 2018, Vol. 38
2018, Vol. 38文章信息
- 孙立群, 李晴岚, 陈骥, 牛俊.
- SUN Liqun, LI Qinglan, CHEN Ji, NIU Jun.
- 欧亚大陆不同生态区植被生长对降水响应的季节变化规律
- Seasonal changes in vegetation response to precipitation across eco-regions over Eurasia
- 生态学报. 2018, 38(22): 8051-8059
- Acta Ecologica Sinica. 2018, 38(22): 8051-8059
- http://dx.doi.org/10.5846/stxb201801080050
-
文章历史
- 收稿日期: 2018-01-08
- 网络出版日期: 2018-08-24





2. 香港大学土木工程系, 香港 999077;
3. 中国农业大学中国农业水问题研究中心, 北京 100083
2. Department of Civil Engineering, The University of Hongkong, HongKong 999077, China;
3. Center for Agricultural Water Research in China, China Agricultural University, Beijing 100083, China
欧亚大陆土地广袤, 涵盖了我国“一带一路”战略规划的大部分国家。欧亚大陆人口众多, 不同国家和地区的地理条件、经济发展差异极大[1]。类似于中国的“胡焕庸线”难题, 水资源对欧亚大陆很多国家, 尤其是中亚干旱地区国家的生态安全和经济发展具有重要的意义[2]。了解欧亚大陆植被生长对降水响应的时空格局是“一带一路”战略顺利实施的科学与生态基础。在气候变化的背景下, 如何因地制宜地应对及适应气候变化, 从而促进当地社会与经济的可持续性发展, 是欧亚大陆每一个国家和地区的人们所需要面对和亟待解决的问题[3]。
植被生态系统作为陆地生态系统的主体, 在不同地理环境和气象条件下对气候变化有着不同响应机制。例如, 在中国的内蒙古地区, 植被生长受水分影响多一些[4]。而在中国东北地区, 则对温度变化更为敏感[5]。植被生长是一个极为复杂的过程, 在排除霜冻、病虫害、水利灌溉和施肥等影响后, 植被生长主要受到温度、水分条件、日照辐射3个自然条件的胁迫[6]。选取合适的方法, 分析陆地植被生长在不同地区对降水变化的响应关系是本文研究的重点。
经过数十年遥感数据的积累, 在大陆尺度研究每个像元上植被生长与气候变化的关系已经成为可能。Peng等[7]使用归一化植被指数NDVI(Normalized Difference Vegetation Index)和全球气象格点数据, 采用偏相关方法(Partial Correlation Coefficient)单独计算了日间和夜间增温对北半球高纬度地区植被生态系统的影响, 发现湿润地区和干旱地区的植被对昼夜增温的响应是不一致的。Shen等[8]使用SPOT(Satellite Pour l′Observation de la Terre)NDVI分析了青藏高原上降水对春季植被物候的影响, 指出干旱地区的植被为了最大限度地利用降水来促进自身的生长, 会比湿润地区的植被表现得对降水更为敏感。因此, 本文将以生态区为视角, 使用NDVI植被指数表征植被生长状态, 分析欧亚大陆植被生长对降水的响应关系在不同生态区的季节差异和该响应关系的变化趋势, 从而分析得到欧亚大陆植被生长对降水响应的时空格局, 为欧亚大陆不同区域应对和适应气候变化提供决策依据。
1 数据和方法本文以欧亚大陆为研究区域, 以陆域植被生态系统为研究对象, 利用GIMMS(Global Inventory Modeling and Mapping Studies)长序列(1982—2015年)第3代植被指数NDVI3g数据来表征陆地生态系统的植被生长状况。NDVI3g采用多颗AVHRR(Advanced Very High Resolution Radiometer)卫星数据计算得到, 数据的空间分辨率为0.0833度, 每月的前15天产生一个a值, 余下的天数的数据产生第二个b值, 在数据发布时已经做了初步的大气校正[9]。为了最大限度去除云的干扰, 在计算月度数据时, 本研究将取每个月a、b两值的最大值作为当月的NDVI值[10-11]。相关研究区域的气象数据来自英国东安哥拉大学气候研究中心发布的空间分辨率为0.5度的全球网格数据集Climatic Research Unit Time-series data sets第4版本(CRUTS V4)[12]。
