 2018, Vol. 38
2018, Vol. 38文章信息
- 旦智措, 鲍敏, 马存新, 李雷雷, 郝会文, 成帆, 才项卓玛, 陈振宁.
- DAN Zhicuo, BAO Min, MA Cunxin, LI Leilei, HAO Huiwen, CHENG Fan, CAI Xiangzhuoma, CHEN Zhenning.
- 青海玉树高原不同生境类型蝶类群落结构与多样性
- Community structure and butterfly diversity in different habitat types in the Qinghai Yushu plateau
- 生态学报. 2018, 38(21): 7557-7564
- Acta Ecologica Sinica. 2018, 38(21): 7557-7564
- http://dx.doi.org/10.5846/stxb201711272115
-
文章历史
- 收稿日期: 2017-11-27
- 网络出版日期: 2018-08-08





生物多样性是人类赖以生存的物质基础, 其保护是当前国际社会瞩目的重大环境问题, 对维持区域可持续发展具有重要的意义[1]。蝴蝶种类丰富、分布广泛, 环境的细微变化能够迅速、准确地反映在蝴蝶的种群动态和群落结构特征中, 因此, 蝴蝶非常适合用来监测环境变化趋势、生态系统健康状况、人力活动对生态系统的干扰程度等, 成为公认的生物多样性监测指示生物[2]。近几年来, 有研究者对玉树地区的蝴蝶资源进行过一些调查, 主要集中在种类调查和区系分析方面[3-5], 并未对该地区的蝴蝶群落多样性进行分析和评价。为进一步了解玉树高原地区蝴蝶种群动态和群落多样性, 在前人研究的基础上, 于2016—2017年对该地区的蝶类资源进行了系统的调查, 对该地区的蝴蝶群落多样性及月季变化进行研究, 为玉树高原地区的蝶类资源保护和环境质量监测提供基础资料。
1 自然环境概况玉树市地处青藏高原东部, 位于青海省玉树藏族自治州最东部, 境内平均海拔4490 m, 地形以山地高原为主, 通天河、扎曲在玉树市境内流过。地理位置为89°35′—97°55′E, 31°35′—36°30′N, 属于典型的高原高寒气候。昼夜温差大, 年均温-2.5—4.9℃, 全年降水量387—526.1 mm, 多集中于5—9月[5]。
2 研究方法 2.1 研究地概况根据环境、海拔高度和植被类型在玉树市选取5种不同生境类型:
Forest:森林(3570—3757 m), 位于龙日村沟内, 主要建群种有青海云杉(Picea crassifolia)、大果圆柏(Sabina tibetica)等。
Shrub:灌木(3501—3574 m), 位于通天河畔勒巴沟内, 灌木和草本植物丰富, 主要植物有中国沙棘(Hippophae rhamnoides)、美丽茶藨子(Ribes pulchellum)、甘青铁线莲(Clematis tangutica)、岩生忍冬(Lonicera rupicola)、刚毛忍冬(Lonicera hispida)、中国黄花柳(Salix sinica)、鲜黄小檗(Berberis diaphana)、西北小檗(Berberis franchetiana)、草玉梅(Anemone rivularia)、高原毛茛(Ranunculus tanguticus)等。
Alpine shrub:高寒灌丛(3735—3985 m), 位于索荣沟内, 灌丛和草本植物丰富, 主要植物有头花杜鹃(Rhododendron capitatum)、窄叶鲜卑花(Sibiraea angustata)、金露梅(Potentilla fruticosa)、昌都锦鸡儿(Caragana changduensis)、高山柳(Salix cupularis)、草玉梅(Anemone rivularia)、小嵩草(Kobresia humilis)、圆穗蓼(Polygonum macrophyllum)等。
Alpine meadow:高寒草甸草原(4183—4382 m), 位于勒巴沟内, 植被以高寒植被为主, 主要植物有小嵩草(Kobresia humilis)、圆穗蓼(Polygonum macrophyllum)、短管兔儿草(Lagotis brevituba)、垫状点地梅(Androsace tapete)、高原毛茛(Ranunculus tanguticus)、青藏龙胆(Gentiana futtereri)、矮火绒草(Leontopodium nanum)等。
