 2018, Vol. 38
2018, Vol. 38文章信息
- 高珊, 尹航, 傅民杰, 吴明根, 董闯, 李龙.
- GAO Shan, YIN Hang, FU Minjie, WU Minggen, DONG Chuang, LI Long.
- 冻融循环对温带3种林型下土壤微生物量碳、氮和氮矿化的影响
- Effects of freeze-thaw cycles on soil microbial biomass carbon, nitrogen, and nitrogen mineralization in three types of forest in the temperate zone
- 生态学报. 2018, 38(21): 7859-7869
- Acta Ecologica Sinica. 2018, 38(21): 7859-7869
- http://dx.doi.org/10.5846/stxb201712152257
-
文章历史
- 收稿日期: 2017-12-15
- 网络出版日期: 2018-08-08





2. 长白山科学研究院, 安图 133613
2. Changbai Mountain Scientific Research Institute, Antu 133613, China
土壤冻融循环是中高纬度和高海拔地区秋末春初季常见的自然现象[1], 主要是指由于季节或昼夜气温的变化使得土壤温度在0℃下波动而产生冻结和融化的一种物理现象和作用[2]。冻融过程通过改变土壤中水分状况及分布情况而影响土壤的物理、化学、生物等性质, 对土壤微生物量产生以及氮素转化过程的影响极其复杂[3]。土壤微生物量是重要的土壤有机组分, 包括土壤微生物量碳和土壤微生物量氮, 是土壤中微生物分解矿化氮元素和有机物的关键指标之一, 而其变化趋势可以表现出土壤肥力的大小和受污染的程度[4]。氮素是森林植物生长必需元素, 同时也在森林生态系统生物化学循环中扮演至关重要的角色[5]。土壤有机氮的矿化是森林土壤氮循环的重要组成部分。有机氮矿化后产生的NH4+和NO3-为土壤微生物以及植物提供主要氮素来源[6]。冻融过程中温度的波动对土壤氮矿化和硝化作用均有较大影响[3]。Christopher[7]发现冻融影响土壤温度和含水量, 进而影响微生物功能及活性, 同时也是控制有机物质矿化的重要因子。Zhou等[8]发现冻融作用下死亡的土壤微生物细胞可以被其他的微生物细胞作为基质。冻融循环可以强烈影响土壤微生物群落结构和组成[9-10]。任伊滨等[11]发现乌伊岭湿地土壤随着冻融时间和频次的增加, 土壤中NH4+-N的含量呈现先增高后降低的趋势, 在3次冻融循环后出现最高值, 而SMBC和SMBN也呈现先增高后下降趋势。有关冻融循环对土壤物理性质以及营养元素的影响国内外已进行了大量的相关研究, 这些研究主要针对中纬度湿地土壤和农田土壤, 但有关冻融过程对我国温带森林土壤的微生物量及氮矿化的影响研究较少[12-13]。长白山地区是典型的北方温带森林生态系统样带区, 森林生态系统的碳、氮循环受土壤冻融循环过程的影响十分明显。本文以长白山地区3种不同林型土壤为研究对象, 通过室内模拟冻融培养试验, 探讨冻融过程对土壤微生物量和土壤氮矿化的影响, 为了解温带森林土壤碳、氮累积潜力和功能提供基础理论数据。
1 材料与方法 1.1 采样地点概况采样地点位于吉林省长白山自然保护区内, 该区域气候属于受季风影响的温带大陆性山地气候, 春季风大干燥, 夏季短暂温凉, 秋季多雾凉爽, 冬季漫长寒冷。年均气温在3—7℃, 年日照时数约2300 h。无霜期100 d左右, 年降水量700—1400 mm。土壤冻结期达7个月。每年10月中旬至11月中下旬和次年的3月中下旬至5月中旬存在多次冻融过程。在保护区内, 分别选择了3种森林类型, 分别是硬阔叶林(Hardwood broad-leafed forest, 记为K, E:127°49′16.97″;N:2°17′09.54″;海拔:1100 m)、红松阔叶林(Korean pine broad-leaved forest, 记为HK, E:128°05′35.47″;N:42°24′43.13″;海拔:761 m)和次生白桦林(Secondary birch forest, 记为B, E:128°04′30.26″;N:42°25′16.22″;海拔:751 m)。3种林型均为天然林。
1.2 研究方法 1.2.1 试验样品采集在2015年10月底采集了硬阔叶林(K)、红松阔叶林(HK)和次生白桦林(B)3种类型的森林土壤。在上述3种森林类型内, 每个林型选取40 m×40 m规格的固定样地3块, 每个固定样地按对角线法选取5个采样点, 每个采样点间隔大于10 m。土壤采样时, 先将土壤表面的凋落物清理干净, 以长宽各20 cm规格分别采集0—10 cm层(上层)和10—20 cm层(下层)的土壤样品。每块样地采集的新鲜土壤样品剔除石块和其他杂质, 按同层次混合均匀后过2 mm筛, 并置于室内风干后备用。各林型土壤的基本理化性质如表 1。
| 林型 Forest types |
硬阔叶林 Hardwood broad-leaved forest |
