 2018, Vol. 38
2018, Vol. 38文章信息
- 张树斌, 王襄平, 吴鹏, 孙晗, 李巧燕, 吴玉莲, 韩威, 武娴.
- ZHANG Shubin, WANG Xiangping, WU Peng, SUN Han, LI Qiaoyan, WU Yulian, HAN Wei, WU Xian.
- 吉林灌木群落物种多样性与气候和局域环境因子的关系
- Relationship between shrub species diversity and climate and local environmental factors across Jilin
- 生态学报. 2018, 38(22): 7990-8000
- Acta Ecologica Sinica. 2018, 38(22): 7990-8000
- http://dx.doi.org/10.5846/stxb201801230178
-
文章历史
- 收稿日期: 2018-01-23
- 网络出版日期: 2018-08-24





很多研究发现物种多样性从低纬度到高纬度、从低海拔到高海拔而逐渐减少的分布规律, 其成因一直吸引着生态学家和生物地理学家的兴趣[1-2]。过去的几十年中, 研究者们提出了很多假说, 包括现代气候、地史成因、几何限制假说等几类解释机制[3-10]。其中, 以气候为基础的几种假说得到了最为广泛的支持, 因此一些研究认为气候因素可以作为预测物种多样性分布格局的首要因子[11]。
目前多数的生物多样性地理格局的研究主要是探讨气候因素对于物种多样性的影响, 而对局域环境因子的影响的研究关注相对较少[12-14]。然而, 一些关于气候与局域环境相对影响的研究发现, 物种多样性格局不仅受到气候变量的影响, 同时还受到冠层盖度、群落结构、人为干扰等局域环境因子的影响[15-16]。如在森林群落中对不同生活型多样性的研究发现, 气候变量虽然对于木本植物具有较强的解释力, 但局域环境因子对于草本植物的解释力显著高于气候因子[17]。因此, 气候和局域因素的相对作用大小, 是随群落层次或生活型的不同而异的。在灌木群落中, 虽然有一些研究探讨了物种丰富度和气候梯度的关系[18], 但关于局域环境因子和气候的相对作用大小的研究还很少。这可能是由于灌丛的群落结构相对于森林来说要简单很多, 因此群落结构等局域因素对灌丛多样性的影响受到的重视不够。
生物多样性由物种丰富度和均匀度两方面组成, 分别代表着群落中的物种数以及不同物种间多度分布的均匀程度[19]。物种丰富度一般从低纬度到高纬度、从低海拔到高海拔而逐渐减少, 而对于均匀度来说, 不同研究发现的格局差异很大[19-21]。目前大多数关于生物多样性的研究都侧重于探究物种丰富度的分布格局及影响因素, 而关于均匀度分布格局的研究较少[22]。均匀度地理格局的解释机制也只是在少数研究中被提及, 而且这些解释机制往往也只考虑了气候因素对于物种均匀度的影响, 并没有将局域环境的作用考虑进去[23-24]。本文将对两个关于均匀度地理格局的假说进行验证。一种假说认为(假说1), 群落均匀度应当随纬度的增加而增加;因为在热带生产力较高使得种间竞争加剧, 导致均匀度降低[20]。但这一假说存在一定问题, 因为较高的生产力未必导致较强的种间竞争。与此相反, 在我国森林群落中的研究发现, 群落均匀度与温度正相关, 即应随纬度的增加而下降[21]。我们认为, 这一现象与北方森林多为单优群落、而南方森林群落物种较多但优势种常不明显的事实一致。这说明群落均匀度应当随着环境条件的改善而增加(假说2), 本文将利用灌木群落的数据中对此进行检验。
目前对于灌木群落多样性格局的研究主要集中在干旱区的地带性灌丛植被[18, 25]。本文的研究区域处于我国半湿润气候区的吉林省, 这里的灌木群落为森林破坏后形成、人为干扰的影响较大, 与干旱区地带性的灌丛在起源、物种组成、群落结构上有很大差异。关于这种半湿润地区次生灌丛的多样性地理格局, 相关研究还很少。而本文将探究在这种次生灌木群落中, 物种多样性的地理格局有什么特点?其解释机制是什么?