Naked rock:裸岩(4427—4662 m), 位于索荣沟内, 海拔在5种生境中最高, 植被稀少, 主要植物有唐古特红景天(Rhodiola tangutica)、水母雪兔子(Saussurea medusa)、梭砂贝母(Fritillaria delavayi)、红花紫堇(Corydalis punicea)等。
2.2 研究方法(1) 研究时间
于2016和2017年的5—8月中旬逐月对玉树市所选取的5种生境采用样线法进行调查, 选择晴朗或多云天气, 蝶类活动频繁的10:00—17:00。
(2) 数据记录方法
生境内样线长度2 km, 以1—1.5 km/h的速度沿样线缓慢匀速前行, 样线宽度设置左右2.5 m。行进过程中仔细观察和记录所发现的蝴蝶种类及数量, 对于常见且能及时辨认的种类, 只记录数量, 不能及时辨认或疑难蝶种, 网捕后确认种类并原地释放, 当场不能确定的, 少量网捕编号, 带回实验室利用图鉴或请专家进行鉴定到种或属。
(3) 鉴定与数据统计
依据《中国蝶类志(上、下册)》、《中国蝴蝶图鉴(全4册)》、《中国灰蝶志》、《中国动物志·昆虫纲(第二十五卷)鳞翅目·凤蝶科》、《中国动物志·昆虫纲(第五十二卷)鳞翅目·粉蝶科》、《中国动物志·昆虫纲(第五十五卷)鳞翅目·弄蝶科》、《中国蛱蝶科志(鳞翅目, 锤角亚目)卷一》进行鉴定与分类[6-12], 分别统计数据。
2.3 数据分析(1) 多样性指数(H′)采用Shannon-Wiener公式[13-14]:
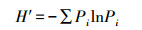
|
式中, Pi是第i种的个体比例, Pi =Ni/N, N是第i种的个体数, N是全部物种的个体总数。
等级多样性指数公式为:

|
式中, H′(G)为属级多样性, H′(S)为种级多样性。
(2) 均匀度指数(J)采用Pielou公式[14]:
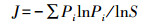
|
式中, J为均匀度, -∑PilnPi为Shannon-Wiener多样性指数, S为物种数。
(3) 优势度指数(D)采用Berger-Parker公式[15]:
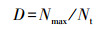
|
式中, Nmax:优势种的种群数量; Nt:所有种类的种群数量。
(4) 物种丰富度(R)采用Margalef公式[15]:
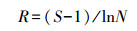
|
式中, S为物种数, N为个体数。
(5) 群落相似性系数采用Jaccard的相似性系数公式[13]:
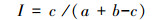
|
式中, a为A生境物种数, b为B生境物种数, c为A、B两生境共有的物种数。根据Jaccard的相似性系数原理, 当I为0—0.25时, 为极不相似; I为0.25—0.50时, 为中等不相似; I为0.50—0.75时, 为中等相似; I为0.75—1.00时, 为极相似。
3 结果和分析 3.1 青海玉树高原区蝶类群落特征 3.1.1 蝶类群落结构特征2016和2017年5月—8月, 共记录到蝶类个体数1580头, 62种, 隶属于7科40属(附表)。其中凤蝶科1种, 绢蝶科7种, 粉蝶科12种, 眼蝶科9种, 蛱蝶科12种, 灰蝶科16种, 弄蝶科5种。
| 种类 Specice |
数量Number of individuals | 总计 Total |
区系成分 Fauna composition |
||||
| 森林 Forest |
灌木 Shrub |
高寒灌丛 Alpine shrub |
高寒草甸草原 Alpine meadow |
裸岩 Naked rock |
|||
| 一、凤蝶科Papilionidae | |||||||
| 1金凤蝶Papilio machaon | 10 | 5 | 4 | 1 | 17 | 37 | W |
| 二、绢蝶科Parnassiidae | |||||||
| 2四川绢蝶Parnassius szechenyii | 24 | 24 | P | ||||
