 2018, Vol. 38
2018, Vol. 38文章信息
- 游桂莹, 张志渊, 张仁铎.
- YOU Guiying, ZHANG Zhiyuan, ZHANG Renduo.
- 全球陆地生态系统光合作用与呼吸作用的温度敏感性
- Temperature sensitivity of photosynthesis and respiration in terrestrial ecosystems globally
- 生态学报. 2018, 38(23): 8392-8399
- Acta Ecologica Sinica. 2018, 38(23): 8392-8399
- http://dx.doi.org/10.5846/stxb201801100071
-
文章历史
- 收稿日期: 2018-01-10
- 网络出版日期: 2018-09-26

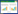




2. 福建农林大学资源与环境学院, 福州 350002
2. Fujian Agriculture and Forestry University, College of Resources and Environmental, Fuzhou 350002, China
生态系统呼吸(Re, 包括地上植物呼吸和土壤呼吸)是生态系统碳循环的主要成分, 该过程将生态系统初级生产力(GPP)固定的大部分碳以CO2的形式释放到大气中[1-2]。Re和GPP都受温度的影响[3-4]。陆地生态系统主要通过光合作用、呼吸作用与大气进行碳交换, 在碳平衡状态下表现为净初级生产力为0(即碳排放等于碳吸收)[5]。不过, 光合作用和呼吸作用对温度响应程度的不同决定生态系统表现为碳源或者碳汇。有研究表明, 温度升高使得生态系统生产力提高了15.7%, 生态系统呼吸增加了6.0%, 最终使得生态系统表现为一个微弱的碳汇[6]。也有研究发现, 温度升高时, 土壤呼吸的增加速率大于净初级生产力的增加速率, 此时, 生态系统表现为碳源[7]。
陆地生态系统与全球气候变化之间的反馈作用会改变陆地生态系统碳源汇之间的平衡, 进而影响未来气候变化。陆地生态系统碳吸收潜力取决于生态系统的类型和环境状况, 即取决于生态系统的物种组成、结构和年龄分布等, 以及所处的纬度区域下的环境, 包括当地气候和土壤状况等。当环境温度高于植物光合作用的最适温度时, 植物的光合速率降低, 而但环境温度低于植物光合的最适温度时, 光合速率随温度的上升而增加[8]。而不同的生态系统间植物的生长条件不同会导致不同植被类型对升温有不同的响应(如土壤水分的可利用性可能会控制光合作用对温度的响应)[9], 最终可导致Q10值(即温度每升高10℃, 光合或呼吸速率增加的倍数, 通常用来表征光合或呼吸作用的温度敏感性)[10]的变化。光合作用和呼吸过程的温度敏感性(Q10, G和Q10, R)是理解生态系统碳平衡的重要因子[11], 对光合作用和呼吸作用的温度敏感性研究有助于解释生态系统过程对气候变化的响应机制。本研究旨在通过对收集到的全球碳通量数据进行分析, 对比全球陆地生态系统光合作用与呼吸作用对温度变化的响应, 分析不同纬度、不同植被类型下光合作用和呼吸作用的温度敏感性分布规律, 为深入理解全球气候变化下陆地生态系统碳循环变化提供有价值的信息。
1 材料与方法数据来源于全球各碳通量网的生态系统呼吸数据库中收集的数据。数据库中通量数据点分布如图 1所示, 主要包括全球通量网FLUXNET数据库(http://www.fluxdata.org/, 主要是Open Dataset)、欧洲通量数据库(European Fluxes Database Cluster, http://gaia.agraria.unitus.it/)、美国橡树岭国家实验室生物地球化学动力学分布式数据存档中心ORNL DAAC(http://daac.ornl.gov/)、亚洲数据库AsiaFlux(http://asiaflux.net/)、日本通量网FFPRI Fluxnet(http://www2.ffpri.affrc.go.jp/labs/flux/)、美国通量网AmeriFlux(http://ameriflux.lbl.gov/)。当不同数据库之间通量点有重叠的时候, 以较大的数据库数据为准, 如FLUXNET(最大的数据库)。收集的数据变量类型包括环境变量和物理变量, 主要有生态系统CO2交换量(NEE)、总初级生产力(GPP)、生态系统呼吸(Re)空气温度、土壤温度、经纬度、降水、土壤含水量、太阳辐射、植被类型与生态系统类型等。共收集647套通量数据, 涵盖全球从38°S到71°N不同区域的共130个地点, 从1991到2007年的通量监测数据。南半球9套数据, 主要集中在低纬度(30°S—40°S)。北半球638套数据, 低纬度37套, 中纬度548套, 高纬度53套。数据库涵盖13种植被类型和7种气候类型。