根据欧亚大陆的地理位置, 参考已有研究成果对季节的划分方案[13], 本文设置4至5月为春季, 6至8月为夏季, 9至10月为秋季, 从而将月值植被数据和气象数据整合生成春、夏、秋3个季节的均值。对季节平均NDVI值小于0.1的像素设置为无植被, 将每个季节的NDVI和气象数据做去趋势化处理[13], 再将气象数据从0.5度重新采样至与NDVI数据保持一致的空间分辨率(1/12度), 参与其后的相关性计算[7]。最后使用世界野生动物基金会(World Wildlife Fund, WWF)发布的全球陆地生态区划图[14], 对植被-降水关系在每个生态区上进行分异研究。WWF发布的生态区划地图是目前学界使用最为广泛的区划方案[15], 在本文研究的欧亚大陆范围内, 共有328个生态区(eco-region)。
为了单独研究区域内植被生长对降水响应的时空变化规律, 本文通过计算偏相关系数(Partial Correlation Coefficient)[16], 排除温度和日照辐射的共同作用[6]。例如, 在计算某像素点上的植被生长对降水的响应关系, 可通过计算二阶偏相关系数, 单独计算植被指数(NDVI)与降水(Precipitation)的偏相关系数(RNDVI-Prep), 并排除平均气温和日照辐射的共同作用, 其计算公式为:
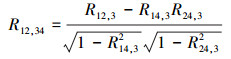
|
(1) |
式(1)中, 4个计算变量分别为:1=植被指数(NDVI);2=降水;3=平均温度;4 =日照辐射(使用云量数据替代)。式(1)中的R12, 3是排除平均温度共同作用之后计算得到的NDVI与降水的偏相关系数, 该相关系数的计算公式如式(2)所示:
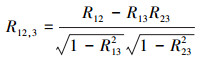
|
(2) |
同理, 参考公式(2), 可计算得到R14, 3和R24, 3。其中R14, 3是排除平均温度共同作用之后计算得到的NDVI与日照辐射的偏相关系数;R24, 3是排除平均温度共同作用之后计算得到的降水与日照辐射的偏相关系数。公式(2)中, R12为NDVI与降水的相关系数, R13为NDVI与温度的相关系数, R23为降水与温度的相关系数。
本文首先分析1982—2015整个时间段上NDVI与降水的偏相关系数(RNDVI-Prep)的分布规律, 并排除平均气温和日照辐射的共同作用。然后以17年为一个时间窗口, 把1982—2015整个时间段共34年平均分两个时间段。第一个17年为1982—1998;最后一个17年为1999—2015。然后使用时间滑动窗口, 每次向后滑动一年, 共产生18个时间窗口。在每个时间窗口i内计算欧亚大陆陆域植被在不同季节对降水的响应关系, 即排除温度和日照辐射共同作用后的NDVI与降水的偏相关关系RNDVI-Prep(i)。将18个时间窗口的偏相关系数采用最小二乘法做趋势分析[17], 进而得到RNDVI-Prep(i)的斜率, 即为TrendR。
同时, 为了表征每个像素上植被对降水的依赖程度和以及该依赖程度的变化趋势, 我们将像素分为四类。在每个像素(x, y)上, 将18个滑动窗口上的偏相关系数RNDVI-Prep(i)做一次筛选, 将偏相关系数为正, 且置信度大于90%(即偏相关系数的绝对值|R|≥0.437)的第i个时间窗口选出来, 并将T(i)标记为1, 不满足这个置信度的则标记为0, 公式如下所示:
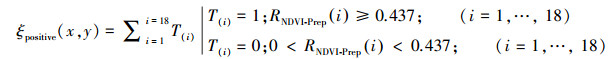
|
(3) |
计算每个像素上ξpositive(x, y)的值, 如果ξpositive(x, y)≥10, 则表明该像素上不同时间滑动窗口偏相关系数RNDVI-Prep(i)为正且显著相关(P < 0.1)的时间段超过9个, 即为总时间窗口的一半以上。这表明该像素上植被对降水的依赖程度较高。如果再叠加上植被与降水的偏相关关系RNDVI-Prep(i)的趋势TrendR, 则可以较为精确地描述每个地区的植被与降水的关系。例如, 某个像素的ξpositive(x, y) ≥10, 而且该像素上偏相关系数的趋势TrendR为正, 则表明像素上对降水的依赖程度较高, 且在1982至2015年间有增强的趋势。
相反, 在每个像素(x, y)上将18个滑动窗口上的相关系数为负且置信度大于90%的第i个时间窗口选出来, 并将T(i)标记为1, 不满足这个置信度的则标记为0, 公式如下所示:

|
(4) |
计算每个像素上ξnegative(x, y)的值, 如果ξnegative(x, y) ≥10, 则表明该像素上不同时间滑动窗口RNDVI-Prep(i)为负且偏相关系数显著相关(P < 0.1)的时间段为总时间窗口数的一半以上。这表明该像素上植被对降水的依赖程度较低, 且过多降水会导致温度和日照辐射相应的减少, 越不利于植被生长。
2 水文气象条件根据Nemani等[6]提出的植被生产力与光照、温度、水分因子这3个自然变量的胁迫关系模型, 本文将该模型由年尺度分解到每个季节。如图 1所示, 春季受水分胁迫(红色区域)影响最大的研究区域西起欧洲伊比利亚半岛南部, 贯穿地中海北岸、阿拉伯半岛、伊朗高原、南亚次大陆, 直至孟加拉湾地区。春季受日照辐射胁迫(绿色区域)的地区主要集中在欧洲、中国华南地区和东南亚地区。高纬度地区在春季主要受温度胁迫(蓝色区域)。在春季, 中国北方地区、蒙古高原、中亚地区的植被同时受水分和温度的胁迫(粉色区域)。

|
| 图 1 植被生长所受自然胁迫条件的季节分布 Fig. 1 Distribution of seasonal potential climatic constraints to plant growth 红色代表水分胁迫区域(Water, W), 蓝色代表温度(Temperature, T)胁迫区域, 绿色代表日照辐射(Radiation, R)胁迫区域 |
如图 1所示, 在夏季, 由于气温升高, 俄罗斯境内大部分地区的植被从春季时受温度胁迫转变为受日照辐射胁迫(绿色区域), 只有俄罗斯远东少部分地区和中国的青藏高原部分地区依然保持受温度胁迫。植被受水分条件胁迫的区域(红色区域)主要集中在伊比利亚半岛、亚平宁半岛、巴尔干半岛、阿拉伯半岛和中亚等地区。其余地区, 包括中国东部和南部地区、南亚及东南亚地区, 其植被大多是受日照辐射胁迫。图 1显示, 相对于夏季, 在秋季受水分胁迫的地区有向南移动的趋势, 在欧洲和中亚地区有所减少, 但是在印度西北部地区呈增加趋势。欧洲大部分地区和中国南部、日本、南亚及中南半岛、东南亚地区的植被主要受日照辐射胁迫。相对于夏季, 中国西北部和蒙古国地区的植被在秋季同时受水分和温度胁迫的区域(粉色区域)有所扩展。
通过上述3个自然胁迫条件的分季节分析可以发现, 在欧亚大陆植被生长对降水的响应有明显的时空差异, 其研究必须在时间尺度和空间尺度同时展开。
3 季节变化通过计算1982—2015年整个时间段上NDVI与降水的偏相关系数RNDVI-Prep(排除了温度与日照辐射的共同作用), 得到欧亚大陆每个像素上春、夏、秋3个季节植被生长对降水的响应关系(图 2)。在春季, 植被生长与降水呈显著正相关关系(P < 0.01)的地区与WWF发布的部分生态区边界有较高的一致性, 这些区域主要集中在东欧大草原(Pontic steppe, PS)、中亚地区的干旱草原与荒漠地区(例如:Central Asian northern desert, CAND), 中国东北和蒙古国东部的草原区(例如:Mongolian-Manchurian grassland, MG)。与此相对, 植被生长与降水的关系为显著负相关(P < 0.01)的区域所处的纬度相对正相关地区的纬度普遍较高, 主要集中北欧瑞典、芬兰以及俄罗斯境内60°N以北的高纬度地区。这些地区的负相关可能与春季的降雪影响有关, 加之该地区在春季主要受到温度胁迫, 过多的降水会使得春季的温度升温来的缓慢, 从而影响这些地区的植被生长。

|
| 图 2 植被生长与降水的偏相关系数分布图 Fig. 2 Distribution of partial correlation coefficient between vegetation growth and precipitation R=±0.447, R=±0.408, R=±0.349, R=±0.295, R=±0.232对应的置信度分别为99%, 98%, 95%, 90%和80%; CAND,Central Asian northern desert;PS, Pontic steppe;MG,Mongolian-Manchurian grassland;YGT,Yamal-Gydan tundra;WST,West Siberian Taiga;EST East Siberian Taiga |