| 3君主绢蝶Parnassius imperator | 2 | 2 | W | ||||
| 4依帕绢蝶Parnassius epaphus | 1 | 25 | 26 | W | |||
| 5珍珠绢蝶parnassius orleans | 6 | 6 | P | ||||
| 6蓝精灵绢蝶Parnassius acdestis | 14 | 14 | P | ||||
| 7西猴绢蝶Parnassius simo | 20 | 20 | W | ||||
| 8元首绢蝶Parnassius cephalus | 4 | 4 | W | ||||
| 三、粉蝶科Pieridae | |||||||
| 9山豆粉蝶Colias montium | 1 | 3 | 4 | 8 | P | ||
| 10橙黄豆粉蝶Cloias fieldii | 1 | 1 | W | ||||
| 11黧豆粉蝶Colias richthofeni | 1 | 1 | 2 | 4 | P | ||
| 12大卫粉蝶Pieris davidis | 11 | 2 | 79 | 9 | 101 | P | |
| 13菜粉蝶Pieris rapae | 9 | 2 | 1 | 12 | W | ||
| 14妹粉蝶Mesapia peloria | 1 | 10 | 37 | 48 | W | ||
| 15箭纹绢粉蝶Aporia procris | 14 | 37 | 2 | 53 | W | ||
| 16暗色绢粉蝶Appria bieti | 1 | 1 | W | ||||
| 17马丁绢粉蝶Aporia martineti | 1 | 10 | 11 | O | |||
| 18酪色绢粉蝶Aporia potanini | 1 | 1 | P | ||||
| 19皮氏尖襟粉蝶Anthocharis bieti | 9 | 35 | 9 | 53 | W | ||
| 20库茨粉蝶Pieris kozlovi | 1 | 7 | 8 | P | |||
| 四、眼蝶科Satyridae | |||||||
| 21小型林眼蝶Aulocera sybillina | 2 | 2 | W | ||||
| 22蛇眼蝶Minois dryas | 10 | 7 | 17 | W | |||
| 23西门珍眼蝶Coenonympha semenovi | 2 | 7 | 9 | 18 | P | ||
| 24牧女珍眼蝶Coenonympha amaryllis | 3 | 5 | 20 | 3 | 31 | W | |
| 25斗毛眼蝶Lasiommata deidamia | 2 | 2 | W | ||||
| 26白瞳舜眼蝶Pararaguttata Bremeret | 1 | 27 | 28 | P | |||
| 27红眼蝶Erebia alcmena | 2 | 8 | 10 | 20 | W | ||
| 28古北拟酒眼蝶Paroeneis palaearctica | 10 | 18 | 15 | 24 | 67 | P | |
| 29阿芬眼蝶Aphantopus hyperanthus | 1 | 12 | 22 | 35 | P | ||
| 五、蛱蝶科Nymphalidae | |||||||
| 30镁斑豹蛱蝶Speyeria clara | 5 | 8 | 8 | 12 | 33 | P | |
| 31银斑豹蛱蝶Speyeria aglaja | 3 | 2 | 10 | 3 | 29 | 47 | P |
| 32龙女宝蛱蝶Boloria pales | 1 | 7 | 42 | 50 | W | ||
| 33菌网蛱蝶Melitaea agar | 4 | 20 | 7 | 17 | 48 | P | |
| 34黑网蛱蝶Melitaea jezabel | 3 | 2 | 3 | 8 | W | ||
| 35网蛱蝶Melitaea protomedia | 5 | 5 | P | ||||
| 36白钩蛱蝶Polygonia c-album | 1 | 3 | 4 | P | |||
| 37荨麻蛱蝶Aglais urticae | 7 | 21 | 2 | 11 | 5 | 46 | W |
| 38拉达麻蛱蝶Aglais ladakensis | 2 | 2 | 4 | 23 | 31 | W | |