本研究对吉林省东部、南部的45块灌木群落样地进行了调查, 主要研究目的在于:1)探究气候因子以及局域环境对物种多样性的相对作用大小;2)这些因素对不同水分生态型灌木丰富度的影响有什么差异?3)物种丰富度和均匀度的影响因素有什么差异?
1 研究方法 1.1 研究区概况研究区位于吉林省东部和南部, 调查样地经纬度范围为125.5—129.0°E、41.6—44.0°N, 海拔范围为600—1327 m(图 1)。本研究区处于湿润、半湿润气候区, 沿纬度、经度设置样线, 以覆盖研究区的温度、水分梯度, 以研究气候、局域因子对灌丛群落的作用。大部分地区年平均气温为3—5 ℃, 最冷月均温在-15.2—19.2 ℃之间, 平均年活动积温2700—3600 ℃, 年降水量550—910 mm。

|
| 图 1 灌丛群落采样样地位置图 Fig. 1 Locations of the plots sampled in the shrub communities across the east and south part of Jilin Province, northeast China |
本研究区域的灌丛主要为森林破坏后形成的次生灌丛, 并经历了不同程度的人为干扰。在调查的45块样方中, 实际调查到灌木植物88种, 草本植物320种, 并依据灌丛群落的优势种对灌丛类型进行划分命名, 共调查到灌丛类型有平榛灌丛、平榛-毛榛灌丛、柳-绣线菊灌丛、胡枝子灌丛、金露梅灌丛、山刺玫-平榛灌丛、柳灌丛和平榛-胡枝子灌丛等共计24种。
1.2 样地调查在吉林安图、敦化、蛟河、通化、抚松等地选取具有代表性的灌丛群落类型进行调查, 最终共设置45个10 m×10 m灌丛样方。群落调查按照方精云等[26]制定的技术规范执行, 记录样地的地理坐标, 海拔、坡度、坡向(记录为N、NE、E、SE、S、SW、W、NW等)以及样地干扰程度(轻微、中度、重度)、土壤水分状况(干燥、湿润、中度)等指标, 并分为灌木、草本两层进行调查。对于灌木层调查, 将灌丛样方划分为4个5 m×5 m的灌木样方, 记录每个灌木样方内出现的全部灌木种的种名、平均高、株数和盖度等信息。对于草本层, 我们在每个灌木样地的四角设置4个1 m× 1 m样方, 然后记录每个样方中每个物种的物种名称、盖度、平均高度和多度。
1.3 气候数据目前一般认为, 影响物种多样性的主要气候要素有[8, 17]:1)能量和水分;水分-能量动态假说(water-energy dynamic hypothesis)认为物种丰富度的地理格局是受水分和能量共同作用的结果。2)冬季低温;寒冷忍耐假说(freezing tolerance hypothesis)认为, 在寒冷地区很多物种由于不能忍受冬季的寒冷而无法生存, 因此, 随着冬季温度的降低物种多样性逐渐减少。研究这些气候要素的作用、验证这些气候假说对我们理解物种多样性分布格局具有重要的意义。
为此, 对相关气候数据进行提取和计算。根据调查样地的地理坐标, 利用ArcGIS 10.0中从全球高分辨率气候数据库中提取各样地1—12月均温与降水量数据(1950—2000年平均值), 并计算下述指标:(1)最冷月均温MTCM (mean temperature of the coldest month, ℃), 即冬季最冷月份的平均气温, 反映冬季低温;(2)年均降水量MAP (mean annual precipitation, mm), 反映水分的有效性(water availability);(3)潜在蒸散量PET (potential evapotranspiration, mm), 是植被-气候关系分析重要的气候指标, 反映能量有效性(energy availability), 计算方法见方精云等[27]。这些气候指标的选取主要是根据热量积累、水分供给、低温限制等影响因子对植物生理活动的限制作用, 从而可能导致物种多样性的变化而确定的[27-28]。
1.4 数据分析本文研究了如下多样性指标和环境因子的关系:群落(灌木+草本)、灌木层、草本层的物种丰富度, 及Pielou均匀度指数(Evnenness)。
物种丰富度计算:S=出现在样方内的物种数
灌木层均匀度计算:每个地点灌丛样地是由4个5 m×5 m的灌木样方组成, 将每个灌木样方中各个物种的盖度相加求和, 而后对4个灌木样方中各个物种的盖度和进行平均, 作为每个灌木物种的重要值, 随后运用Pielou指数公式计算出均匀度。
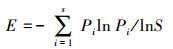
|
式中, Pi为种i的重要值, S为出现在样方内的物种数。
草本层均匀度计算:每个5 m×5 m的灌木样方中有4个1 m× 1 m的草本样方, 将每个物种的盖度相加, 除以草本总样地数, 而作为每个草本物种的重要值, 而后进行均匀度计算。
群落均匀度计算:将草本物种重要值和灌木重要值, 通过乘或除一定面积进行标准化, 而得到样地群落的物种重要值, 而后进行均匀度计算。
同时, 我们还对不同水分生态型灌木种的物种丰富度与环境的关系进行了分析以比较其差异。有研究表明, 不同水分生态型与气候因子的关系可能相反, 对灌丛多样性格局有重要影响[18]。灌木物种的水分生态型根据《东北植物志》划分为湿中生、中生、旱中生。由于只是将不同水分生态型的灌木种数进行分类统计, 作为不同水分生态型进行分析, 划分后导致不少样地中有的水分生态型只有1个物种, 因此灌木不同水分生态型的比较分析没有计算均匀度、而只针对物种丰富度。
