 0.001 | |
| 地上部Aboveground | W=a·Db | 0.244 (0.036) | 1.990 (0.062) | 0.949 | 0.224 | < 0.001 |
| W=a·(D2H)b | 0.234 (0.033) | 0.683 (0.020) | 0.954 | 0.214 | < 0.001 | |
| 树根Root | W=a·Db | 0.031 (0.005) | 2.106 (0.068) | 0.946 | 0.246 | < 0.001 |
| W=a·(D2H)b | 0.030 (0.005) | 0.719 (0.024) | 0.943 | 0.253 | < 0.001 | |
| 整株Whole tree | W=a·Db | 0.277 (0.039) | 2.004 (0.059) | 0.955 | 0.213 | < 0.001 |
| W=a·(D2H)b | 0.266 (0.036) | 0.687 (0.019) | 0.958 | 0.205 | < 0.001 | |
| *标准木的数量为57, 其胸径(D)和树高(H)范围分别为2.8—17.5 cm和2.7—15.3 m;S.E.为标准误;R2为决定系数;S.E.E.为估计值的标准误 | ||||||
在每块样地, 沿对角线设置5个2 m × 2 m的灌木样方和5个1 m × 1 m的草本样方。灌木和草本生物量的测定均采用样方全挖法, 测定地上和地下两部分的鲜质量。凋落物采用样方收获法, 即收集5个草本样方内的凋落物, 称其鲜质量。各部分随机取样品200 g以上带回实验室, 85℃恒温下烘干至恒重, 测定各部分的含水率和含碳量, 计算每个样方灌木、草本和凋落物的生物量, 进而推算它们单位面积的生物量(Mg/hm2)。
土壤测定与取样在草本样方中进行, 挖取土壤剖面, 沿着剖面深度0—10、10—30 cm和30—50 cm进行分层测定与取样。利用100 cm3环刀测定每层单位体积土壤湿重, 同时用铝盒采集土壤样品, 带回实验室在105℃烘干24 h后, 测定每层土壤的含水率, 计算每层土壤的容重。此外, 每层另取约500 g土样带回实验室风干处理, 待土样自然风干后过100目土壤筛, 测定土壤有机碳含量。
植物和凋落物的含碳量均采用元素分析仪Elementar Vario EL(德国)测定;土壤有机碳含量采用重铬酸钾外加热法测定。
1.3 统计分析基于单位面积生物量、器官含碳量、土壤容重、土壤有机碳含量等数据, 计算生态系统碳储量(ECS, Mg/hm2), 公式如下:
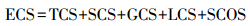
|
(1) |
式中, TCS=∑TBi·TCi;SCS=∑SBi·SCi;GCS=∑GBi·GCi;LCS=LB·LC;SCOS=∑BDi·SOCi·Di。TCS、SCS、GCS、LCS和SCOS分别为乔木、灌木、草本、凋落物和土壤的碳储量(Mg/hm2);TBi和TCi分别为乔木第i器官(树干、树枝、树叶和树根)的单位面积生物量(Mg/hm2)和含碳量(%);SBi和SCi分别为灌木第i器官(地上和地下)的单位面积生物量(Mg/hm2)和含碳量(%);GBi和GCi分别为草本第i器官(地上和地下)的单位面积生物量(Mg/hm2)和含碳量(%);LB和LC分别为凋落物的单位面积生物量(Mg/hm2)和含碳量(%);BDi为第i层土壤容重(g/cm3);SOCi为第i层土壤有机碳含量(%);Di为第i层土壤厚度(cm)。
两组数据间的差异采用t检验进行比较;两组以上数据间的差异采用单因素方差分析进行比较, 若组间差异显著(P < 0.05), Duncan方法用于两两比较。t检验和单因素方差分析均在软件SPSS Statistics 23.0中完成, 制图使用OriginPro 2016。
2 结果 2.1 植物和凋落物的含碳量华北落叶松树干、树枝、树叶和树根的含碳量为(51.3±1.0)%(平均值±标准偏差, 下同)、(51.7±0.5)%、(50.4±0.4)%和(50.0±0.6)%(图 1)。树干和树枝的含碳量均显著高于树叶和树根(P < 0.05), 但树干和树枝之间以及树叶和树根之间的含碳量并无显著差异(P=0.20和P=0.18)。灌木和草本地上器官的含碳量(48.6%和42.2%)都明显高于其地下器官(47.7%和38.6%)(P=0.02和P=0.02)(图 1)。乔木(华北落叶松)的含碳量(50.7%)显著高于灌木(48.2%)、草本(40.4%)和凋落物(41.4%), 但草本和凋落物之间并无显著差异(P=0.14)(图 1)。