| 39珍蛱蝶Clossiana gong | 22 | 2 | 26 | 50 | W | ||
| 40曲斑珠蛱蝶Issoria eugenia | 18 | 34 | 35 | 108 | 17 | 212 | W |
| 41缕蛱蝶Pieris kozlovi | 1 | 1 | W | ||||
| 六、灰蝶科Lycaenidae | |||||||
| 42李梳灰蝶Ahlbergia leei | 2 | 2 | 1 | 5 | P | ||
| 43枯灰蝶Cupido minimus | 7 | 2 | 2 | 11 | P | ||
| 44胡麻霾灰蝶Maculinea teleia | 1 | 3 | 4 | P | |||
| 45庞呃灰蝶Lycaena pang | 8 | 14 | 1 | 23 | W | ||
| 46红灰蝶Lycaena phlaeas | 4 | 2 | 5 | 1 | 12 | W | |
| 47喇灿灰蝶Agriades lamasem | 10 | 7 | 9 | 26 | W | ||
| 48婀灰蝶Albulina gaborronkayi | 33 | 23 | 54 | 54 | 21 | 185 | W |
| 49安婀灰蝶Albulina amphirrhoe | 7 | 7 | O | ||||
| 50多眼灰蝶Polyommatus eros | 2 | 3 | 5 | P | |||
| 51维纳斯眼灰蝶Polyommatus venus | 10 | 1 | 2 | 13 | P | ||
| 52丽罕莱灰蝶Helleia li | 2 | 2 | 4 | O | |||
| 53青海红珠灰蝶Lycaeides qinghaiensis | 19 | 8 | 2 | 29 | P | ||
| 54珞灰蝶Scolitantides orion | 5 | 1 | 6 | P | |||
| 55烂僖灰蝶Sinia lanty | 1 | 1 | W | ||||
| 56大紫琉璃灰蝶Celastrina oreas oreas | 32 | 32 | W | ||||
| 57埃灰蝶Eumedonia eumedon | 12 | 12 | P | ||||
| 七、弄蝶科Hesperiidae | |||||||
| 58小赭弄蝶Ochlodes venata | 1 | 1 | W | ||||
| 59克里银弄蝶Carterocephalus | 1 | 2 | 3 | P | |||
| 60北方花弄蝶Pyrgus alveus | 1 | 2 | 3 | W | |||
| 61三纹花弄蝶Pyrgus dejeani | 1 | 7 | 8 | W | |||
| 62黄点银弄蝶 Carrerocephalus flavomaculatus |
1 | 1 | W | ||||
| 合计The total | 227 | 333 | 408 | 302 | 310 | 1580 | |
| W:广布种,the widely distributed species;P:古北种,the palaearctic species;O:东洋种,the oriental species | |||||||
根据调查结果, 统计各科蝴蝶群落数量特征的各项指数(表 1)。从物种的组成看, 灰蝶科>蛱蝶科=粉蝶科>眼蝶科>绢蝶科>弄蝶科>凤蝶科; 从个体数量看, 蛱蝶科>灰蝶科>粉蝶科>眼蝶科>绢蝶科>凤蝶科>弄蝶科, 多样性指数排序为:灰蝶科>眼蝶科>蛱蝶科>粉蝶科>弄蝶科>绢蝶科>凤蝶科; 均匀度指数排序为:眼蝶科>绢蝶科>弄蝶科>蛱蝶科>粉蝶科>灰蝶科>凤蝶科; 优势度指数排序为:绢蝶科>眼蝶科>粉蝶科>蛱蝶科>灰蝶科>弄蝶科>凤蝶科; 物种丰富度指数依次为:灰蝶科>粉蝶科>绢蝶科>蛱蝶科>弄蝶科>眼蝶科>凤蝶科。
| 科名 Families |
属数 Genus |
种数 Species |
个体数 Individuals |
多样性指数 Diversity index(H′) |
均匀度指数 Evenness index(J) |
优势度指数 Dominance index(D) |
物种丰富度指数 Specific richness(R) |
||
| H′(GS) | H′(G) | H′(S) | |||||||