首先采用单因素分析研究物种多样性和各气候、局域环境指标之间的关系。由于单因素分析表明气候指标并不能很好的解释研究区灌丛的多样性地理格局, 所以运用一般线性模型分析以下两类变量对物种丰富度和均匀度指数的相对影响大小:(1)气候指标:PET、MTCM、MAP;(2)局域环境指标:坡度、坡向、土壤水分状况、灌木层盖度、灌丛高度、人为干扰程度。对这些指标采用基于AIC (Akaike Information Criterion)的模型选择方法进行变量筛选, 以研究影响灌木群落物种多样性的主要因子。
数据分析利用R软件进行[29]。
2 结果 2.1 灌木群落物种多样性的地理格局本研究中, 群落物种丰富度随纬度和经度的增加而增加, 均匀度随纬度和经度没有明显的变化(图 2)。对于灌木层来说, 物种丰富度和均匀度指数随纬度和经度都没有显著的变化;对于草本层, 物种丰富度和均匀度指数都随纬度和经度的增加而增加。

|
| 图 2 样地物种丰富度、均匀度随纬度、经度的变化 Fig. 2 Species richness, evenness changes with latitude, longitude 实线、点线分别为群落、草本的回归线, 图中给出的回归线均在P < 0.05水平显著 |
与气候因子的相关分析表明, 群落物种丰富度与降水(MAP)呈显著负相关关系, 而与冬季低温(MTCM)、能量(PET)没有显著的相关性;灌木物种丰富度、均匀度与3个气候指标都没有显著的相关性;而草本的物种丰富度、均匀度指数与MAP均呈显著负相关关系, 而与MTCM、PET没有显著关系(表 1、图 3)。
| 物种多样性指数 Species diversity index |
单变量解释力Univariate explanatory power/% | ||||||||
| MTCM | MAP | PET | 坡度 Slope |
灌木层层高 Shrub layer height |
灌木层盖度 Shrub layer coverage |
坡向 Aspect |
干扰程度 Disturb degree |
水分状况 Water condition |
|
| 物种丰富度Species richness | |||||||||
| 群落Community | -3.0 | -26.0*** | +3.6 | +0.4 | +0.8 | -11.0* | 14 | 26.2** | 26.93** |
| 灌木Shrub | -0.6 | +2.0 | -3.0 | -2.0 | -0.4 | -0.2 | 8 | 1.0 | 2.0 |
| 草本Herb | -4.0 | -36.0* | +6.0 | +1.0 | +1.7 | -9.3* | 22 | 26.3** | 30.62** |
| 均匀度指数Evenness | |||||||||
| 群落Community | -0.09 | +6.0 | -7.0 | -1.0 | -2.0 | -2.0 | 10.9 | 3.4 | 4.0 |
| 灌木Shrub | +4.0 | +0.8 | +0.004 | +1.3 | -0.8 | -3.0 | 16.8 | 8.5 | 3.5 |
| 草本Herb | -6.0 | -10.2* | +0.04 | -1.9 | +1.3 | -15.92** | 10.3 | 20.6** | 13.2 |
| 灌木不同水分生态型物种数Number of different water ecological species of shrub | |||||||||
| 湿中生Phreatophyte- | +0.04 | +25.0** | -14.0* | -4.0 | +0.3 | +0.1 | 13.4 | 2.0 | 1.8 |
| 中生Mesoxero- | -2.3 | -1.2 | -0.04 | -0.9 | -2.5 | -1.0 | 6 | 2.0 | 3.7 |
| 旱中生Xero- | +0.3 | -8.4 | +16.0** | +5.8 | -0.3 | +0.01 | 4 | 4.3 | 1.5 |
| P < 0.1; *P < 0.05; **P < 0.01; ***P < 0.001;前标“-”为负相关关系, “+”为正相关关系。未标者为类型变量;MTCM:最冷月均温, mean temperature of the coldest month;MAP:年均降水量, mean annual precipitation;PET:潜在蒸散量, potential evapotranspiration | |||||||||

|
| 图 3 物种丰富度和均匀度指数与年降水量(MAP)、潜在蒸散量(PET)、最冷月低温(MTCM)等气候因子的关系 Fig. 3 Relationship between species richness, evenness index and climate factors such as annual precipitation (MAP), potential evapotranspiration (PET), coldest month low temperature (MTCM) and so on 实线、点线分别为群落、草本的回归线, 图中给出的回归线均在P < 0.05水平显著 |