红松阔叶林 Korean pine broad-leaved forest |
次生白桦林 Secondary birch forest |
|||
| 层次Soil layer/cm | 0-10 | 10-20 | 0-10 | 10-20 | 0-10 | 10-20 |
| 总氮Total nitrogen/(g/kg) | 10.0±1.5 | 6.1±207 | 7.4±3.0 | 1.7±0.2 | 12.3±4.1 | 6.5±1.3 |
| 总磷Total phosphor/(g/kg) | 1.3±0.2 | 1.1±0.1 | 1.0±0.2 | 0.6±0.2 | 1.6±0.4 | 0.9±0.1 |
| 总钾Total potassium/(g/kg) | 5.3±0.1 | 5.6±1.4 | 5.5±0.9 | 4.3±0.7 | 5.3±0.1 | 5.1±0.7 |
| 有机质Organic matter/(g/kg) | 145.5±38.5 | 65.1±7.8 | 111.5±26.8 | 39.1±12.3 | 109.7±22.3 | 52.5±6.4 |
| 水解氮Hydrolytic nitrogen/(mg/kg) | 156.9±8.1 | 216.0±29.5 | 302.9±38.9 | 158.7±43.0 | 280.2±2.3 | 237±32.2 |
| 速效钾Available potassium/(mg/kg) | 161.1±66.9 | 50.1±7.7 | 90.0±39.0 | 36.2±10.5 | 103.5±42.2 | 39.8±7.9 |
| 容重Bulk density/(g/cm3) | 0.6±0.0 | 0.8±0.0 | 0.6±0.1 | 1.6±0.2 | 0.6±0.1 | 1.1±0.0 |
根据历年气象数据, 长白山地区10月中旬至11月中下旬的冻结温度与融化温度分别介于-20—0℃和0—10℃之间, 冻融过程持续的天数为20—30 d左右。依据上述气候特点, 本次室内模拟冻融试验选取自然条件下的居间数据作为试验条件, 即将结冻温度下限设置为-15℃, 解冻温度上限设为5℃, 模拟冻融过程采用梯度降温模式。试验具体过程如下:将上述每种林型不同土层过筛后的风干土壤(1000 g)与少量(50 g)同一样地新鲜土壤混合均匀后, 调节含水量使其达到田间最大持水量的60%, 并置于大口塑料瓶中, 封口后放入25℃的恒温培养箱中预培养7 d, 期间每天开口通气1次。上述预培养结束后, 将培养温度下调至5℃继续恒温培养4 d, 使培养条件接近秋季土壤冻结前条件, 培养期间水、气调节方法同预培养。随后将培养温度以2 ℃/h变温速率由5 ℃逐渐降至-15℃, 并在-15℃条件下持续培养33 h, 然后再以相同变温速率将培养箱温度逐渐提升至5℃, 在此温度下培养19 h后, 形成一次冻融循环过程。后续培养重复前面的变温过程和持续时间。模拟试验中, 以2℃/h变温速率计算, 培养箱内温度由5℃降至-15 ℃或由-15℃升至5℃均需要用时10 h, 因此, 试验中一次冻融循环中, 0℃以下结冻时间为48 h, 其中-15℃持续时间为33 h, 0℃以上解冻时间为24 h, 其中5℃持续时间为19 h, 一次冻融循环周期用时72 h。试验共进行8次循环。每次冻融循环结束时取样测定土壤样品的NH4+-N、NO3--N、SMBC、SMBN和含水量等指标。每个林型土壤样品设3次重复。
1.2.3 土壤样品测定SMBC和SMBN浸提采用氯仿熏蒸K2SO4提取法[14]。SMBC用重铬酸钾氧化法测定。SMBN采用AA3流动分析仪(Seal Analytical, 德国)测定。土壤NH4+-N和NO3--N采用2 mol/L KCl溶液浸提, AA3流动分析仪测定。净硝化率、净氮矿化率计算公式[6]如下:
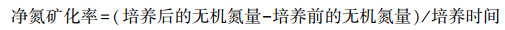
|
(1) |
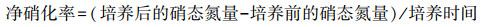
|
(2) |
文中数据均采用SPSS 17.0软件进行统计分析。不同冻融循环周期下3种林型下的土壤氮矿化率及SMBC和SMBN差异显著性比较均采用单因素方差分析法(One-way ANOVA);多重比较采用Duncan法(α=0.05);同一林型上、下土层土壤微生物量及氮矿化的差异显著性分析采用配对样本t-检验方法。所有图形均采用Excel 2003软件绘制。图中短竖线代表标准偏差。
2 结果分析 2.1 冻融循环对土壤微生物量碳(SMBC)的影响由图 1可知, 3种林型下的SMBC的空间分布对冻融循环的响应表现不同, 其中, 次生白桦林SMBC在整个冻融循环周期上层SMBC显著高于下层(P<0.05), 而硬阔叶林和红松阔叶林则分别在第2次冻融循环和第4次冻融循环后才表现出SMBC空间分布特征。此外, 冻融过程对SMBC影响因林型和土壤层次的不同表现出较大的差异, 红松阔叶林、硬阔叶林和次生白桦林上层土壤的SMBC动态分别表现出了单峰、双峰和三峰模式。其中, 红松阔叶林最大SMBC峰值比其CK增加100.3%;硬阔叶林SMBC两次SMBC峰值间无显著差异(P>0.05), 均比CK增加90%以上;次生白桦林SMBC在冻融过程中变化最为剧烈, 分别在第1、第3和第7次冻融循环时出现较大的提升, 但3次峰值间差异不显著(P>0.05), 且比CK增加的程度也明显低于其他2种森林类型。3种林型上层土壤各冻融循环频次的SMBC含量多高于其CK处理。3种林型下层土壤在冻融过程中均出现两次较大的SMBC波动, 且在峰值出现的次数与峰值出现的时间上与其对应的上层土壤存在明显不同。3种林型下层土壤的SMBC除峰值外, 其他冻融循环频次的SMBC含量与CK相近或低于CK处理(图 1)。