|
| 图 1 植物(乔木、灌木和草本)和凋落物的含碳量 Fig. 1 Carbon concentrations of plant(tree, shrub and herb)and litter 图中误差线表示标准偏差, 不同小写英文字母表示组间差异显著(P < 0.05) |
灌木林植被和凋落物的碳储量分别为(11.39±2.65)Mg/hm2和(1.22±0.29)Mg/hm2, 其中植被地上和地下碳储量为(6.29±1.83)Mg/hm2和(5.10±1.07)Mg/hm2(图 2)。与灌木林相比, 10年生林分植被地上、植被地下、整个植被和凋落物的碳储量低了5.4%、70.9%、34.7%和42.8%, 但只有植被地下碳储量是显著减少的(P < 0.01)(图 2)。林龄从10年到35年, 华北落叶松林植被地上、植被地下、整个植被和凋落物的碳储量分别增加了11.9、6.2、10.7倍和24.5倍, 虽然27年和35年的植被地下碳储量之间的差异并不显著(P=0.24)(图 2)。植被各层的碳储量随林龄的变化并不相同(表 3)。乔木及其器官的碳储量随林龄的增大而增加, 除树叶外, 它们在林龄之间的差异均显著(P < 0.05);灌木及其器官的碳储量呈先增加后减小的趋势;草本层及其器官的碳储量虽有起伏, 但总体呈减小的趋势。18年生林分植被地上、整个植被和凋落物的碳储量显著高于灌木林(P < 0.01), 而且此时的植被地下碳储量与之没有显著差异(P=0.89)。随着林龄的继续增大, 植被地下碳储量也超过灌木林(图 2)。但是, 无论林龄多大, 华北落叶松林灌木、草本及其器官的碳储量均显著小于灌木林(P < 0.01)(表 3)。

|
| 图 2 植被和凋落物碳储量随林龄的变化 Fig. 2 Carbon storage changes in vegetation and litter with stand age ●, 华北落叶松(Larix principis-rupprechtii Mayr.)人工林;○, 灌木林, 其为华北落叶松人工林的对照(CK);图中虚线表示灌木林某碳储量组分的平均水平;不同小写英文字母表示林龄间的差异显著(P < 0.05) |
| 层次 Layer |
器官 Component |
林龄Stand age/a | |||||
| CK | 10 | 18 | 23 | 27 | 35 | ||
| 乔木Tree | 树干 | ― | 2.49±0.67e | 15.34±2.04d | 24.91±2.54c | 35.66±6.79b | 63.74±6.04a |
| 树枝 | ― | 1.70±0.15e | 3.80±0.81d | 5.20±1.06c | 6.89±1.33b | 8.76±0.82a | |
| 树叶 | ― | 0.94±0.21d | 1.68±0.17c | 2.09±0.45bc | 2.43±0.65b | 3.25±0.71a | |
| 地上 | ― | 5.13±0.66e | 20.81±2.52d | 32.20±3.26c | 44.99±8.15b | 75.74±7.49a | |
| 树根 | ― | 0.62±0.17e | 3.63±1.03d | 6.12±1.52c | 8.64±1.51b | 10.25±0.90a | |
| 小计 | ― | 5.74±0.74e | 24.44±3.41d | 38.32±4.77c | 53.63±8.81b | 86.00±7.30a | |
| 灌木Shrub | 地上 | 5.88±1.81a | 0.59±0.06c | 1.75±0.54bc | 2.35±0.40b | 1.15±0.43bc | 0.68±0.31c |
| 地下 | 4.23±1.05a | 0.49±0.21c | 1.40±0.32b | 1.75±0.42b | 1.02±0.35bc | 0.29±0.13c | |
| 小计 | 10.11±2.68a | 1.08±0.27cd | 3.15±0.81bc | 4.10±0.70b | 2.17±0.74bcd | 0.96±0.43d | |