主要植被类型有:农田(croplands, CRO, 65套), 落叶阔叶林(deciduous broad-leaf forests, DBF, 111套), 常绿阔叶林(evergreen broad-leaf forests, EBF, 39套), 落叶针叶林(deciduous coniferous forest, ENF, 229套), 草地(grassland, GRA, 100套), 混交林(mixed forests, MF, 31套), 开阔灌丛(open shrublands, OSH, 18套), 永久湿地(permanent wetlands, WET, 27套)。主要气候类型有:温带(348套)、寒带(102套)、亚热带-地中海气候(160套)、热带(14套)。

|
| 图 1 研究中收集到的碳通量数据站点的地理分布(引自http://fluxnet.fluxdata.org/sites/site-summary/[12]) Fig. 1 Geographical distributions of the eddy flux tower sites involved in this study(cited from http://fluxnet.fluxdata.org/sites/site-summary/[12]) |
由涡度相关法测得的数据通常整合成半小时测量的通量数据[13]。在此基础上, 再计算从每日到每年的不同时间尺度的碳通量。涡度相关法测得的有效数据量大约只有总数的70%, 其中有效的夜间CO2通量数据所占夜间数据的比例更低。涡度相关法所测的结果存在很大的不确定性, 尤其在夜间。主要原因是由于夜间, 空气层通常比较稳定, 无法满足涡度相关观测空气强对流的假设, 导致涡度相关仪器无法响应, 从而造成夜间碳通量测量的不确定性, 因此需要对夜间的观测数据进行校正[14]。本研究中所收集的数据中采用国际通量网的通用方法进行校正, 即利用温度和u*观测值之间的关系来确定u*的临界值, 从而筛选出符合质量的数据[15]。该方法中, 首先将全年夜间数据等分为6个温度组, 每个温度组再等分成20个u*小组。如果某个小组中存在某一夜间碳通量大于本温度组中后面所有小组平均通量的99%, 则记录该通量值对应的u*值。当该温度组中, u*与温度不相关或者弱相关(r < |0.4|)时, 记录的u*则为该温度组的u*临界值。最后全年的u*临界值为6个温度组u*的中位数。同时, 为降低由传统插值法带来的误差, 研究中采用迭代奇异谱分析(Singular Spectrum Analysis, 简称SSA)对原序列进行插值。SSA插值法值利用序列的时间相关性, 将时间序列分解成不同周期, 产生新的不同时间尺度的序列, 利用新的时间序列对原序列空缺处进行插值, 反复迭代, 最终产生误差最小的序列。其中, 日均生态系统呼吸量是夜间每半小时所测CO2通量的均值。但是每天的夜间数据中至少有5个测量值, 才可用来计算该日均呼吸量[16]。日均温度则取相应的CO2通量对应的夜间空气温度均值。
1.2 Q10值的计算代谢速率和温度的关系可用以下经验模型来表征[17]:
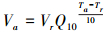
|
(1) |
式中, Ta、Tr分别代表是周围空气温度和参考温度(℃), 本研究中设Tr=15℃, Va和Vr分别是对应于温度为Ta和Tr的生态系统代谢量(gC m-2 d-1), Q10是代谢速率的温度敏感性系数。将式1两边取对数可得:
|
(2) |
式中, 将lnVa和Ta进行线性回归便可计算Q10值。
2 结果与讨论在全球尺度下, Q10, G和Q10, R的均值分别是3.99±0.21和2.28±0.074。其95%的置信区间分别为(3.78, 4.20)和(2.20, 2.35)。Q10, G和Q10, R的变异系数分别是67%和41%, 即在全球尺度下, 相较于光合作用, 呼吸作用的温度敏感性波动较小。
2.1 温度、纬度对Q10值的影响研究表明(图 2、图 3), Q10, G和Q10, R都与年均温度呈负相关(即与纬度呈正相关)。其中, Q10, G和Q10, R对温度的相关系数别为-0.19(P < 0.05)、-0.04(P < 0.05)。对Q10值与经纬度的相关性进行分析可得, Q10, G和Q10, R与经度都呈不相关, 而与纬度存在显著相关(P < 0.05)。这是由于在低纬度区域年均温度较高, 生态系统代谢速率本身较高, 因此, 呼吸和光合作用速率随温度升高的幅度相对较小, 导致较小Q10值。在高纬度地区, 生态系统对气候变暖更加敏感, 这是由于高纬度地区生态系统严格受到温度和营养物质的限制[18-19]。此外, 温度的升高会使植物的生长期延长, 冰川退缩、永冻土融化等[20], 通常, 较寒冷的区域比温度高的区域的活化能高对应较大的Q10值[21], 这些因素也可使得高纬度地区Q10值较大。

|