对局域环境因子的相关分析表明, 群落物种丰富度与灌木层盖度呈显著负相关关系, 与土壤水分状况和干扰程度呈显著正相关关系, 与坡度、灌木层层高、坡向等局域环境指标都没有显著的相关性;群落均匀度指数、灌木物种丰富度、均匀度指数与6个局域环境指标都没有显著的相关性;草本的物种丰富度只与干扰程度和土壤水分状况呈显著的正相关关系, 与其他几个局域环境指标都没有显著的相关性, 而草本的均匀度指数只与灌木层盖度呈显著的负相关关系, 与干扰程度呈显著的正相关关系(表 1、图 3)。
AIC模型筛选的结果表明(表 2), 对于群落和草本物种丰富度来说, 气候没有显著解释力, 只有灌木层盖度、干扰程度以及土壤水分状况等局域环境因子进入最终的解释模型, 分别解释了44%—47%的变异。灌木层物种丰富度, 则没有变量可以显著解释(见讨论)。
| 物种多样性指数 Species diversity index |
保留变量(解释百分率%) Reserved variables (Explanations%) |
模型解释力Model explanatory force/% |
| 物种丰富度Species richness | ||
| 群落Community | 灌木层盖度(-10.6), 干扰程度(10.2), 土壤水分状况(23.3) | 44.1 |
| 灌木层Shrub | 无 | 0 |
| 草本层Herb | 灌木层盖度(-9.4), 干扰程度(10.1), 土壤水分状况(27.2) | 46.7 |
| 均匀度指数Evenness | ||
| 群落Community | MAP(+9.7), 灌木层盖度(-6.0) | 13.7 |
| 灌木层Shrub | MTCM(+13.7), 土壤水分状况(3.47) | 17.2 |
| 草本层Herb | 灌木层盖度(-15.9), 干扰程度(12.83) | 28.7 |
| 1)前标“-”为负作用, “+”为正作用。未标者为类型变量;初始模型包含了3个气候因子(PET、MAP和MTCM)以及6个局域环境因子(灌木层盖度、土壤水分状况、人为干扰程度等, 见研究方法)。对于保留在模型中的变量, 括号中给出了其解释力;对于数值变量, 给出了其作用方向(+或-) | ||
对于群落和灌木层均匀度, 气候和局域环境因子都进入了模型, 但气候因子(MAP或MTCM)的解释力大于后者。对于草本层均匀度, 气候因子则没有显著解释力, 局域环境因子灌木层盖度和干扰程度分别解释了15.9%和12.8%的变异(表 2)。
2.3 不同水分生态型灌木物种丰富度的差异把灌木划分为不同的水分生态型分别进行分析, 湿中生灌木种的物种丰富度与MAP显著正相关、与PET负相关, 而旱中生灌木种的物种丰富度与PET显著正相关, 中生种则与MAP、PET和MTCM都没有显著相关性(图 4; 表 1)。逐步回归分析则表明(表 3), MAP、PET分别是影响湿中生、旱中生灌木物种丰富度的主要因素, 分别解释了20%和16.2%的变异。中生灌木种的物种丰富度, 则无变量可解释。

|
| 图 4 不同水分生态型灌木的物种丰富度与年降水量(MAP)、潜在蒸散量(PET)、最冷月低温(MTCM)的关系 Fig. 4 Relationship between species richness and annual precipitation (MAP), potential evapotranspiration (PET), coldest month low temperature (MTCM) for different water ecotypes 实线、点线分别为湿中生、旱中生的回归线, 图中给出的回归线均在P < 0.05水平显著 |
| 物种多样性指数 Species diversity index |
保留变量(解释百分率%) Reserved variables (Explanations%) |
模型解释力 Model explanatory force/% |
| 灌木不同水分生态型物种数Number of different water ecological species of shrub | ||
| 湿中生灌木Phreatophyte- | MAP(+20) | 20.0 |
| 中生灌木Mesoxero- | — | — |
| 旱中生灌木Xero- | PET(+16.2) | 16.2 |
| 前标“-”为负作用, “+”为正作用。未标者为类型变量; 初始模型包含了3个气候因子(PET、MAP和MTCM)以及6个局域环境因子(灌木层盖度、土壤水分状况、人为干扰程度等, 见研究方法); 对于保留在模型中的变量, 括号中给出了其解释力;对于数值变量, 给出了其作用方向(+或-) | ||
以往的研究表明, 气候因素与物种多样性分布格局有着密切的关系。在众多的气候因素中, 热量、水分和冬季低温是决定大尺度上多样性格局的主导因子[8]。在低纬度的地区, 热量条件较高, 水分条件也较好, 往往其物种多样性也就越丰富[30-31], 而在本研究区中, 灌木群落和草本的物种丰富度在低纬度地区反而较小, 并与水分呈负相关关系, 与热量及冬季低温都没有显著关系(图 2、3)。这些都与以往研究的结果有着明显的差异。