|
| 图 1 冻融循环过程中3种林型土壤微生物量碳变化 Fig. 1 Changes of soil microbial biomass carbon in three forest types during freezing and thawing cycles K, 硬阔叶林Hardwood broad-leaved forest; HK, 红松阔叶林Korean pine broad-leaved; B, 次生白桦林Secondary birch forest |
冻融过程中, 3种林型土壤的SMBN的垂直空间分布规律一致, 均表现出上层SMBN显著高于下层的特征(图 2)。但冻融过程中SMBN时间变化特征在林型与不同土壤层次间均存在明显的不同:从上层土壤的SMBN时间变化动态来看, 次生白桦林、红松阔叶林和硬阔叶林分别表现出单峰、双峰和三峰变化特征, 其中, 红松阔叶林和硬阔叶林在前5次冻融循环中SMBN动态基本一致;3种林型SMBN的最大峰值均出现于第5次冻融循环, 该阶段硬阔叶林、红松阔叶林和次生白桦林SMBN依次为(139.8±30.5)mg/kg、(121.2±9.2)mg/kg和(117.5±1.9)mg/kg, 分别比其对照增加59.5%、37.4%和152.4%, 而经过8次冻融循环后, 则分别比其CK下降了62.7%、34.6%和0.5%, 且均为各林型冻融过程中的最低值。3种林型上层土壤的SMBN在各冻融频次间均表现出显著差异性(P<0.05), 表明冻融过程对上层土壤的SMBN有着重要的影响。3种林型下层土壤的SMBN变化动态与其上层存在明显的非同步性特征, 且仅有硬阔叶林土壤各冻融频次的SMBN间存在显著差异(P<0.05), 其他2种林型匀未表现出显著差异性(P>0.05), 表明冻融过程对各林型的10—20 cm下层土壤的SMBN影响作用明显弱于对0—10 cm上层土壤SMBN的影响。3种林型下层土壤中SMBN含量均在第8次冻融后降至最低, 与上层SMBN对冻融频次的反应表现一致。

|
| 图 2 冻融循环过程中3种林型土壤微生物量氮变化 Fig. 2 Changes of soil microbial biomass nitrogen in three forest types during freezing and thawing cycles |
由表 2可知, 冻融过程中, 每种林型同一层次土壤的微生物C/N在冻融频次间均存在显著差异(P<0.05), 表明周期性的冻融循环过程对各林型土壤中的微生物C/N(土壤微生物群落结构)产生了明显的影响。另外, 土壤微生物C/N变化动态因林型和土壤层次的不同也表现出一定的差异性, 其中, 硬阔叶林和红松阔叶林的上层土壤在经历3次冻融循环后, 土壤微生物C/N均显示出显著增加特征, 而次生白桦林上层土壤经历多次冻融循环(4—6次)后则显示出其微生物C/N显著下降特征。硬阔叶林下层土壤微生物C/N变化动态与其上层土壤相近, 但红松阔叶林和次生白桦林下层土壤微生物C/N变化明显区别于其上层土壤, 且微生物C/N波动范围也相对小于其上层土壤(表 2)。
| 林型 Forest types 层次/cm Soil layer |
硬阔叶林 Hardwood broad-leaved forest |
红松阔叶林 Korean pine broad-leaved forest |
次生白桦林 Secondary birch forest |
||||||
| 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | ||||
| 冻融频次 Frequency of freezing-thawing |
对照 | 6.6±1.2c | 7.8±2.8b | 5.8±1.5c | 9.0±2.2ab | 18.7±7.1ab | 9.9±2.5b | ||
| 1 | 6.0±0.8c | 8.9±1.1b | 4.2±0.2d | 8.5±3.5ac | 20.1±8.5a | 11.2±1.7ab | |||
| 2 | 8.6±0.8b | 5.8±1.6c | 6.8±0.9c | 12.3±2.2a | 12.5±0.6ac | 7.9±2.3b | |||
| 3 | 23.5±4.4a | 27.0±4.5a | 28.0±1.7a | 10.2±2.5a | 18.6±2.0a | 5.2±2.1c | |||
| 4 | 9.9±0.4bc | 4.9±0.5c | 20.9±8.8b | 4.8±1.1bc | 10.1±1.3bc | 7.2±0.6b | |||