夏季由于温度的增长, RNDVI-Prep呈显著正相关(P < 0.01)的区域明显增加。这些区域广泛地分布在35°—60°N之间(图 2中虚线所示)的欧亚大陆干旱、半干旱地区。它们西起东欧大草原, 贯穿中亚草原和干旱地区, 东至我国东北和蒙古国地区的草原生态区。另外巴基斯坦和印度西部的干旱森林和荒漠地区, 以及印度南部疏林生态区的植被也表现出与降水呈显著正相关关系。需要指出的是, 在我国华北平原和中欧部分地区, 农业区域占比很高, 这些地区的植被生长与降水也呈现很高的正相关关系。与此相对, 植被生长与降水呈负相关(P < 0.01)的区域对比春季明显减少, 这些区域主要集中在北欧及俄罗斯境内的部分高纬度地区。这些地区的植被在夏季虽然主要受到日照辐射胁迫, 但是夏季的降水还是会导致区域性的降温, 因此可能对植被生长造成不利的影响。值得注意的是, 在阿富汗南部和巴基斯坦西南部以及阿拉伯半岛的荒漠地区, 也有部分区域表现出植被生长与降水的负相关关系, 这可能主要是由于本文选取的最小NDVI阈值为0.1, 并没有很好地将该地区的非真NDVI像素剔除所致(图 2)。
相对于春季和夏季, 植被与降水的相关关系在秋季则更为集中。显著正相关(P < 0.01)的区域主要集中环黑海的中东欧地区, 印度西北部的干旱地区, 中国北方干旱的黄土高原和鄂尔多斯高原, 以及印度尼西亚苏门答腊和加里曼丹岛南部的雨林地区。与此相对, 植被生长与降水呈显著负相关的地区主要分布在北欧斯堪的纳维亚半岛中东部的泰加森林(Taiga), 并且与该半岛南端的混合林生态区有着明显的差别, 可明显看出WWF生态区划边界南北两侧的差异。在俄罗斯, 西西伯利亚泰加森林(West Siberian Taiga, WST)和东西伯利亚泰加森林(East Siberian Taiga, EST)对降水的响应也有着明显差异, 西西伯利亚地区植被对降水的关系以负相关居多, 东西伯利亚地区植被对降水的关系以正相关居多。值得关注的是, 在俄罗斯极地地区的亚马尔-格达苔原(Yamal-Gydan tundra, YGT), 植被生长与降水表现出极强的正相关关系, 与其他高纬度生态区的响应关系的表现极为不同。通过分析该生态区的植被与降水的时间序列可知, 这主要是由于该地区近十多年来秋季降水显著减少, 并且植被指数NDVI在2013—2015也相对于往年减少较明显所致。
4 趋势分析通过计算1982—2015年间的18个时间窗口的偏相关系数的斜率, 进而得到每个像素上植被指数与降水的响应关系的趋势(图 3), 即为TrendR。如果一个像素上NDVI与降水的相关性的趋势为正值, 则表明该像素上的植被对降水的依赖在1982—2015年之间越来越高。反之, 则表明该像素上的植被对降水的依赖在这34年之间在减小。如图 3所示, 植被与降水的偏相关关系RNDVI-Prep(i)的趋势TrendR在春、夏、秋3个季节上都表现出显著的空间分异特征。

|
| 图 3 1982—2015年间植被生长与降水的偏相关系数的趋势分布图 Fig. 3 Trend of the partial correlation coefficient between vegetation growth and precipitation from 1982 to 2015 |
Piao等[18]通过计算植被生产力与温度的偏相关系数的趋势发现北半球植被与温度的相关关系在减弱。同理, 针对RNDVI-Prep(i)的趋势TrendR分析虽然能表征植被对降水相关系数的变化趋势, 但通过判断TrendR的正负关系和置信度来判断植被生长对降水的响应关系是有一定的局限性的。这主要是因为18个滑动窗口得到如果是非显著相关的RNDVI-Prep(i), 但通过趋势计算也可能得到一个显著增强的TrendR。例如, 当某上像素上植被与降水的偏相关关系由显著负相关慢慢转变为非显著的正相关, 如果只依靠TrendR来描述该地区植被与降水的关系, 便会表述成该地区植被对降水的依赖程度在增加, 这显然是不客观、不准确的。因此, 本文结合TrendR和本文数据方法部分介绍的ξ(x, y)这两个要素来共同表征植被对降水的依赖程度, 从而将研究区域的像素分成4类。
表 1中, 第一类像素(在图 4中标记为红色)表示在1982至2015年间, 在18个滑动时间窗口上, RNDVI-Prep(i)呈显著正相关(P < 0.1)的时间窗口超过一半(即ξ(x, y)≥10), 因此本文设定该像素的植被在大部分时间段对降水形成显著依赖, 而且由于TrendR为正, 这说明该像素上植被生长对降水的依赖程度在增加。
| 分类要素 Classification element |