| 草本Herb | 地上 | 0.41±0.16a | 0.23±0.03b | 0.12±0.03bc | 0.12±0.04bc | 0.09±0.05c | 0.06±0.03c |
| 地下 | 0.87±0.41a | 0.38±0.12b | 0.18±0.04b | 0.15±0.04b | 0.06±0.03b | 0.14±0.02b | |
| 小计 | 1.28±0.54a | 0.62±0.15b | 0.30±0.04bc | 0.28±0.05bc | 0.16±0.08c | 0.21±0.05bc | |
| * CK是指灌木林, 其为华北落叶松(Larix principis-rupprechtii Mayr.)人工林的对照;同行不同小写英文字母表示林龄间的差异显著(P < 0.05) | |||||||
与灌木林相比, 华北落叶松林表层土壤(0—10 cm)的容重没有明显变化(P=0.30), 10—30 cm和30—50 cm土层的容重在造林初期也没有明显变化(P>0.05), 造林后期却显著减小(P < 0.05)(图 3)。然而, 土壤有机碳含量及储量在造林初期出现了不同程度的下降(P < 0.05)(图 3)。10年生林分不同土层的有机碳含量及储量相对于灌木林显著下降了27.1%—36.0%(P < 0.05), 土壤有机碳储量(0—50 cm)也明显下降了32.6%(P < 0.01)。随着林龄的增大, 华北落叶松林不同土层的有机碳含量及储量均呈增加的趋势, 而且相对于30—50 cm土层, 0—10 cm和10—30 cm土层有机碳含量和储量的增加更为明显(图 3)。土壤有机碳储量(0—50 cm)随着林龄的增大也逐渐增加, 虽然并非所有林龄之间的差异都显著(图 3)。18年生林分的表层土壤有机碳含量及储量与灌木林之间均没有显著差异(P>0.05), 但10—30 cm和30—50 cm土层有机碳含量及储量依然低于灌木林(P < 0.05), 土壤有机碳储量(0—50 cm)也低于灌木林(P < 0.05)(图 3)。23年生林分的10—30 cm和30—50 cm土层有机碳含量及储量与灌木林之间均没有显著差异(P>0.05), 而且土壤有机碳储量(0—50 cm)也与之没有显著差异(P>0.05)。随着林龄的继续增大, 土壤有机碳含量及储量不断积累, 并将全面超过灌木林(图 3)。

|
| 图 3 不同土层土壤容重、土壤有机碳含量和土壤有机碳储量随林龄的变化 Fig. 3 Changes in soil bulk density, soil organic carbon content and soil organic carbon storage with stand age ●, 华北落叶松(Larix principis-rupprechtii Mayr.)人工林;○, 灌木林, 其为华北落叶松人工林的对照(CK);图中横虚线表示灌木林某土壤特征的平均水平;不同小写英文字母表示林龄间的差异显著(P < 0.05) |
灌木林生态系统碳储量为(125.61±18.55)Mg/hm2, 其中植被、凋落物和土壤平均储存了生态系统碳储量的9.1%、1.0%和89.9%(图 4)。与灌木林相比, 10年生林分的生态系统碳储量显著减少了32.9%(P < 0.01)。林龄从10年到35年, 华北落叶松林的生态系统碳储量由(84.27±8.65)Mg/hm2增加到(219.82±20.29)Mg/hm2(图 4), 其中植被和凋落物占生态系统碳储量的比例从8.9%和0.9%增加到39.7%和8.1%, 土壤则从90.3%下降到52.2%(图 4)。18年生林分的生态系统碳储量与灌木林之间没有显著差异(P=0.83), 之后随着林龄的增大, 生态系统碳储量逐渐增加并超过灌木林(图 4)。

|
| 图 4 生态系统碳储量及其分配随林龄的变化 Fig. 4 Changes in ecosystem carbon storage and its allocation with stand age ●, 华北落叶松(Larix principis-rupprechtii Mayr.)人工林;○, 灌木林, 其为华北落叶松人工林的对照(CK);图中虚线表示灌木林生态系统碳储量的平均水平;不同小写英文字母表示林龄间的差异显著(P < 0.05) |
用何种方式在何处开展植被恢复与重建, 不仅是国内外学者普遍关注的理论和实践问题[4, 16], 也是林业碳汇项目开发中一个需要首先明确的问题[17]。本研究围绕着退化灌木林转变为乔木林的固碳效应, 以关帝山林区为例, 从生态系统的角度, 探讨了这种转变对生态系统碳储量及其组分的影响。