| 凤蝶科Papilionidae | 1 | 1 | 37 | 0 | 0 | 0 | 0 | 1 | 0 |
| 绢蝶科Parnassiidae | 1 | 7 | 96 | 1.6943 | 0 | 1.6943 | 0.8315 | 3.8400 | 2.1747 |
| 粉蝶科Pieridae | 5 | 12 | 301 | 3.3008 | 1.4735 | 1.8273 | 0.7354 | 2.9802 | 2.3844 |
| 眼蝶科Satyridae | 8 | 9 | 220 | 3.9423 | 2.0431 | 1.8992 | 0.8643 | 3.2836 | 1.9026 |
| 蛱蝶科Nymphalidae | 8 | 12 | 535 | 3.9217 | 1.9792 | 1.9425 | 0.7817 | 2.5236 | 2.0535 |
| 灰蝶科Lycaenidae | 14 | 6 | 375 | 4.5207 | 2.5993 | 1.9214 | 0.6930 | 2.0271 | 2.8734 |
| 弄蝶科Hesperiidae | 3 | 5 | 16 | 2.3757 | 1.0549 | 1.3208 | 0.8207 | 2 | 1.9236 |
| H′(F):科级多样性指数;H′(G):属级多样性指数;H′(S):种级的多样性指数 | |||||||||
从世界地理区系看, 该地区古北界19种, 占总种数的30%, 东洋界2种, 占总种数的3%, 两界共有种(广布种)41种, 占总种数的67%;从中国地理区划来看, 多区共有种占大多数, 单一区种类较少, 青藏区特有成分12种, 占总数的19%。玉树市在中国动物地理区划上属于古北界青藏区, 但其蝴蝶区系组成上古北种仅占30%, 广布种以67%占绝对优势, 这与其特殊的地理位置和复杂的生境有着密切的关系, 该区位于古北界青藏区的边缘, 靠近东洋界的西南区, 且境内的通天河峡谷作为一条天然通道, 为少数环境适应能力较强的蝶类创造了渗透北移的条件。除此之外, 该地区的气候虽然寒冷, 但蝴蝶种类相较于典型的青藏区内陆更为丰富, 与该区纬度较低,有相邻较低海拔的蝶类种类的渗透有关。
3.2 不同生境类型的蝶类群落与多样性 3.2.1 不同生境类型的蝶类群落特征根据调查结果, 统计生境蝴蝶群落数量特征的各项指数(表 2), 高寒灌丛生境在个体数量上最多。从物种组成上看, 灌木=高寒灌丛>森林>裸岩>高寒草甸草原; 从个体数量看, 高寒灌丛>灌木>裸岩>高寒草甸草原>森林; 多样性指数依次为灌木>森林>高寒灌丛>裸岩>高寒草甸草原; 均匀度指数依次为裸岩>森林>高寒灌丛>灌木>高寒草甸草原; 优势度指数依次为高寒草甸草原>高寒灌丛>森林>裸岩>灌木; 物种丰富度指数依次为灌木>高寒灌丛>森林>裸岩>高寒草甸草原。
| 生境 Habitat |
科数 Family |
属数 Genus |
种数 Species |
个体数 Individuals |
多样性指数 Diversity index(H′) |
均匀度指数 Evenness index(J) |
优势度指数 Dominance index(D) |
物种丰富度指数 Specific Richness (R) |
||
| H′(GS) | H′(G) | H′(S) | ||||||||
| 森林Forest | 6 | 27 | 34 | 227 | 6.2793 | 3.2409 | 3.0383 | 0.8616 | 0.1454 | 6.0830 |
| 灌木Shrub | 6 | 29 | 38 | 318 | 6.3156 | 3.3319 | 2.9837 | 0.8202 | 0.1159 | 6.4178 |
| 高寒灌丛 Alpine shrub |
6 | 29 | 38 | 423 | 6.2758 | 3.2817 | 2.9941 | 0.8231 | 0.1868 | 6.1184 |
| 高寒草甸草原 Alpine meadow |
7 | 14 | 19 | 302 | 4.6873 | 2.5796 | 2.1076 | 0.7158 | 0.3576 | 3.1521 |
| 裸岩Naked rock | 7 | 15 | 23 | 310 | 5.2977 | 2.4645 | 2.8332 | 0.9036 | 0.1355 | 3.8351 |