满足条件 Condition |
分类 Category |
| TrendR ξ(x, y) |
趋势为正 ξpositive(x, y)居多 |
第1类(红色) |
| TrendR ξ(x, y) |
趋势为负 ξpositive(x, y)居多 |
第2类(橙色) |
| TrendR ξ(x, y) |
趋势为正 ξnegative(x, y)居多 |
第3类(绿色) |
| TrendR ξ(x, y) |
趋势为负 ξnegative(x, y)居多 |
第4类(蓝色) |

|
| 图 4 植被生长对降水的依赖程度 Fig. 4 Dependency of the vegetation growth to precipitation BMF, Baltic mixed forests; MG, Mongolian-Manchurian grassland; SRT, Scandinavian and Russian taiga; WST, West Siberian taiga; PS, Pontic steppe; KS, Kazakh steppe; TD, Thar Desert; SDF, South Deccan Plateau dry deciduous forests |
同理, 第二类像素(橙色)表征的是植被在大部分时间段对降水形成显著依赖, 但是TrendR为负, 这说明该像素上植被生长对降水的依赖程度在减小。依此类推, 第三类像素(绿色)表征的是植被在大部分时间段对降水形成显著相斥, 但是TrendR为正, 这说明该像素上植被生长对降水的响应关系在由相斥转为逐步依赖。第四类像素(蓝色)表征的是植被在大部分时间段对降水是显著相斥, 而且TrendR为负, 这说明该像素上植被生长对降水表现出相斥的程度在继续加大。
通过对春、夏、秋3个季节植被对降水依赖程度的分析(图 4), 可以看出在春季植被生长对降水表现出相斥的区域零星地分布在高纬度地区(图 4), 包括英国南部和北欧斯堪的纳维亚半岛部分生态区, 以及俄罗斯的西伯利亚部分区域。与此相对, 植被对降水表现出依赖的地区主要集中在35°N至60°N之间的地区。在欧洲表现最为明显的生态区是波兰西北部的波罗的海混合林(Baltic mixed forests, BMF)。另外, 在中亚地区的干旱草原和荒漠地区, 中国东北和蒙古国东部的草原区(MG), 植被不仅对降水在大部分时间段表现出显著依赖(橙色像素), 而且部分地区的依赖程度还在不断的加强(红色像素)。
相对于春季, 在夏季(图 4), 植被对降水表现出依赖的地区(红色和橙色像素)纬度有明显抬升, 这些地区主要集中在东欧大草原(Pontic steppe, PS)和中亚地区的哈萨克草原(Kazakh steppe, KS)以及中国东北和蒙古国东部的草原区(MG)。另外, 在巴基斯坦和印度西北部地区的塔尔沙漠(Thar Desert, TD)和南部的南德干高原干旱落叶森林(South Deccan Plateau dry deciduous forests, SDF), 以及印度尼西亚的雨林等地区, 植被对降水都表现出依赖, 但是依赖程度在减弱(橙色像素)。在夏季植被对降水表现出相斥的地区(绿色和蓝色像素)主要集中在60°N以北更高纬度生态区, 如斯堪的纳维亚和俄罗斯泰加林(Scandinavian and Russian taiga, SRT)和西西伯利亚泰加林区(West Siberian taiga, WST)。
由图 4所示, 在秋季, 植被对降水表现出依赖的地区相对于春季和夏季则表现的较为集中。在欧洲, 这些地区主要集中在东欧大草原(PS)和环黑海的众多混合林生态区。在亚洲, 植被对降水表现出依赖的地区主要集中在巴基斯坦和印度西北部的干旱荒漠地区(例如, 塔尔沙漠, TD)和印度尼西亚苏门答腊岛和婆罗洲南部的雨林地区。中国的黄土高原混合林和鄂尔多斯高原草地生态区的植被对降水也有很强的依赖关系, 且依赖程度在增强。与此相对, 在欧洲地区, 植被对降水表现出相斥的区域(绿色和蓝色像素)主要集中在法国东部的西欧阔叶林区和瑞典中部的泰加林区地区。在亚洲, 植被对降水表现出相斥的地区主要集中在中亚的干旱荒漠地区以及俄罗斯的西西伯利亚泰加林区。