本研究中(表 2), 对于群落和草本层的物种丰富度来说, 气候因子并没有进入物种丰富度的解释模型, 只有灌木层盖度、干扰程度以及土壤水分状况等局域环境因子进入最终的模型。进一步对局域环境因子进行的分析表明, 灌木层盖度与MAP正相关, 而群落和草本的物种丰富度与灌木层盖度负相关(图 5)。这说明随着降水的增多灌木层盖度增大, 草本层由于接受到的光照下降因而生产力降低, 从而导致草本层的丰富度降低[32]。在本研究区, 降水从西南向东北减少, 由于灌木层盖度随降水的变化, 从而形成了草本物种丰富度随纬度上升而上升的独特格局。灌木群落中草本的物种数要远高于灌木层, 群落物种丰富度大小主要是由草本决定的, 因此其地理格局、主要影响因子都和草本层相似。以往的研究多注重分析物种丰富度和气候关系, 而对局域环境因子重视不够。本文的结果表明, 局域环境因子如灌木层盖度、干扰程度以及土壤水分状况对多样性格局起主要作用, 图 2、3中群落和草本的物种丰富度与纬度、MAP之间的关系与一般的研究相反, 充分说明了局域环境因子的重要性:气候因子是通过灌木层盖度间接影响草本层多样性格局的。因此, 简单分析多样性和气候之间的相关关系, 有可能掩盖气候对多样性格局的真实影响机制。

|
| 图 5 灌木层盖度与年降水量(MAP)和物种丰富度、均匀度指数的关系 Fig. 5 Relationship between shrub layer coverage and annual precipitation (MAP) and species richness and evenness index 图a中实线为灌木层盖度和MAP (mm)的回归线;图b和图c中实线和点线分别为群落和草本的回归线, 图中给出的回归线均在P < 0.05水平显著 |
本文中灌木物种丰富度与气候没有显著的关系, 与以往对内蒙灌丛的研究一致[18]。但是, 这并不意味着气候对灌木物种丰富度没有影响。对不同水分生态型灌木的分析表明(表 3, 图 4), 湿中生灌木丰富度主要受降水影响, 随降水增加而上升, 并随PET上升而下降。这种物种丰富度与降水和能量关系相反的现象, 是多样性受水分限制的典型表现, 也常见于其他干旱到半湿润区的研究中[8, 18, 28]。这是因为湿中生灌木对水分要求相对较高, 而能量上升导致蒸散增加, 从而使得水分有效性降低、物种丰富度下降。与此不同, 旱中生灌木由于对水分要求不高, 因此其物种数随PET的增加而上升。由此可见, 不同水分生态型对气候梯度的响应方向不同甚至相反(图 4), 是导致灌木的总物种丰富度与气候没有显著相关关系的一个重要原因。代爽等[18]在对内蒙干旱灌丛的研究中也发现了相似的结果, 并认为这反映了物种的生态特性以及种间相互作用是在气候因子之外影响多样性地理格局的重要因素。本研究的不同水分生态型灌木的差异支持该观点。上述研究结果再次表明, 简单地分析气候因子和总物种数的相关性, 还可能会掩盖植物功能型对气候响应的差异、得到不适当的结论。
3.2 物种均匀度与气候因子以及局域环境的关系在以往的研究中, 对于均匀度地理分布格局的研究相对较少, 然而均匀度作为反映生物多样性变化的一个方面, 对于生态系统功能具有重要的影响。比如, 一些研究发现较高的均匀度可促进群落生产力[19-20, 23]。这里我们利用本研究的调查数据, 对引言中的两个假说进行了检验, 以进一步理解群落均匀度地理变化的影响机制。
本研究中, 草本层均匀度与纬度显著正相关(图 2)。表面来看, 这支持假说1, 即均匀度应随纬度的增加而增加[20]。但多元分析的结果表明, 在考虑了局域环境因子后, 群落和草本层均匀度都随降水或温度的上升而上升(见表 2中变量正负号)。这一结果支持假说2, 即均匀度在温度、水分条件较好的地方较高, 也与Qiao等[21]在中国纬度梯度上的森林群落中得到的结果一致。至于草本层均匀度, 表 2表明局域环境因子是其主要影响因子, 气候因子则没有显著的解释力。而且, 草本层均匀度随灌木层盖度的增加而下降。因此, 图 2中草本层均匀度与纬度的显著正相关, 也是由于灌木层盖度随水分增加而增加, 导致草本层的光照减少、进而导致草本层均匀度的下降(图 5)。因此, 虽然本研究中草本均匀度和群落、灌木层均匀度主要的影响因子有明显不同(后二者主要受降水和温度的影响), 但本质上的影响机制是一致的, 即均匀度在环境条件较好、生产力较高的条件下较高。
因此, 本文结论支持假说2, 而并不支持假说1所阐述的影响机制。假说1的问题在于, 较高的生产力未必导致较强的种间竞争。这是因为, 虽然生产力较高的地方往往物种较多, 但较高的生产力也可提供更多的资源以供不同的物种之间分配。如很多研究表明, 热带森林的群落空间结构要比北方的森林复杂的多, 即可以使得更多的物种通过生态位分化(避免竞争)而共存。生态位分化是物种间减少竞争的有效机制, 许多研究认为这是高生产力地区能维持较高的生物多样性的重要原因[33-36]。不过, 关于这两个均匀度的假说还需要进一步的检验, 因为确实也有不少研究发现均匀度随纬度增加而上升或无显著变化, 关于均匀度的地理格局和成因还有着很大的争议[19-20]。搞清不同研究之间为什么会有这些差异, 对于理解生物多样性的形成机制, 有着重要的意义。
4 结论本文通过对半湿润区次生灌木群落的物种多样性进行调查, 研究结果表明:
1) 吉林灌丛物种丰富度主要受局域环境因子影响, 草本丰富度与纬度、气候(MAP)的关系与以往研究相反, 是由于灌木盖度随水分增加而上升, 导致草本层物种数下降。而灌木层总丰富度与气候无相关关系, 是因为不同水分生态型的灌木对气候梯度响应相反, 反映出功能型对多样性格局的重要影响。
2) 草本层均匀度随纬度增加而上升, 与降水负相关, 是因为气候通过对灌木层盖度的作用, 导致草本层生产力下降从而使均匀度降低;对于灌木群落和灌木层均匀度而言, 气候有着较大的影响。但不管是群落, 还是灌木、草本层均匀度, 都支持随着环境条件的改善均匀度增加的假说。
3) 本文的结果反映了气候对多样性格局影响机制的复杂性。除了气候条件本身对多样性的影响外, 气候还可通过局域生物因素(如盖度)、及物种性状(如水分生态型)作用于多样性格局, 这种间接作用在以往的气候-多样性关系的研究中没有得到足够的重视。由于局域生物因素也随气候而变化, 仅研究多样性和气候的表面关系, 是无法准确预测气候变化对多样性的影响的。
| [1] | Hillebrand H. On the generality of the latitudinal diversity gradient. The American Naturalist, 2004, 163(2): 192–211. DOI:10.1086/381004 |
| [2] | Renner S S, Lomolino M V, Riddle B R, Brown J H. Biogeography, third edition. Systematic Biology, 2006, 55(4): 696–698. DOI:10.1080/10635150600899764 |
| [3] | Brown J H, Gillooly J F, Allen A P, Savage V M, West G B. Response to forum commentary on "toward a metabolic theory of ecology". Ecology, 2004, 85(7): 1818–1821. DOI:10.1890/03-0800 |
| [4] | Wiens J J, Donoghue M J. Historical biogeography, ecology and species richness. Trends in Ecology & Evolution, 2004, 19(12): 639–644. |
| [5] | Storch D, Davies R G, Zajiček S, Orme C D L, Olson V, Thomas G H, Ding T, Rasmussen P C, Ridgely R S, Bennett P M, Blackburn T M, Owens I P F, Gaston K J. Energy, range dynamics and global species richness patterns:reconciling mid-domain effects and environmental determinants of avian diversity. Ecology Letters, 2006, 9(12): 1308–1320. DOI:10.1111/ele.2006.9.issue-12 |
| [6] | McCain C M. Global analysis of bird elevational diversity. Global Ecology and Biogeography, 2009, 18(3): 346–360. DOI:10.1111/geb.2009.18.issue-3 |
| [7] | Rangel T F L V B, Diniz-Filho J A F. Neutral community dynamics, the mid-domain effect and spatial patterns in species richness. Ecology Letters, 2005, 8(8): 783–790. DOI:10.1111/ele.2005.8.issue-8 |
| [8] | 王志恒, 唐志尧, 方精云. 物种多样性地理格局的能量假说. 生物多样性, 2009, 17(6): 613–624. |
| [9] | 唐志尧, 王志恒, 方精云. 生物多样性分布格局的地史成因假说. 生物多样性, 2009, 17(6): 635–643. |
| [10] | 王襄平, 方精云, 唐志尧. 中域效应假说:模型、证据和局限性. 生物多样性, 2009, 17(6): 568–578. |
| [11] | Kerr J T, Currie D J. The relative importance of evolutionary and environmental controls on broad-scale patterns of species richness in North America. Écoscience, 1999, 6(3): 329–337. DOI:10.1080/11956860.1999.11682546 |