| 5 | 4.6±1.1c | 11.5±1.4b | 6.4±1.2c | 4.5±1.0bc | 6.8±0.1c | 13.9±1.2a | |||
| 6 | 2.6±3.6b | 6.0±1.7c | 6.7±1.0c | 12.2±2.7a | 4.4±1.3c | 8.5±2.7bc | |||
| 7 | 8.6±3.0b | 9.3±3.3bc | 8.0±1.5cd | 3.3±0.4c | 16.9±5.4a | 9.2±2.1b | |||
| 8 | 24.0±4.4a | 25.0±1.4a | 12.3±3.1c | 6.9±4.3a | 16.8±2.2a | 5.5±0.4c | |||
| 不同的小写字母代表同层土壤不同冻融频次间的微生物C/N差异显著(P<0.05, n=3) | |||||||||
由图 3可知, 冻融循环过程对3种林型土壤NO3--N含量变化的影响较小, 未出现明显的NO3--N波动, 每种林型同层次不同冻融频次的土壤NO3--N含量间均无显著差异(P>0.05), 且与冻融前的CK处理间也未表现出显著差异(P>0.05)。但从冻融循环期间NO3--N总体变化趋势上来看, 3种林型上、下层的变化动态有所不同, 其中, 3种林型上层土壤NO3--N含量随着冻融频次的增加整体呈现相同变化趋势, 即各林型均在第2次冻融循环周期后出现升高现象, 而NO3--N最低值均出现在第8次冻融循环后, 相比CK处理降低了7.0%(硬阔叶林)、10.8%(红松阔叶林)和12.6%(次生白桦林)。3种林型下层土壤NO3--N含量变化表现不同, 其中, 在冻融循环过程中, 硬阔叶林和次生白桦林下层土壤NO3--N含量均未高于CK处理, 经8冻融循环过程后土壤NO3--N分别下降了10.2%和8.2%, 而红松阔叶林下层土壤NO3--N含量峰值出现在第7次冻融循环周期后。硬阔叶林、红松阔叶林和次生白桦林上层土壤的土壤NO3--N含量显著高于其下层土壤(P<0.05), 表现出明显的空间垂直分布特征。

|
| 图 3 冻融循环过程中3种林型NO3--N变化 Fig. 3 Changes of NO3--N in three forest types during freezing and thawing cycles |
由图 4可知, 冻融循环对3种林型不同层次的土壤NH4+-N影响不同。在上层土壤中, 土壤NH4+-N含量随着冻融频次的增加均呈现先明显升高而后降低的趋势, 均在第3、4、5次冻融循环时出现明显的NH4+-N释放特征, 其中以第5次冻融循环时的土壤NH4+-N含量为最高(硬阔叶林:(171.8±94.2) mg/kg、红松阔叶林:(87.7±26.3) mg/kg和次生白桦林:(102.6±43.3) mg/kg), 依次比冻融前CK处理增加了81.3%(硬阔叶林)、57.9%(红松阔叶林)、264.2%(次生白桦林)。3种林型的上层土壤NH4+-N含量在各个冻融频次间均存在显著差异(P<0.05)。各林型下层土壤NH4+-N含量变化动态差异较大, 硬阔叶林土壤NH4+-N含量峰值((54.6±12.4) mg/kg)出现于第8次冻融循环后, 但仍比冻融前CK处理下降了18.6%;红松阔叶林下层土壤NH4+-N在第3次冻融循环后达到最大峰值((46.4±31.8) mg/kg), 最后一次冻融循环结束时NH4+-N含量比CK处理增加了17.3%;次生白桦林下层土壤NH4+-N含量随着冻融频次的增加呈现先下降后上升的趋势, 最后一次冻融循环结束时土壤NH4+-N含量比CK处理增加了49.9%。冻融循环过程对3种林型土壤NH4+-N分布过程产生明显影响, 但该影响表现出阶段性特征, 即在第3—第5次循环期间存在明显的NH4+-N空间分布, 而其他冻融循环过程对NH4+-N空间分布无明显影响(图 4)。

|
| 图 4 冻融循环过程中3种林型NH4+-N变化 Fig. 4 Changes of NH4+-N in three forest types during freezing and thawing cycles |
冻融过程中3种林型上层和下层土壤净硝化率整体波动较大, 且冻融过程对土壤硝化作用的影响因林型与土壤层次而异(图 5)。其中, 3种林型上层土壤净硝化率在8次冻融循环过程中表现出较好的一致性, 均在第2、第5和第7次循环过程中表现出明显的NO3--N净释放过程, 且均以第2次冻融循环净硝化率最高(硬阔叶林:(1.9±1.5) mg kg-1 d-1;红松阔叶林:(1.2±0.1) mg kg-1 d-1;次生白桦林:(2.0±2.4) mg kg-1 d-1), 第5和第7次循环次之, 而其他冻融循环过程中, 净硝化率均为负值, 表现出明显的土壤NO3--N净吸收现象, 但吸收强度在林型和冻融频次间有所不同。3种林型中, 硬阔叶林和红松阔叶林下层土壤的硝化过程表现一致, 且均在第3次和第8次冻融循环过程中表现出土壤NO3--N净释放现象, 而此期间上层土壤则表现出明显的NO3--N净吸收过程。次生白桦林下层土壤在第2、第6和第8次冻融循环中均与上述2种林型下层土壤表现出相反的NO3--N释放特征(图 5)。

|