5 讨论本文使用了目前最新的第3代植被指数NDVI3g数据来表征欧亚大陆陆域植被生长状态。与之前文献所采用的方法不同[7, 13, 18], 本研究采用最新的全球空间分辨率为0.5度的气象格点数据(CRUTS V4)通过最简单的临近采样法重新采样至与植被数据精度(1/12度)保持一致, 参与其后的相关系数计算。这样的处理方法的有两个优点:一方面将对植被数据的细节进行最大程度的保留;另一方面, 考虑到气象要素中, 除了降水以外, 气温和日照辐射在一定的范围内都有很好的空间一致性。因此把0.5度的气象数据采样成1/12度能最大程度上保留植被指数信息的细节, 得到更精细的分析结果。限于篇幅, 在未来可考虑采用ANUSPLIN等专业气象插值软件, 对比不同采样方法和插值工具对研究结果的影响。
另外, 本文通过改进Nemani等[6]提出的气象条件对植被生长的胁迫模型, 将该模型分解到春、夏、秋3个季节, 分析了欧亚大陆不同季节上气象条件对植被生长胁迫的基本分布情况。进一步, 本文以像素为基本研究对象, 通过偏相关关系计算, 分析了欧亚大陆每个像素上植被指数NDVI对降水在不同季节的响应关系, 并同时排除了温度和日照辐射的共同作用。研究表明这些地区植被生长对水资源的响应在不同季节、不同地点表现出明显的时间和空间分异特征, 并且其空间差异的边界与现有的生态区边界在某些地区吻合极好, 有可能成为未来生产更精细化的生态区划图的一个重要参考。
通过统计1982—2015整个时间段上植被NDVI与降水的响应关系, 我们发现在春季(图 2), RNDVI-Prep为显著正相关的地区基本限制在35°—60°N之间的东欧大草原、中亚地区的干旱草原与荒漠地区以及中国东北和蒙古国东部的草原区。而在夏季(图 2), 由于温度升高, 蒸发加大, 植被与降水呈正相关的地区相对于春季明显增加。而且这些区域西起东欧大草原, 东至我国东北和蒙古国地区的草原生态区, 贯穿了整个欧亚大陆的干旱和半干旱地区。不仅如此, 甚至在60°N以北的高纬度泰加林生态区, 很多地区的植被也表现出与降水的正相关关系。相对于春季和夏季, 植被与降水的相关关系在秋季(图 2)则更为集中, 主要集中环黑海的中东欧地区, 南亚次大陆西北部的干旱地区, 以及印度尼西亚的雨林地区。在中国, 北方干旱的黄土高原混合林生态区和鄂尔多斯高原草甸构成一条东北至西南走向的植被与降水呈正相关关系的生物带。
为了更好地了解NDVI与降水的响应关系在不同时间段上的变化, 本文使用了时间滑动窗口来分析该响应关系的在每个季节上的变化趋势。我们发现在春、夏、秋3个季节, 植被生长与降水响应关系在滑动时间窗口上的变化趋势表现出极大的空间异质特性(图 3)。在不同地点不同季节, 其植被对降水的依赖程度不同, 依赖程度随时间变化的趋势也不同。考虑到不同时间窗口对偏相关系数计算结果的影响, 本文同时使用偏相关关系的趋势TrendR和RNDVI-Prep(i)呈显著相关的所有时间窗口的个数ξ(x, y)来共同表征植被对降水的依赖程度, 将植被对降水敏感的区域分为四类(图 4)。35°N以北地区, 在春秋两季, 植被生长对降水呈依赖关系地区的纬度要低于那些植被对降水呈相斥关系地区的纬度。通过对比图 4, 可以明显发现在夏季受增温影响, 蒸发加大, 植被对降水呈依赖关系的区域比春季要多, 而且有向高纬度地区扩张的趋势。35°N以南地区, 植被对降水呈依赖关系的地区主要集中在夏、秋两季的南亚次大陆西北部的干旱、半干旱地区及东南亚的雨林地区。
本研究目前只考虑了气象因素对植被生长的影响, 并未考虑诸如人工施肥与灌溉、病虫害、霜冻、火灾等影响。例如, 本文在图 2中发现俄罗斯极地地区的亚马尔-格达苔原(Yamal-Gydan tundra, YGT)的植被和降水的响应关系在秋季与极地其他地区明显不同, 但是这是否也是和该地区驯鹿的过度养殖[19]有关还有待进行进一步研究。
6 结论采用1982—2015年GIMMS NDVI3g数据和气象格点数据对欧亚大陆不同生态区上植被对降水响应的季节规律进行了分析, 得到如下结论:
(1) 在春节, 植被对降水的响应关系(RNDVI-Prep)为显著正相关的地区广泛地分布在东欧和中亚地区的草原及荒漠地带, 以及中国东北和蒙古国东部的草原地区;在夏季, RNDVI-Prep为显著正相关的地区广泛地分布在35°N—60°N之间的欧亚大陆干旱、半干旱地区以及南亚次大陆西北和南部地区;在秋季, RNDVI-Prep为显著正相关的地区分布则相对于春夏两季显得更为集中, 主要分布在环黑海的中东欧地区, 印度西北部的干旱地区, 中国北方干旱的黄土高原和鄂尔多斯高原, 以及印度尼西亚苏门答腊和加里曼丹岛南部的雨林地区。
(2) 在欧亚大陆, 夏季植被呈现对降水依赖的地区所处纬度要高于春、秋两季。中亚干旱、半干旱生态区的植被在春、夏两季对降水依赖较强;南亚次大陆西北部地区的干旱、半干旱及东南亚的雨林地区的植被生长在夏、秋两季对降水依赖较强。