本研究中, 华北落叶松各器官的含碳量(50.0%—51.7%)略高于两个常用于估算森林植被碳储量的含碳量缺省值(45.0%和50.0%)[18], 灌木的含碳量(47.7%—48.6%)介于这两个缺省值之间, 但草本(38.6%—42.2%)和凋落物(41.4%)的含碳量均低于这两个缺省值。此外, 乔木(华北落叶松)的平均含碳量(50.7%)>灌木(48.2%)>草本(40.4%), 这与已有结论相一致[18]。因此, 为降低森林植被碳储量估算的不确定性, 应优先考虑植物含碳量的本地值, 而非区域乃至全球的缺省值。
自然状态下, 随着林龄的增大, 植被碳储量(生物量)逐渐增加, 而且乔木层占植被碳储量的比例也随之增加[19-20]。林龄从10年到35年, 华北落叶松林植被碳储量增加了10.7倍, 乔木层占植被碳储量的比例从77.2%上升到98.6%。华北落叶松林凋落物碳储量也随林龄的增大而增加, 这与罗云建等[20]研究中国森林生态系统生物量及其分配格局的结论相一致。与地上生物量碳库和凋落物碳库相比, 地下生物量碳库需要更长的时间才能恢复到造林前(灌木林)的碳水平(图 2)。然而, 随林龄的增大, 华北落叶松林林下层(灌木和草本)碳储量呈现先增加后减小的趋势(表 3)。这是因为, 林冠郁闭前, 由于林内光照充足、林木竞争不充分等原因, 林下层生物量和碳储量呈增加的趋势, 但随着林龄的增大, 林冠也从相接到郁闭, 林木个体和光照竞争加剧, 林下环境发生了较大的变化, 林下层生物量和碳储量也随之下降。
关于造林/再造林对土壤有机碳储量的影响及相关机制, 目前的认知仍然不够全面[21]。当植被和土壤管理措施发生变化时, 土壤有机碳含量变化的方向和速率主要取决于动植物残体、凋落物、根系分泌物和微生物残体及其代谢产物的输入与输出的动态平衡[22-23]。造林/再造林活动如果有利于以下过程, 将促进土壤有机物质的积累, 提高土壤对碳的固定:①土壤中有机物质输入速率的增加;②易分解性有机物质输入的增加;③土壤生物活性的提高, 增加土壤深层有机物质的输入;④形成土壤团聚体、有机-矿物复合体, 增强土壤有机物质的物理性保护。理论上, 造林/再造林初期由于林木较小, 凋落物归还也少, 加之造林过程的扰动, 加速了土壤有机物质的氧化分解, 以致土壤有机碳含量和储量出现不同程度的下降。随着林龄的增大, 凋落物、地下根系以及土壤生物种类和数量的逐渐增加, 促使输入土壤的有机物质增加, 同时加快了有机物质的周转, 这些都有利于土壤有机碳的积累及向土壤深层的输入, 使得土壤有机碳逐渐恢复。大量研究表明, 与前期土地利用方式相比, 造林/再造林后土壤有机碳通常先下降后慢慢积累, 开始积累的时间在温带地区一般少于10年, 而在热带地区要晚一些[9-13]。造林后土壤有机碳何时开始积累并能否达到前期土地利用方式的水平, 是由前期土地利用方式、气候、林龄、树种、土壤类型、管理措施等因素共同控制的[9-13]。通过研究退化灌木林转变为华北落叶松林对土壤有机碳的影响, 我们发现, 与灌木林相比, 不同土层的有机碳含量及储量在造林初期出现了不同程度的下降, 但其随着林龄的增大而增加甚至超过灌木林, 而且深层土壤有机碳相对于表层(0—10 cm)需要更长的时间才能恢复到造林前的水平(图 3)。
与前期土地利用方式相比, 造林后的生态系统碳储量及其组成需要多久才能达到其碳储量水平, 目前还不很清楚[24]。Liao等[12]通过收集全球天然林和人工林的对比研究案例, 发现人工林地上生物量、地下生物量、土壤有机碳储量和生态系统碳储量平均下降了34.4%、40.0%、15.5%和28.8%, 而且相比于地上碳库, 地下碳库的损失更大, 其恢复可能需要更长的时间。我们也证实了这一点, 与植被地上碳库相比, 植被地下碳库需要更长的时间才能恢复造林前的碳水平(图 2), 而且土壤有机碳库的恢复时间比植被地下碳库还要长, 其中深层土壤有机碳的恢复时间大于表层土壤有机碳(0—10 cm)(图 3)。由此可见, 要恢复到前期土地利用方式的碳水平, 生态系统各个碳库需要的时间存在较大差异。
| [1] | 谢宗强, 唐志尧. 中国灌丛生态系统碳储量的研究. 植物生态学报, 2017, 41(1): 1–4. |