| H′(F):科级多样性指数;H′(G):属级多样性指数;H′(S):种级的多样性指数 | ||||||||||
由表 2可知, 灌木生境蝶类多样性指数、均匀度指数、物种丰富度指数都比较高, 由于该生境内灌木和林下植被种类丰富, 水源充足, 沟内狭窄,气候温暖, 相较于其他生境存在有较多的蝴蝶寄主植物和蜜源植物。灌木生境的优势度指数最低,优势种不明显,表明其生境内的自我调节能力强,适合多种蝶类的生存,蝴蝶种类丰富。
高寒草甸草原的多样性指数、均匀度指数、物种丰富度指数都比其他生境低, 该生境内海拔较高, 是为蝴蝶生长发育的主要制约因素。优势度指数高于其他生境, 表明在该生境中蝴蝶种类和数量分布不均匀, 生境内自身调节能力弱, 抗外界干扰能力差。
裸岩生境在5种生境中的海拔最高, 该生境内蝶类属数和种数都较少, 并且多样性指数、物种丰富度指数、优势度指数都较低, 主要是因为该生境海拔较高、温度低、植物种类稀少, 只适合少数高海拔蝶类生存。
3.2.2 不同生境蝶类群落相似性分析不同生境蝴蝶分布的相似性系数中(表 3), 森林生境与灌木、高寒灌丛、高寒草甸草原、裸岩生境, 高寒灌丛与高寒草甸草原和裸岩生境的物种相似性系数都在0.25—0.05之间, 为中等不相似。灌木生境和高寒草甸草原、裸岩生境, 高寒草甸草原和裸岩生境的物种相似性系数都在0.50—0.75之间, 为中等相似。灌木生境和高寒灌丛的物种相似性系数在0—0.25之间, 为极不相似。
| 生境 Habitat |
森林 Forest |
灌木 Shrub |
高寒灌丛 Alpine shrub |
高寒草甸草原 Alpine meadow |
裸岩 Naked rock |
| 森林Forest | 12 | 9 | 11 | 15 | |
| 灌木Shrub | 0.2667 | 9 | 24 | 27 | |
| 高寒灌丛Alpine shrub | 0.2727 | 0.1731 | 10 | 14 | |
| 高寒草甸草原Alpine meadow | 0.2619 | 0.5 | 0.2127 | 25 | |
| 裸岩Naked rock | 0.3571 | 0.551 | 0.2978 | 0.5319 |
由表 3可知, 灌木和高寒灌丛的物种相似性为极不相似, 这主要是由于二者海拔差距较大, 植被组成各不相同, 而且在空间距离上也相隔较远。高寒草甸草原和裸岩的物种相似性为中等相似, 这主要是因为二者海拔高度较相近, 并且植物分布有相似。
3.2.3 不同生境蝶类群落的月季变化特征对各月份蝴蝶个体数、种数、多样性指数、丰富度指数进行时序动态分析(图 1、图 2)。7月份蝴蝶的种类最多, 共40种, 5月份蝴蝶的种类最少, 共15种, 个体数的高峰出现在8月份, 共495头, 5月份蝶类个体数最低, 共116头。多样性指数从5—7月逐渐升高, 并且在7月达到最高点, 随后逐渐下降, 物种丰富度指数从5—7月逐渐上升, 并且在7月达到最高点, 随后逐渐下降。蝴蝶的多样性指数和物种丰富度指数都在7月份达到最高值。这种变化趋势与当地的气候变化相符, 5月份玉树地区温度较低, 活动的蝴蝶大部分是以成虫越冬、或以老龄幼虫越冬的种类, 因此蝴蝶的种类和数量都较少。6月份温度上升, 大部分植物进入展叶期, 以蔷薇科及小檗科等植物为寄主的箭纹绢粉蝶和以十字花科植物为寄主的皮氏尖襟粉蝶和大卫粉蝶在此时羽化成蝶。7月份是玉树市温度最适合植物生长的季节, 蝴蝶的丰富度达到最高。此时植物进入花期, 生长繁茂, 开花植物会吸引婀灰蝶、珍蛱蝶等访花蝴蝶并为它们提供食物来源。8月份温度降低,气温的变化对蜜源植物和寄主植物都有影响,蝴蝶成虫逐渐减少,开始以不同虫态越冬。

|
| 图 1 玉树市蝴蝶物种数和个体数的时间动态 Fig. 1 Temporal dynamics of the number of the butterfly species and individuals in Yushu city |

|
| 图 2 玉树市蝴蝶多样性指数和丰富度指数的时间动态 Fig. 2 Temporal dynamics of diversity index and richness of butterfly in Yushu city |