| 图 5 冻融循环过程中3种林型净硝化率变化 Fig. 5 Changes of Net nitrification rate in three forest types during freezing and thawing cycles |
从图 6中可知, 冻融作用下3种林型下的土壤净氮矿化率均表现出较大的变化波动, 且每种林型各冻融频数间的土壤净氮矿化率差异显著(P<0.05)。3种林型上层土壤在第2、3、5次冻融循环时净氮矿化率均为正值, 且均是第3次冻融循环的土壤净氮矿化率显著高于同林型同层次的其他次冻融循环(P<0.05), 表现出强烈的无机氮(NH4+-N或NO3--N)释放特征。3种林型上层土壤均在第6次冻融循环时表现最低净氮矿化率, 显示出明显的无机氮吸收现象。3种林型的下层土壤在第5和7次冻融循环的土壤净氮矿化率均为正值, 冻融循环初期的土壤净氮矿化率多为负值, 且最大无机氮释放与吸收的峰值出现的时间在林型间表现不同。

|
| 图 6 冻融循环过程中3种林型净氮矿化率变化 Fig. 6 Changes of Net N mineralization rate in three forest types during freezing and thawing cycles |
土壤中的SMBC和SMBN含量主要依赖于土壤微生物的种类和数量, 受土壤水分、温度、营养、土壤通气性等各种生态因子的强烈影响[7, 15]。虽然Koponen等[16]和Grogan等[1]曾报道在室内模拟试验条件下少次冻融循环未对SMBC产生明显影响, 但本研究发现冻融循环过程对3种林型土壤的SMBC时间动态影响较大且不同林型的SMBC变化表现迥异。已有研究表明, 持续的冻结过程会导致土壤细菌和放射线菌数量下降, 特别是对不耐低温的微生物种类和数量产生的影响更为明显, 而冻融过程则有利于真菌与放线菌的增加, 导致微生物种群结构和活性发生明显改变[1, 17-18]。本研究在-15—5℃的变温范围下, 通过8次强烈的冻融循环过程, 3种林型各冻融阶段中微生物C/N变化差异较大(表 2), 说明冻融过程中土壤水分周期性的凝结与融化和土壤温度的周期性极限波动对土壤微生物产生了明显影响, 进而导致3种林型土壤中的SMBC均产生明显时间波动特征。同时, 3种林型的不同植物群落构成, 也造成了土壤微生物群落的不同[11], 导致冻融过程对每种林型SMBC的影响不同。一般认为, 冻融过程会对土壤SMBN产生强烈影响, 如Fan等[19]发现冻融过程影响了青藏高原北缘高山土壤生态系统的SMBN含量。任伊滨等[11]采用原位法对小兴安岭湿地的研究发现, 冻融低温导致SMBN会随冻融时间的延续持续下降。范志平等[20]研究也表明, 经过10次模拟冻融循环后, SMBN含量同样表现出下降趋势。本研究中, 尽管不同林型和土壤层次的SMBN对冻融循环的响应存在差异, 但3种林型的SMBN均在整个冻融循环周期后下降至最低, 与上述原位模拟的研究相吻合, 说明低温与较大温差对冻融期土壤微生物产生了明显影响。本研究中土壤分层(上层:0—10 cm;下层:10—20 cm)虽然未按土壤发生学方法划分, 但仍然显示出所有林型的上层土壤SMBC和SMBN含量显著高于下层的特征。导致这种结果的主要原因在于上下层土壤的理化性质的差异。从表 1中可知, 3种林型的上层土壤在有机质、总氮及总磷等方面均明显高于下层土壤, 而土壤容重则显著低于下层土壤, 良好的营养与通气状况为上层微生物的繁殖创造了适宜条件, 由此导致了SMBC和SMBN垂直空间分布。Plymale等[21]在阔叶林的研究中也获得了与本试验相近的结果。
3.2 冻融循环对3种林型下的土壤NO3--N和NH4+-N的影响冻融循环对土壤氮矿化影响的众多研究表明, 土壤有机氮的矿化过程受冻融频数、冻结时间、融化温度以及土壤微生物群落构成和活性的多重影响[20, 22-24]。一般认为越冬和冰雪融化过程是控制N素循环和累积的关键因素, 冻融过程对土壤NO3--N含量影响显著[25]。Judd等[24]对美国Hubbard Brook温带森林流域土壤的研究表明, 冻融频数对NO3--N含量影响显著。然而, 本研究表明, 冻融循环过程对3种林型下的土壤NO3--N含量时间动态影响较小, NO3--N含量整体波动不明显。本次研究模拟的单次冻融循环的冻结时间(48 h)和循环周期(72 h)的设定均远大于实际或模拟试验设定的冻融周期经历的时间, 加之较大的冻融温差使得参与土壤硝化作用的微生物种类及数量在冻融试验的早期阶段即被筛选和确定。加之土壤冻结过程中大量底物和水分会被固定[26], 此时的土壤孔隙和氧气含量减少, 因此导致好氧的硝化过程被抑制而厌氧反硝化作用被促进[27], 由此可能导致本试验的NO3--N含量变化未表现出显著性波动特征。另外, 本研究中各林型土壤经过冻融循环过程后, 其上、下层土壤NO3--N含量均低于未冻结的CK处理。分析认为在室内模拟冻融培养试验中没有养分累积淋溶以及植被吸收和挥发作用等氮损失过程, 因此在多次连续的冻融循环作用下, 土壤氮矿化累计产物增加, 矿化作用受其抑制[28], 协同增强的反硝化作用共同导致了NO3--N含量的下降。Bochove等[29]研究结果也表明冻融循环过程导致了土壤NO3--N含量降低, 与本文结果相符。与NO3--N含量动态不同, 冻融循环过程中3种林型下的上层土壤NH4+-N含量随着冻融频次的增加均表现出了先明显升高而后降低的趋势。分析发现, 经过两次冻融循环过程后, 3种林型土壤中的SMBC逐渐增加(图 1)而SMBN逐渐下降(图 2), 而土壤C/N也逐渐下降(表 2), 说明土壤微生物数量及适宜性细菌群落在两次冻融循环后不断增加, 由此增强了土壤的氨化过程, 造成3种林型下的土壤NH4+-N含量的暴发性增加。任伊滨等[11]对乌伊岭湿地土壤的原位研究也获得了相似的结果。