| 图 2 Q10, G值和Q10, R值与年均温度的关系 Fig. 2 The relationships between mean annual temperature and the value of Q10, G、Q10, R |

|
| 图 3 Q10, G值和Q10, R值与纬度的关系 Fig. 3 The relationships between latitude and the value of Q10, G、Q10, R |
在纬度上, Q10, G和Q10, R都表现为高纬>中纬>低纬, 即随着纬度的增加, 光合和呼吸作用对温度的敏感性增加, 这与全球尺度下的纬度分析结果一致。在低纬度带上, Q10, G小于Q10, R, 而在中、高纬度上, Q10, G大于Q10, R(表 1)。在低纬度, 平均空气温度比较高(大部分地点的年平均空气温度都大于20℃), 因此生态系统呼吸速率本身就很高, 在这种情况下, 呼吸随温度升高而增加的幅度是有限的, 导致Q10, R值较小, 而在较高纬度区域, 空气温度相对较低, 呼吸对温度变化的敏感性较高, 随温度升高而增加的幅度较大, 导致Q10, R较大。
| 纬度区域 Latitude regions |
样本数 Sample size |
Q10 | 均值 Average value |
标准差 Standard deviation |
变异系数 Variable coefficient/% |
95%置信区间 95% Confidence intervals |
|
| 北半球 | 低纬度 | 37 | Q10, G | 1.40 | 0.75 | 57 | (1.12, 1.68) |
| The northern hemisphere | Q10, R | 1.73 | 0.29 | 53 | (1.59, 1.88) | ||
| 中纬度 | 548 | Q10, G | 3.99 | 2.65 | 74 | (3.76, 4.22) | |
| Q10, R | 2.21 | 0.78 | 35 | (2.14, 2.27) | |||
| 高纬度 | 53 | Q10, G | 5.54 | 0.88 | 16 | (5.22, 5.86) | |
| Q10, R | 2.73 | 0.29 | 11 | (2.63, 2.84) | |||
| 南半球 | 中纬 | 9 | Q10, G | 2.06 | 0.37 | 18 | (1.78, 2.34) |
| The southern hemisphere | Q10, R | 1.42 | 0.72 | 42 | (0.49, 1.38) | ||
| Q10, G和Q10, R分别表示光合作用和呼吸作用的Q10值 | |||||||
不同植被类型的Q10, G和Q10, R分布如图 4, 同一种植被类型下(除多树热带草原、常绿阔叶林外), 对应的Q10, G都大于Q10, R。Q10, G变化幅度较大, 为1.34—12.44, 其中热带稀树草原的值最大。与Q10, G相比, 不同植被类型, Q10, R变化幅度较小, 为1.44—3.27。而Q10, G和Q10, R值最小都分布在常绿阔叶林。分析不同植被类型中Q10值的分布, 可知:针叶林>阔叶林。

|
| 图 4 不同植被类型下的Q10值 Fig. 4 Q10 values among different plant functional types 植被类型:CRO:农田, croplands;CSH:密闭灌丛, closed shrublands;DBF:落叶阔叶林, deciduous broad-leaf forests;EBF:常绿阔叶林, evergreen broad-leaf forests;ENF:常绿针叶林, evergreen needle-leaf forests;GRA:草地, grasslands;MF:混交林, mixed forests;OSH:稀疏灌丛, open shrublands;SAV:热带稀树草原, savannas;WET:永久湿地, permanent wetlands;WSA:多树热带草原, woody savannas |
在3个不同纬度区域, 不同植被类型下的温度敏感性存在差异, 尤其体现在光合作用的温度敏感性。一般地, 类似的气候条件, 落叶林的植物活动和物候现象比常绿林表现出更大的季节变异性。对于呼吸作用, 春季落叶林的根生长活动比常绿林剧烈, 使得落叶林的土壤呼吸由异养呼吸为主导转变为以自养呼吸为主导, 而呼吸成分的改变导致了较高的温度敏感性[15]。同时, 植被类型对呼吸温度敏感性的影响是因为微生物群落和有机碳成分不同, 从而影响呼吸对温度的响应[22]。而增温对植物的光合作用生理的影响也因植物种类而异[23]。升温会增加土壤、植物碳库(包括地上和地下部分)及凋落物碳库的质量损失, 而不同生态系统、不同植被类型下各碳库对温度的响应不同[6], 最终导致Q10不同。