玉树市以其独特的地理位置、复杂多样的地形地貌和丰富的植物资源, 为蝶类提供了良好的生存环境。陈振宁等[5]在玉树地区蝴蝶调查中记录到蝶类7科30属47种, 而本次研究记录到蝶类7科40属62种, 从各科蝴蝶的种数来说, 本次研究所调查到的粉蝶科、灰蝶科、弄蝶科蝴蝶种类比陈振宁等[5]所记录种类增多, 本次调查新记录到蝴蝶34种。
4.2 蝶类群落结构特征由附表可见, 粉蝶科的大卫粉蝶(Pieris davidis)、皮氏尖襟粉蝶(Anthocharis bieti), 眼蝶科的古北拟酒眼蝶(Paroeneis palaearctica)、蛱蝶科的曲斑珠蛱蝶(Issoria eugenia)、灰蝶科的婀灰蝶(Albulina gaborronkayi)在本地区分布较普遍, 凤蝶科仅有金凤蝶(Papilio machaon)1种, 绢蝶作为高海拔地区分布的代表种, 在该地区分布较多, 弄蝶科蝴蝶种类及数量都较少; 灌木生境蝴蝶种类较多, 裸岩生境和高寒草甸草原蝴蝶种类较少, 这与该地区的海拔高度有关, 各生境中蝶类群落的物种数总体上随着海拔的逐渐升高呈逐渐下降趋势, 多样性指数也呈现出较明显的下降趋势, 这与张鑫等[16]、陈振宁等[5]的研究结果相符。
4.3 不同生境蝶类多样性植物及其环境是蝴蝶赖以生存的基础,也是蝴蝶的食物来源,同时由于对寄主植物的依赖性,蝴蝶的分布以寄主植物为中心,因此植被丰富程度和植物种类组成是影响蝶类群落的重要因素[17-18]。
不同生境蝶类物种丰富度的顺序为:灌木>高寒灌丛>森林>裸岩>高寒草甸草原。通过对不同生境蝴蝶种类和数量的比较, 发现蝴蝶种类和数量与生境类型的复杂程度呈正相关, 即生境越复杂, 蝴蝶种类和数量越多, 这与陈振宁等[19]、王敏等[20]研究的结果相符。
4.4 蝶类多样性保护玉树地区独特的地理位置, 使其拥有丰富的动植物资源, 同时也为蝶类提供了适宜的栖息环境, 有利于蝶类种群发展。近些年随着雪线上移, 部分分布在高寒草甸草原生境的蝶类向高海拔的裸岩生境迁移, 使裸岩生境的物种丰富度增加, 而高寒草甸草原生境的物种多样性减少, 有研究表明, 物种丰富度与海拔高度呈峰状关系[21-22], 如果低海拔物种的迁移不能够补偿高海拔物种的减少, 那么低海拔物种向高海拔迁移最终导致整个地区的物种丰富度的下降[23], 为了降低气候变化对蝴蝶生物多样性的影响, 应该加强气候变化对生物多样性具体影响的研究, 并积极寻求适应性应对办法, 以减少气候变化带来的负面影响。
昆虫是生态系统中的重要组成部分, 蝴蝶成虫可为植物授粉, 幼虫主要寄生在植物表面, 其整个生活史都与植物密切相关。蝴蝶类群分布格局的变化会导致其与寄主植物之间协同关系的改变, 为了有效的对其进行保护, 需要增强对蝴蝶物种的监测, 减少本地物种的高海拔迁移现象, 有效的保护当地的蝶类资源。
| [1] | 李志刚, 李军, 张碧胜, 龚鹏博, 韩诗畴. 广州市蝴蝶群落结构与多样性. 生态学杂志, 2008, 27(6): 1047–1050. |
| [2] | 马方舟, 徐海根, 丁晖, 周旭, 张彦静, 贾凤海. 我国蝴蝶多样性的保护现状与对策. 世界环境, 2016(S1): 29–31. |
| [3] | 武素功, 冯祚建. 青海可可西里地区生物与人体高山生理. 北京: 科学出版社, 1996. |
| [4] | 蔡振声, 史先鹏, 徐培河. 青海经济昆虫志. 西宁: 青海人民出版社, 1994. |
| [5] | 陈振宁, 刘长海, 曾阳, 张明庆. 青海玉树地区蝶类及其区系分析. 四川动物, 2000, 19(4): 229–232. DOI:10.3969/j.issn.1000-7083.2000.04.008 |
| [6] | 周尧. 中国蝶类志(上、下册). 郑州: 河南科学技术出版社, 1994. |
| [7] | 武春生, 徐堉峰. 中国蝴蝶图鉴(全4册). 福州: 海峡书局, 2017. |
| [8] | 王敏, 范骁凌. 中国灰蝶志. 郑州: 河南科学技术出版社, 2002. |
| [9] | 武春生. 中国动物志·昆虫纲(第二十五卷)鳞翅目·凤蝶科. 北京: 科学出版社, 2001. |
| [10] | 武春生. 中国动物志·昆虫纲(第五十二卷)鳞翅目·粉蝶科. 北京: 科学出版社, 2010. |
| [11] | 袁锋, 袁向群, 薛国喜. 中国动物志·昆虫纲(第五十五卷)鳞翅目·弄蝶科. 北京: 科学出版社, 2015. |