| [12] | Goetz S J, Sun M, Zolkos S, Hansen A, Dubayah R. The relative importance of climate and vegetation properties on patterns of North American breeding bird species richness. Environmental Research Letters, 2014, 9(3): 034013. DOI:10.1088/1748-9326/9/3/034013 |
| [13] | Marini L, Nascimbene J, Nimis P L. Large-scale patterns of epiphytic lichen species richness:photobiont-dependent response to climate and forest structure. Science of The Total Environment, 2011, 409(20): 4381–4386. DOI:10.1016/j.scitotenv.2011.07.010 |
| [14] | White E P, Hurlbert A H. The combined influence of the local environment and regional enrichment on bird species richness. The American Naturalist, 2010, 175(2): E35–E43. DOI:10.1086/649578 |
| [15] | Laughlin D C, Bakker J D, Fulé P Z. Understorey plant community structure in lower montane and subalpine forests, Grand Canyon National Park, USA. Journal of Biogeography, 2005, 32(12): 2083–2102. DOI:10.1111/jbi.2005.32.issue-12 |
| [16] | Li Q, Yang X, Soininen J, Chu C J, Zhang J Q, Yu K L, Wang G. Relative importance of spatial processes and environmental factors in shaping alpine meadow communities. Journal of Plant Ecology, 2011, 4(4): 249–258. DOI:10.1093/jpe/rtq034 |
| [17] | Wang X P, Fang J Y, Sanders N J, White P S, Tang Z Y. Relative importance of climate vs local factors in shaping the regional patterns of forest plant richness across northeast China. Ecography, 2009, 32(1): 133–142. DOI:10.1111/eco.2009.32.issue-1 |
| [18] | 代爽, 王襄平, 刘超, 武娴, 李巧燕, 王敏. 内蒙古灌木群落物种丰富度与气候的关系. 北京大学学报:自然科学版, 2013, 49(4): 689–698. |
| [19] | Willig M R, Kaufman D M, Stevens R D. Latitudinal gradients of biodiversity:pattern, process, scale, and synthesis. Annual Review of Ecology, Evolution, and Systematics, 2003, 34: 273–309. DOI:10.1146/annurev.ecolsys.34.012103.144032 |
| [20] | Soininen J, Passy S, Hillebrand H. The relationship between species richness and evenness:a meta-analysis of studies across aquatic ecosystems. Oecologia, 2012, 169(3): 803–809. DOI:10.1007/s00442-011-2236-1 |
| [21] | Qiao X J, Tang Z Y, Shen Z H, Fang J Y. What causes geographical variation in the species-area relationships? A test from forests in China. Ecography, 2012, 35(12): 1110–1116. DOI:10.1111/j.1600-0587.2011.06869.x |
| [22] | Hillebrand H, Bennett D M, Cadotte M W. Consequences of dominance:a review of evenness effects on local and regional ecosystem processes. Ecology, 2008, 89(6): 1510–1520. DOI:10.1890/07-1053.1 |