2.3 植被类型和纬度分布对Q10值的复合影响由表 2可知, 在不同的纬度区域, 植被类型对Q10, G和Q10, R的影响程度存在差异。在低纬度区域, 多树热带草原的Q10, G值最大, 是常绿阔叶林的两倍;Q10, R是常绿阔叶林的两倍以上。在中纬度区域, 植被类型丰富, Q10值对植被类型的响应又不同于低纬度区域。由表 2知, 同一纬度区域下, 各植被类型(常绿阔叶林除外)中, Q10, G值都大于Q10, R值。农田的Q10, R最大, 多树热带草原的Q10, R最小, 可能是人工干扰给生态系统呼吸带来了更大的变异性, 因此导致了较高的Q10, R, 表现出高度的变异性。而落叶阔叶林的Q10, G值最大, 多树热带草原的Q10, G最小。在高纬度区域, Q10, G值值大于Q10, R值, 与中纬度区域分析结果一致, 但在高纬度地区, 热带稀树草原中Q10, R值大于Q10, G值, 原因可能是由于热带稀树草原样本较少(N=3), 因此未在表 2中列举出来。
| 植被类型 Vegetation types |
低纬度Low latitude | 中纬度Middle latitude | 高纬度High latitude | ||||||||
| Q10, G | Q10, R | 样本数 Sample size |
Q10, G | Q10, R | 样本数 Sample size |
Q10, G | Q10, R | 样本数 Sample size |
|||
| 农田(CRO) | — | — | — | 5.19±1.21 | 3.27±0.60 | 65 | — | — | — | ||
| 密闭灌丛(CSH) | — | — | — | 8.00±3.66 | 2.61±0.41 | 5 | — | — | — | ||
| 落叶阔叶林(DBF) | — | — | — | 8.03±0.89 | 2.38±0.19 | 221 | — | — | — | ||
| 常绿阔叶林(EBF) | 1.33±0.34 | 0.75±0.92 | 6 | 1.34±0.19 | 1.56±0.25 | 66 | — | — | — | ||
| 落叶针叶林(ENF) | 1.50±0.22 | 1.78±0.22 | 15 | 3.49±0.32 | 2.63±0.27 | 179 | 6.09±0.82 | 2.75±0.29 | 35 | ||
| 草地(GRA) | — | — | — | 3.40±0.62 | 3.07±0.48 | 100 | — | — | — | ||
| 混交林(MF) | — | — | — | 4.70±0.74 | 2.42±0.30 | 31 | — | — | — | ||
| 稀疏灌丛(OSH) | — | — | — | 2.82±0.87 | 2.39±0.57 | 15 | — | — | — | ||
| 永久湿地(WET) | — | — | — | 4.64±0.66 | 2.34±0.50 | 13 | 6.69±1.83 | 2.69±0.33 | 12 | ||
| 多树热带草原(WSA) | 2.70±1.87 | 3.57±1.64 | 10 | 1.04±0.24 | 0.82±0.43 | 6 | — | — | — | ||
| CRO: croplands; CSH: closed shrublands; DBF:deciduous broad-leaf forests; DNF: deciduous needle leaf forests; EBF: evergreen broadleaf forest; ENF:evergreen needle-leaf forests; GRA: grasslands; MF: mixed forests; OSH: open shrublands; SAV: savannas; WET: permanent wetlands; WSA: woody savannas | |||||||||||
由表 2得知, 相同植被类型下的Q10, G和Q10, R对纬度变化的响应可能存在差异, 如常绿针叶林的Q10, G和Q10, R都随纬度升高而升高, 而多树热带草原则相反。这是由于植被类型和纬度因素对温度敏感性的复合影响, 且在不同情况下, 主导影响因素可能不同, 使得Q10值的实际分布更加复杂。通过双因素方差分析发现, 在不考虑纬度因素与植被类型间的交互效应时, 无论是Q10, G还是Q10, R, 植被类型对Q10的总变异贡献更大(Q10, G:ηa2=0.034>ηb2=0.011;Q10, R:ηa2=0.269>ηb2=0.136;其中ηa2、ηb2分别代表植被类型、纬度的效应量估计)。
2.4 不同气候带和气候类型对Q10值的影响由图 5可知, 干旱带下的Q10, G最小, 而冷温带的Q10, G最高。而Q10, R在不同气候带下相对较稳定。由均值分析知, 不同气候带下, Q10, G的变异系数为65%, 而Q10, R的变异系数高达95%。