| [12] | Lang S Y. The Nymphalidae of China (Lepidoptera, Rhopalocera), Volume 1: Libytheinae, Danainae, Calinaginae, Morphinae, Heliconiinae, Nymphalinae, Charaxinae, Apaturinae, Cyrestinae, Biblidinae, Limenitinae. Pardubice, Czech Republic: Tshikolovets Publications, 2012. |
| [13] | 马克平. 生物群落多样性的测度方法:Ⅰ-α多样性的测度方法(上). 生物多样性, 1994, 2(3): 162–168. DOI:10.3321/j.issn:1005-0094.1994.03.007 |
| [14] | 马克平, 刘玉明. 生物群落多样性的测度方法:Ⅰ-α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231–239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
| [15] | 赵志模, 郭依泉. 生境类型生态学原理和方法. 重庆: 科学技术出版社重庆分社, 1990: 147–172. |
| [16] | 张鑫, 胡红英, 吕昭智. 新疆东部天山蝶类多样性及其垂直分布. 生态学报, 2013, 33(17): 5329–5338. |
| [17] | Weibull A C, Bengtsson J, Nohlgren E. Diversity of butterflies in the agricultural landscape:The role of farming system and landscape heterogeneity. Ecography, 2000, 23(6): 743–750. DOI:10.1111/j.1600-0587.2000.tb00317.x |
| [18] | Vu L V. Diversity and similarity of butterfly communities in five different habitat types at Tam Dao National Park, Vietnam. Journal of Zoology, 2009, 277(1): 15–22. DOI:10.1111/jzo.2009.277.issue-1 |
| [19] | 陈振宁, 曾阳. 青海祁连地区不同生境类型蝶类多样性研究. 生物多样性, 2001, 9(2): 109–114. DOI:10.3321/j.issn:1005-0094.2001.02.003 |
| [20] | 王敏, 黄国华, 范骁凌, 谢国忠, 黄林生, 戴克元. 石门台自然保护区蝴蝶物种多样性研究. 生物多样性, 2003, 11(6): 441–453. DOI:10.3321/j.issn:1005-0094.2003.06.002 |
| [21] | Rahbek C. The elevational gradient of species richness:a uniform pattern?. Ecography, 1995, 18(2): 200–205. DOI:10.1111/eco.1995.18.issue-2 |
| [22] | McCain C M. Elevational gradients in diversity of small mammals. Ecology, 2005, 86(2): 366–372. DOI:10.1890/03-3147 |
| [23] | Wilson R J, Gutiérrez D, Gutiérrez J, Monserrat V J. An elevational shift in butterfly species richness and composition accompanying recent climate change. Global Change Biology, 2007, 13(9): 1873–1887. DOI:10.1111/gcb.2007.13.issue-9 |