3.3 冻融循环对3种林型下的土壤氮矿化的影响许多研究表明, 冻融过程对土壤氮素转化的影响作用主要是通过周期性的土壤温度与水分变化造成了土壤微生物群落多样性及其细胞生物活性的改变[7, 22, 30]。本研究中, 冻融处理后的土壤NH4+-N和NO3--N相对于未经冻结的CK处理波动较大, 也说明冻融过程对参与土壤氮矿化与硝化微生物活动产生了强烈影响。同时, 3种林型的土壤微生物群落的差异性(图 1、图 2和表 2), 导致其土壤净氮矿化率和净硝化率对温度变化的响应不同[19]。另外, 冻融作用下3种林型下的土壤各冻融频数间的土壤净硝化率和净氮矿化率整体波动较大, 且上下层土壤在整个冻融期间内其土壤的矿化作用表现不一致, 这与前人[23, 28]研究所得结果相吻合, 即土壤氮矿化作用对冻融过程的响应受土壤层次以及冻融频次影响。同一温度条件下, 森林土壤有机氮的转化最直接影响因素就是森林植被的结构组成[20]。不同森林间有机质资源、群落结构组成以及微生物种类和数量的差异, 使得本研究中冻融循环对3种林型的土壤氮矿化作用差异较大。Urakawa等[23]对日本列岛10种温带森林的研究也有着相同的结果。本研究中, 3种林型的上层土壤在第3次冻融循环的土壤净氮矿化率均显著高于同林型同层次的其他次冻融循环, 表现出强烈的无机氮释放特征, 而在第6次冻融循环时均显示出明显的无机氮吸收现象。Amador等[31]认为短期冻融循环促使土壤氮素加快转化, 但随着冻融频次增加, 这种情况随之减弱, 土壤氮矿化作用受长期冻融产物积累的抑制而使土壤净氮矿化率减小。这说明虽然冻融过程中不同森林群落结构导致土壤有机氮矿化作用存在明显的分异性[23], 但冻融频次和温度的变化对森林氮矿化作用有相似而重要的影响。
4 结论(1) 冻融过程对温带3种林型下的土壤微生物量的影响不同, 不同林型的SMBC和SMBN的时间动态变化存在明显的差异性, 但均表现上层土壤中SMBC和SMBN含量多显著高于下层, 表现出明显的空间垂直变异性。整个冻融过程中其上层土壤表现出微生物量富集特征。
(2) 森林土壤NO3--N对冻融过程的响应不明显, 且冻融各阶段每种林型上、下层土壤NO3--N变化幅度较小, 均未表现出显著的时间动态变化差异。冻融过程对土壤NH4+-N变化的影响因冻融频次、林型和土壤层次而异, 且3种林型在第3—5次冻融循环后为NH4+-N释放过程, 而下层土壤在冻融期间存在明显分异性。
(3) 温带3种林型下的土壤氮矿(硝)化作用在冻融过程中受冻融频次、森林类型、土壤层次和冻融温度变化的共同影响。3种林型土壤在短期冻融循环内存在氮素释放过程, 而后受长期冻融产物积累的抑制表现出明显的无机氮吸收现象。
| [1] | Grogan P, Michelsen A, Ambus P, Jonasson S. Freeze-thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms. Soil Biology and Biochemistry, 2004, 36(4): 641–654. DOI:10.1016/j.soilbio.2003.12.007 |
| [2] | 王连峰, 蔡延江, 解宏图. 冻融作用下土壤物理和微生物性状变化与氧化亚氮排放的关系. 应用生态学报, 2007, 18(10): 2361–2366. |
| [3] | 蔡延江, 王小丹, 丁维新, 鄢燕, 鲁旭阳, 杜子银. 冻融对土壤氮素转化和N2O排放的影响研究进展. 土壤学报, 2013, 50(5): 1032–1042. |
| [4] | 陈怀满. 环境土壤学(第二版). 北京: 科学出版社, 2010: 1–103. |
| [5] | 殷睿, 徐振峰, 吴福忠, 苟小林, 熊莉, 唐仕姗, 谢承瀚, 杨万勤. 川西高山森林林窗对季节性冻融期土壤氮动态的影响. 生态学杂志, 2014, 33(9): 2483–2489. |
| [6] | 周才平, 欧阳华. 温度和湿度对长白山两种林型下土壤氮矿化的影响. 应用生态学报, 2001, 12(4): 505–508. DOI:10.3321/j.issn:1001-9332.2001.04.006 |
| [7] | Christopher S F, Shibata H, Ozawa M, Nakagawa Y, Mitchell M J. The effect of soil freezing on N cycling:comparison of two headwater subcatchments with different vegetation and snowpack conditions in the northern Hokkaido Island of Japan. Biogeochemistry, 2008, 88(1): 15–30. DOI:10.1007/s10533-008-9189-4 |