|
| 图 5 Q10值在不同气候带下的分布 Fig. 5 Q10 values among different climatic zones |
根据柯本气候分类法, 气候带:A:热带, B:干旱带(低纬度、低海拔);C:暖温带, D:冷温带, 降水:m:季风型, w:冬天旱季型, s:夏天旱季型, f:湿地型, S:草原型, W:沙漠型气温:h:炎热干燥, k:寒冷干燥, a:夏季炎热, b:夏季温暖, c:夏季凉快;Cfb1:海洋西海岸:温和无旱季, 温暖的夏天在气候带上, 干旱带(低纬度、低海拔)的Q10, G最小, 而冷温带的Q10, G最高, 这一结果与纬度、温度分布分析结果一致, 而Q10, R相对较稳定, 这可能是因为高纬度地区低温的影响以及较少的植被类型所导致的。不同气候类型下的Q10值分布如图 6所示, (温暖冬干气候下的Q10, G为28.13, Q10, R为-0.37, 与其他气候类型相差太大, 可能是样本数较少所致(N=3), 故不在图中标记), Q10, G的变化范围在0.38—9.29之间, Q10, R的变化范围为0.15—5.76。除草原气候、温暖夏干气候外, Q10, G都大于Q10, R。此外, 还可以得知, 在干燥条件下对应较低的Q10, G, 而Q10, R值在湿度较大的区域较小(Cfc除外)。

|
| 图 6 不同气候类型下的Q10值 Fig. 6 Q10values among different climate types |
不同气候带下的Q10, G表现出较大的差异, 而Q10, R相对较稳定。干旱带下的Q10, G最小, 这可能是由于在干旱条件下, 植物光合作用及生态系统呼吸作用容易受水分限制的影响, 此时温度不再是主导因素, 甚至在极端条件下, 几乎不受温度影响。而冷温带下, 年均温度较低, 在未达到植物最适生长温度时, 光合作用随温度的升高而升高, 即对应较大的Q10, G值。
3 结论(1) 光合作用和呼吸作用Q10值都存在空间变异性。在95%的置信区间下, Q10, G值在北半球低、中、高纬度3个区域的范围分别是1.12—1.68、3.76—4.22、5.22—5.86;Q10, R值在低、中、高纬度3个区域的范围分别是1.59—1.88、2.14—2.27、2.63—2.84。在南半球中纬Q10, G值与Q10, R值的范围分别是1.78—2.34、0.49—1.38。
(2) 从Q10值的纬度分布规律来看, 无论是光合作用还是呼吸作用, 在全球尺度下, Q10值都随纬度的升高而升高。在纬度上, Q10, G和Q10, R都表现为高纬>中纬>低纬, 即随着纬度的增加, 光合和呼吸作用对温度的敏感性增加。
(3) 植被类型会影响光合作用和呼吸作用的Q10值分布, 除热带多树草原、常绿落叶林外, Q10, G都大于Q10, R。分析不同植被类型中Q10值的分布, 可以得出:针叶林>阔叶林。
(4) 不同纬度区域, 植被类型对Q10值的影响不同。当植被类型和纬度区域共同影响Q10值时, 植被类型对Q10值的总变异贡献最大。
(5) 气候带和气候类型对Q10, G和Q10, R都有显著影响。在气候带上, 干旱带(低纬度、低海拔)的Q10, G最小, 而冷温带的Q10, G最高。不同气候条件下(除温带草原气候外)的Q10, G都大于Q10, R。
| [1] | King A W, Gunderson C A, Post W M, Weston D J, Wullschleger S D. Plant respiration in a warmer world. Science, 2006, 312(5773): 536–537. DOI:10.1126/science.1114166 |
| [2] | Piao S L, Luyssaert S, Ciais P, Janssens I A, Chen A P, Cao C, Fang J Y, Friedlingstein P, Luo Y Q, Wang S P. Forest annual carbon cost:a global-scale analysis of autotrophic respiration. Ecology, 2010, 91(3): 652–661. DOI:10.1890/08-2176.1 |
| [3] | Mahecha M D, Reichstein M, Carvalhais N, Lasslop G, Lange H, Seneviratne S I, Vargas R, Ammann C, Arain M A, Cescatti A, Janssens I A, Migliavacca M, Montagnani L, Richardson A D. Response to comment on "global convergence in the temperature sensitivity of respiration at ecosystem level". Science, 2011, 331(6022): 1265. |