(3) 以生态区为视角, 了解欧亚大陆的植被对降水响应在不同季节的变化规律可以为不同区域应对和适应气候变化提供决策依据。
| [1] | 刘卫东. "一带一路"战略的科学内涵与科学问题. 地理科学进展, 2015, 34(5): 538–544. |
| [2] | 张琪, 袁秀亮, 陈曦, 罗格平, 李龙辉. 1982-2012年中亚植被变化及其对气候变化的响应. 植物生态学报, 2016, 40(1): 13–23. |
| [3] | 戚伟, 刘盛和, 赵美风. "胡焕庸线"的稳定性及其两侧人口集疏模式差异. 地理学报, 2015, 70(4): 551–566. |
| [4] | 薛海源, 陈海山, 华文剑. 内蒙古地区植被对气候变化的响应. 气象与减灾研究, 2015, 38(2): 8–15. DOI:10.3969/j.issn.1007-9033.2015.02.002 |
| [5] | 焦珂伟, 高江波, 吴绍洪, 侯文娟. 植被活动对气候变化的响应过程研究进展. 生态学报, 2018, 38(6): 2229–2238. |
| [6] | Nemani R R, Keeling C D, Hashimoto H, Jolly W M, Piper S C, Tucker C J, Myneni R B, Running S W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science, 2003, 300(5625): 1560–1563. DOI:10.1126/science.1082750 |
| [7] | Peng S S, Piao S L, Ciais P, Myneni R B, Chen A P, Chevallier F, Dolman A J, Janssens I A, Peñuelas J, Zhang G X, Vicca S, Wan S Q, Wang S P, Zeng H. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature, 2013, 501(7465): 88–92. DOI:10.1038/nature12434 |
| [8] | Shen M G, Piao S L, Cong N, Zhang G X, Jassens I A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Global Change Biology, 2015, 21(10): 3647–3656. DOI:10.1111/gcb.12961 |
| [9] | Tucker C J, Pinzon J E, Brown M E, Slayback D A, Pak E W, Mahoney R, Vermote E F, El Saleous N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. International Journal of Remote Sensing, 2005, 26(20): 4485–4498. DOI:10.1080/01431160500168686 |
| [10] | Tourre Y M, Jarlan L, Lacaux J P, Rotela C H, Lafaye M. Spatio-temporal variability of NDVI-precipitation over southernmost South America:possible linkages between climate signals and epidemics. Environmental Research Letters, 2008, 3(4): 044008. DOI:10.1088/1748-9326/3/4/044008 |
| [11] | White A B, Kumar P, Tcheng D. A data mining approach for understanding topographic control on climate-induced inter-annual vegetation variability over the United States. Remote Sensing of Environment, 2005, 98(1): 1–20. DOI:10.1016/j.rse.2005.05.017 |