| [2] | Fang J Y, Guo Z D, Piao S L, Chen A P. Terrestrial vegetation carbon sinks in China, 1981-2000. Science in China Series D:Earth Sciences, 2007, 50(9): 1341–1350. DOI:10.1007/s11430-007-0049-1 |
| [3] | 邓东周, 张小平, 鄢武先, 慕长龙. 低效林改造研究综述. 世界林业研究, 2010, 23(4): 65–69. |
| [4] | Gao Y, Zhu X J, Yu G R, He N P, Wang Q F, Tian J. Water use efficiency threshold for terrestrial ecosystem carbon sequestration in China under afforestation. Agricultural and Forest Meteorology, 2014, 195-196: 32–37. DOI:10.1016/j.agrformet.2014.04.010 |
| [5] | Garcia-Franco N, Wiesmeier M, Goberna M, Martínez-Mena M, Albaladejo J. Carbon dynamics after afforestation of semiarid shrublands:implications of site preparation techniques. Forest Ecology and Management, 2014, 319: 107–115. DOI:10.1016/j.foreco.2014.01.043 |
| [6] | 程彩芳, 李正才, 周君刚, 吴亚丛, 赵志霞, 孙娇娇. 北亚热带地区退化灌木林改造为人工阔叶林后土壤活性碳库的变化. 林业科学研究, 2015, 28(1): 101–108. |
| [7] | 商素云, 李永夫, 姜培坤, 周国模, 刘娟, 吴家森, 林琳. 天然灌木林改造成板栗林对土壤碳库和氮库的影响. 应用生态学报, 2012, 23(3): 659–665. |
| [8] | Bustamante M, Robledo-Abad C, Harper R, Mbow C, Ravindranat N H, Sperling F, Haberl H, Pinto A D S, Smith P. Co-benefits, trade-offs, barriers and policies for greenhouse gas mitigation in the agriculture, forestry and other land use (AFOLU) sector. Global Change Biology, 2014, 20(10): 3270–3290. DOI:10.1111/gcb.2014.20.issue-10 |
| [9] | Bárcena T G, Kiær L P, Vesterdal L, Stefánsdóttir H M, Gundersen P, Sigurdsson B D. Soil carbon stock change following afforestation in Northern Europe:a meta-analysis. Global Change Biology, 2014, 20(8): 2393–2405. DOI:10.1111/gcb.2014.20.issue-8 |
| [10] | Deng L, Liu G B, Shangguan Z P. Land-use conversion and changing soil carbon stocks in China's 'Grain-for-Green' Program:a synthesis. Global Change Biology, 2014, 20(11): 3544–3556. DOI:10.1111/gcb.12508 |
| [11] | Li D J, Niu S L, Luo Y Q. Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation:a meta-analysis. New Phytologist, 2012, 195(1): 172–181. DOI:10.1111/j.1469-8137.2012.04150.x |