| [8] | Zhou W M, Chen H, Zhou L, Lewis B J, Ye Y J, Tian J, Li G W, Dai L M. Effect of freezing-thawing on nitrogen mineralization in vegetation soils of four landscape zones of Changbai Mountain. Annals of Forest Science, 2011, 68(5): 943–951. DOI:10.1007/s13595-011-0100-4 |
| [9] | 杨万勤, 冯瑞芳, 张健, 王开运. 中国西部3个亚高山森林土壤有机层和矿质层碳储量和生化特性. 生态学报, 2007, 27(10): 4157–4165. DOI:10.3321/j.issn:1000-0933.2007.10.024 |
| [10] | Sjursen H, Michelsen A, Holmstrup M. Effects of freeze-thaw cycles on microarthropods and nutrient availability in a sub-Arctic soil. Applied Soil Ecology, 2005, 28(1): 79–93. DOI:10.1016/j.apsoil.2004.06.003 |
| [11] | 任伊滨, 任南琪, 李志强. 冻融对小兴安岭湿地土壤微生物碳、氮和氮转换的影响. 哈尔滨工程大学学报, 2013, 34(4): 530–535. |
| [12] | 赵媛, 周旺明, 王守乐, 周莉, 于大炮, 代力民. 冻融对温带森林土壤碳、氮矿化作用的影响. 生态学杂志, 2017, 36(6): 1548–1554. |
| [13] | 贾国晶, 周永斌, 代力民, 周旺明. 冻融对长白山森林土壤碳氮矿化的影响. 生态环境学报, 2012, 21(4): 624–628. DOI:10.3969/j.issn.1674-5906.2012.04.006 |
| [14] | 吴金水. 土壤微生物生物量测定方法及其应用. 北京: 中国气象出版社, 2006: 65–67. |
| [15] | 林尤伟, 金光泽. 冻融期去根处理对小兴安岭6种林型土壤微生物量的影响. 生态学报, 2016, 36(19): 6159–6169. |
| [16] | Koponen H T, Jaakkola T, Keinänen-Toivola M M, Kaipainen S, Tuomainen J, Servomaa K, Martikainen P J. Microbial communities, biomass, and activities in soils as affected by freeze thaw cycles. Soil Biology and Biochemistry, 2006, 38(7): 1861–1871. DOI:10.1016/j.soilbio.2005.12.010 |
| [17] | Lipson D A, Schmidt S K. Seasonal changes in an alpine soil bacterial community in the colorado rocky mountains. Applied and Environmental Microbiology, 2004, 70(5): 2867–2879. DOI:10.1128/AEM.70.5.2867-2879.2004 |
| [18] | Six J, Bossuyt H, Degryze S, Denef K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil and Tillage Research, 2004, 79(1): 7–31. DOI:10.1016/j.still.2004.03.008 |
| [19] | Fan J H, Cao Y Z, Yan Y, Lu X Y, Wang X D. Freezing-thawing cycles effect on the water soluble organic carbon, nitrogen and microbial biomass of alpine grassland soil in Northern Tibet. African Journal of Microbiology Research, 2012, 6(3): 562–567. |
| [20] | 范志平, 李胜男, 李法云, 高洪翠, 闫加亮. 冻融交替对河岸缓冲带土壤无机氮和土壤微生物量氮的影响. 气象与环境学报, 2013, 29(4): 106–111. DOI:10.3969/j.issn.1673-503X.2013.04.017 |