| [4] | Ciais P, Reichstein M, Viovy N, Granier A, Ogée J, Allard V, Aubinet M, Buchmann N, Bernhofer C, Carrara A, Chevallier F, De Noblet N, Friend A D, Friedlingstein P, Grünwald T, Heinesch B, Keronen P, Knohl A, Krinner G, Loustau D, Manca G, Matteucci G, Miglietta F, Ourcival J M, Papale D, Pilegaard K, Rambal S, Seufert G, Soussana J F, Sanz M J, Schulze E D, Vesala T, Valentini R. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature, 2005, 437(7058): 529–533. DOI:10.1038/nature03972 |
| [5] | Braswell B H, Schimel D S, Linder E, Moore Ⅲ B. The response of global terrestrial ecosystems to interannual temperature variability. Science, 1997, 278(5339): 870–873. DOI:10.1126/science.278.5339.870 |
| [6] | Lu M, Zhou X H, Yang Q, Li H, Luo Y Q, Fang C M, Chen J K, Yang X, Li B. Responses of ecosystem carbon cycle to experimental warming:a meta-analysis. Ecology, 2013, 94(3): 726–738. DOI:10.1890/12-0279.1 |
| [7] | Huntingford C, Cox P M, Lenton T M. Contrasting responses of a simple terrestrial ecosystem model to global change. Ecological Modelling, 2000, 134(1): 41–58. DOI:10.1016/S0304-3800(00)00330-6 |
| [8] | 王为民, 王晨, 李春俭, 林伟宏. 大气二氧化碳浓度升高对植物生长的影响. 西北植物学报, 2000, 20(4): 676–83. DOI:10.3321/j.issn:1000-4025.2000.04.030 |
| [9] | Niu S L, Li Z X, Xia J Y, Han Y, Wu M Y, Wan S Q. Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of northern China. Environmental and Experimental Botany, 2008, 63(1/3): 91–101. |
| [10] | Davidson E A, Janssens I A, Luo Y Q. On the variability of respiration in terrestrial ecosystems:moving beyond Q. Global Change Biology, 2006, 12(2): 154–164. |
| [11] | Vesala T, Launiainen S, Kolari P, Pumpanen J, Sevanto S, Hari P, Nikinmaa E, Kaski P, Mannila H, Ukkonen E, Piao S L, Ciais P. Autumn temperature and carbon balance of a boreal Scots pine forest in Southern Finland. Biogeosciences, 2010, 7(1): 163–176. |
| [12] | FLUXNET 2015 Dataset. http://fluxnet.fluxdata.org/sites/site-summary/. |