| [12] | Harris I, Jones P D, Osborn T J, Lister D H. Updated high-resolution grids of monthly climatic observations-the CRU TS3.10 Dataset. International Journal of Climatology, 2014, 34(3): 623–642. DOI:10.1002/joc.3711 |
| [13] | Tan J G, Piao S L, Chen A P, Zeng Z Z, Ciais P, Janssens I A, Mao J F, Myneni R B, Peng S S, Peñuelas J, Shi X Y, Vicca S. Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere. Global Change Biology, 2015, 21(1): 377–387. |
| [14] | Olson D M, Dinerstein E, Wikramanayake E D, Burgess N D, Powell G V N, Underwood E C, D'Amico J A, Itoua I, Strand H E, Morrison J C, Loucks C J, Allnutt T F, Ricketts T H, Kura Y, Lamoreux J F, Wettengel W W, Hedao P, Kassem K R. Terrestrial ecoregions of the world:a new map of life on Earth:a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience, 2001, 51(11): 933–938. DOI:10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2 |
| [15] | 刘焱序, 傅伯杰, 王帅, 赵文武. 从生物地理区划到生态功能区划:全球生态区划研究进展. 生态学报, 2017, 37(23): 7761–7768. |
| [16] | Garrett H E, Woodworth R S. Statistics in Psychology and Education. New York: David McKay, 1981. |
| [17] | Chen J, Li Q L, Niu J, Sun L Q. Regional climate change and local urbanization effects on weather variables in Southeast China. Stochastic Environmental Research and Risk Assessment, 2011, 25(4): 555–565. DOI:10.1007/s00477-010-0421-0 |
| [18] | Piao S L, Nan H J, Huntingford C, Ciais P, Friedlingstein P, Sitch S, Peng S S, Ahlstrom A, Canadell J G, Cong N, Levis S, Levy P E, Liu L L, Lomas M R, Mao J F, Myneni R B, Peylin P, Poulter B, Shi X Y, Yin G D, Viovy N, Wang T, Wang X H, Zaehle S, Zeng N, Zeng Z Z, Chen A P. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nature Communications, 2014, 5: 5018. DOI:10.1038/ncomms6018 |
| [19] | Epstein H E, Myers-Smith I, Walker D A. Recent dynamics of arctic and sub-arctic vegetation. Environmental Research Letters, 2013, 8(1): 015040. DOI:10.1088/1748-9326/8/1/015040 |