| [21] | Plymale A E, Boerner R E J, Logan T J. Relative nitrogen mineralization and nitrification in soils of two contrasting hardwood forests:Effects of site microclimate and initial soil chemistry. Forest Ecology and Management, 1987, 21(1/2): 21–36. |
| [22] | 李志杰, 杨万勤, 岳楷, 贺若阳, 杨开军, 庄丽燕, 谭波, 徐振锋. 温度对川西亚高山3种森林土壤氮矿化的影响. 生态学报, 2017, 37(12): 4045–4052. |
| [23] | Urakawa R, Shibata H, Kuroiwa M, Inagaki Y, Tateno R, Hishi T, Fukuzawa K, Hirai K, Toda H, Oyanagi N, Nakata M, Nakanishi A, Fukushima K, Enoki T, Suwa Y. Effects of freeze-thaw cycles resulting from winter climate change on soil nitrogen cycling in ten temperate forest ecosystems throughout the Japanese archipelago. Soil Biology and Biochemistry, 2014, 74: 82–94. DOI:10.1016/j.soilbio.2014.02.022 |
| [24] | Judd K E, Likens G E, Buso D C, Bailey A S. Minimal response in watershed nitrate export to severe soil frost raises questions about nutrient dynamics in the Hubbard Brook experimental forest. Biogeochemistry, 2011, 106(3): 443–459. DOI:10.1007/s10533-010-9524-4 |
| [25] | Zhao Q, Zeng D H, Fan Z P. Effects of freeze-thaw on soil nitrogen and phosphorus availability at the Keerqin Sandy Lands, China. Journal of Forestry Research, 2008, 19(1): 44–48. DOI:10.1007/s11676-008-0007-6 |
| [26] | Brooks P D, Williams M W. Snowpack controls on nitrogen cycling and export in seasonally snow-covered catchments. Hydrological Processes, 1999, 13(14/15): 2177–2190. |
| [27] | 宋长春, 王毅勇, 王跃思, 赵志春. 季节性冻融期沼泽湿地CO2、CH4和N2O排放动态. 环境科学, 2005, 26(4): 7–12. |
| [28] | 傅民杰, 王传宽, 王颖, 刘实. 四种温带森林土壤氮矿化与硝化时空格局. 生态学报, 2009, 29(7): 3747–3758. DOI:10.3321/j.issn:1000-0933.2009.07.036 |
| [29] | Van Bochove E, Prévost D, Pelletier F. Effects of freeze-thaw and soil structure on nitrous oxide produced in a clay soil. Soil Science Society of America Journal, 2000, 64(5): 1638–1643. DOI:10.2136/sssaj2000.6451638x |
| [30] | 王常慧, 刑雪荣, 韩兴国. 温度和湿度对我国内蒙古羊草草原土壤净氮矿化的影响. 生态学报, 2004, 24(11): 2472–2476. DOI:10.3321/j.issn:1000-0933.2004.11.018 |
| [31] | Amador J A, Görres J H, Savin M C. Role of soil water content in the carbon and nitrogen dynamics of Lumbricus terrestris L. burrow soil. Applied Soil Ecology, 2005, 28(1): 15–22. DOI:10.1016/j.apsoil.2004.06.009 |