| [13] | Goulden M L, Munger J W, Fan S M, Daube B C, Wofsy S C. Measurements of carbon sequestration by long-term eddy covariance:methods and a critical evaluation of accuracy. Global Change Biology, 1996, 2(3): 169–182. DOI:10.1111/gcb.1996.2.issue-3 |
| [14] | Massman W J, Lee X. Eddy covariance flux corrections and uncertainties in long-term studies of carbon and energy exchanges. Agricultural and Forest Meteorology, 2002, 113(1/4): 121–144. |
| [15] | Griffis T J, Black T A, Gaumont-Guay D, Drewitt G B, Nesic Z, Barr A G, Morgenstern K, Kljun N. Seasonal variation and partitioning of ecosystem respiration in a southern boreal aspen forest. Agricultural and Forest Meteorology, 2004, 125(3/4): 207–223. |
| [16] | Mahecha M D, Reichstein M, Carvalhais N, Lasslop G, Lange H, Seneviratne S I, Vargas R, Ammann C, Arain M A, Cescatti A, Janssens I A, Migliavacca M, Montagnani L, Richardson A D. Global convergence in the temperature sensitivity of respiration at ecosystem level. Science, 2010, 329(5993): 838–840. DOI:10.1126/science.1189587 |
| [17] | W J. Lectures on theoretical and physical chemistry leçons de chimie physique. Nature, 1901, 63(1632): 343–344. DOI:10.1038/063343a0 |
| [18] | Bliss L C, Heal O W, Moore J J. Tundra Ecosystems:A Comparative Analysis. Cambridge: Cambridge University Press, 1981. |
| [19] | Chabot B F, Mooney H A. Physiological Ecology of North American Plant Communities. New York: Chapman and Hall, 1985. |
| [20] | 王春权, 孟宪民, 张晓光, 夏丹丹. 陆地生态系统碳收支/碳平衡研究进展. 资源开发与市场, 2009, 25(2): 165–171. DOI:10.3969/j.issn.1005-8141.2009.02.021 |
| [21] | Song B, Niu S L, Luo R S, Luo Y Q, Chen J Q, Yu G R, Olejnik J, Wohlfahrt G, Kiely G, Noormets A, Montagnani L, Cescatti A, Magliulo V, Law B E, Lund M, Varlagin A, Raschi A, Peichl M, Nilsson M B, Merbold L. Divergent apparent temperature sensitivity of terrestrial ecosystem respiration. Journal of Plant Ecology, 2014, 7(5): 419–428. DOI:10.1093/jpe/rtu014 |
| [22] | Zhou T, Shi P J, Hui D F, Luo Y Q. Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implications for carbon-climate feedback. Journal of Geophysical Research, 2009, 114(G2): G02016. |
| [23] | 徐满厚, 薛娴. 气候变暖对陆地植被-土壤生态系统的影响研究. 生命科学, 2012, 24(5): 492–500. |