| [23] | Jost L. The relation between evenness and diversity. Diversity, 2010, 2(2): 207–232. DOI:10.3390/d2020207 |
| [24] | Maestre F T, Castillo-Monroy A P, Bowker M A, Ochoa-Hueso R. Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. Journal of Ecology, 2012, 100(2): 317–330. DOI:10.1111/jec.2012.100.issue-2 |
| [25] | 冶民生, 关文彬, 谭辉, 马克明, 刘国华, 汪西林. 岷江干旱河谷灌丛α多样性分析. 生态学报, 2004, 24(6): 1123–1130. DOI:10.3321/j.issn:1000-0933.2004.06.005 |
| [26] | 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533–548. |
| [27] | 方精云. 全球生态学:气候变化与生态响应. 北京: 高等教育出版社, 2000. |
| [28] | Hawkins B A, Porter E E. Water-energy balance and the geographic pattern of species richness of western Palearctic butterflies. Ecological Entomology, 2003, 28(6): 678–686. DOI:10.1111/een.2003.28.issue-6 |
| [29] | Bivand R S, Pebesma E J, Gomez-Rubio V.空间数据分析与R语言实践.徐爱萍, 舒红, 译.北京: 清华大学出版社, 2013. |
| [30] | Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220–227. DOI:10.1038/35012228 |
| [31] | Clarke A, Gaston K J. Climate, energy and diversity. Proceedings of the Royal Society B:Biological Sciences, 2006, 273(1599): 2257–2266. DOI:10.1098/rspb.2006.3545 |
| [32] | Evans K L, James N A, Gaston K J. Abundance, species richness and energy availability in the North American avifauna. Global Ecology and Biogeography, 2006, 15(4): 372–385. DOI:10.1111/geb.2006.15.issue-4 |
| [33] | Colwell R K, Rahbek C, Gotelli N J. The mid-domain effect and species richness patterns:what have we learned so far?. The American Naturalist, 2004, 163(3): E1–E23. DOI:10.1086/382056 |
| [34] | Fernández-Palacios J M, de Nicolas J P. Altitudinal pattern of vegetation variation on Tenerife. Journal of Vegetation Science, 1995, 6(2): 183–190. DOI:10.2307/3236213 |
| [35] | Sang W G. Plant diversity patterns and their relationships with soil and climatic factors along an altitudinal gradient in the middle Tianshan Mountain area, Xinjiang, China. Ecological Research, 2009, 24(2): 303–314. DOI:10.1007/s11284-008-0507-z |
| [36] | Shah D N, Domisch S, Pauls S U, Haase P, Jäehnig S C. Current and future latitudinal gradients in stream macroinvertebrate richness across North America. Freshwater Science, 2014, 33(4): 1136–1147. DOI:10.1086/678492 |