| [12] | Liao C Z, Luo Y Q, Fang C M, Li B. Ecosystem carbon stock influenced by plantation practice:implications for planting forests as a measure of climate change mitigation. PLoS One, 2010, 5(5): e10867. DOI:10.1371/journal.pone.0010867 |
| [13] | Liao C Z, Luo Y Q, Fang C M, Chen J K, Li B. The effects of plantation practice on soil properties based on the comparison between natural and planted forests:a meta-analysis. Global Ecology and Biogeography, 2012, 21(3/4): 318–327. |
| [14] | 吴中伦. 中国森林(第2卷):针叶林. 北京: 中国林业出版社, 1999. |
| [15] | 罗云建, 王效科, 逯非. 中国主要林木生物量模型手册. 北京: 中国林业出版社, 2015. |
| [16] | Cao S X. Why large-scale afforestation efforts in China have failed to solve the desertification problem. Environmental Science and Technology, 2008, 42(6): 1826–1831. DOI:10.1021/es0870597 |
| [17] | 张小全, 武曙红. 林业碳汇项目理论与实践. 北京: 中国林业出版社, 2010. |
| [18] | 郑帷婕, 包维楷, 辜彬, 何晓, 冷俐. 陆生高等植物碳含量及其特点. 生态学杂志, 2007, 26(3): 307–313. |
| [19] | Lorenz K, Lal R. Carbon Sequestration in Forest Ecosystems. Dordrecht: Springer, 2010. |
| [20] | 罗云建, 王效科, 张小全, 逯非. 中国森林生态系统生物量及其分配研究. 北京: 中国林业出版社, 2013. |
| [21] | Mobley M L, Lajtha K, Kramer M G, Bacon A R, Heine P R, Richter D D. Surficial gains and subsoil losses of soil carbon and nitrogen during secondary forest development. Global Change Biology, 2015, 21(2): 986–996. DOI:10.1111/gcb.12715 |
| [22] | Lal R. Forest soils and carbon sequestration. Forest Ecology and Management, 2005, 220(1/3): 242–258. |
| [23] | Post W M, Kwon K C. Soil carbon sequestration and land-use change:processes and potential. Global Change Biology, 2000, 6(3): 317–327. DOI:10.1046/j.1365-2486.2000.00308.x |
| [24] | Pang X Y, Huang J S, Zhao Q X, Feng D F, Bao W K, Tian G L. Ecosystem carbon stock across a chronosequence of spruce plantations established on cutovers of a high-elevation region. Journal of Soils and Sediments, 2017, 17(9): 2239–2249. DOI:10.1007/s11368-016-1415-4 |