 2018, Vol. 38
2018, Vol. 38文章信息
- 马少薇, 刘果厚, 刘利红, 张岑, 刘冠志, 郭娇, 王蕾, 鄂佳庆.
- MA Shaowei, LIU Guohou, LIU Lihong, ZHANG Cen, LIU Guanzhi, GUO Jiao, WANG Lei, E Jiaqing.
- 浑善达克沙地黄柳种群性比及空间分布
- Sex ratio and spatial distribution of Salix gordejevii in Hunshandake Sandy Land
- 生态学报. 2018, 38(21): 7708-7717
- Acta Ecologica Sinica. 2018, 38(21): 7708-7717
- http://dx.doi.org/10.5846/stxb201711021960
-
文章历史
- 收稿日期: 2017-11-02
- 网络出版日期: 2018-08-08





2. 正蓝旗草原工作站, 正蓝旗 027200;
3. 内蒙古农业大学沙漠治理学院, 呼和浩特 010019;
4. 内蒙古自治区林业监测规划院, 呼和浩特 010020
2. Grassland Workstation of Zhenglan Banner, Zhenglan Banner 027200, China;
3. College of Desert Control Science and Engineering, Inner Mongolia Agricultural University, Hohhot 010019, China;
4. Monitoring & Planning Institute of Inner Mongolia Forestry Administration, Hohhot 010020, China
雌雄异株植物是指雌花和雄花分别生长在不同植株上的植物, 尽管该类植物在24万被子植物中仅占6%(约14620种), 但其所属科的数量却占被子植物门总科数的40%, 是地球陆地生态系统不可或缺的组成部分[1-2]。自1930年Fisher首次提出当雌株和雄株对后代的资源投资相等时种群性比应该是平衡[3]的观点后, 针对雌雄异株植物的研究就吸引了大家越来越多的注意。大量研究表明, 与雄株相比, 雌株会在生殖方面投资更多的资源以孕育种子[4], 造成种群性比出现不同程度的失衡, 通常以雄株多于雌株为主, 但也有一些物种性比偏雌。如Barrett等基于前人研究统计了126种雌雄异株植物(隶属于47科)的种群性比, 发现有67%的物种出现了性比偏倚, 其中性比偏雄性的物种约为偏雌性的2倍[5]。Lei等对贡嘎山高海拔地区2种杨柳科优势植物冬瓜杨(Populus purdomii)和川滇柳(Salix rehderiana)进行性比调查, 发现冬瓜杨性比偏雄、川滇柳性比偏雌[6]。雌雄异株植物性比偏倚是由其生长、繁殖、适应环境策略的不同所致, 性比偏倚反过来会通过影响植株生殖分配, 在更大程度上造成性别差异[7]。雌雄异株植物在空间分布中普遍呈非随机性分布, 即性别空间分离[8]。如赵亚洲等发现太行山南端黄连木(Pistacia chinensis)种群雌株和雄株在空间上相互排斥, 存在性别空间分离现象[9]。资源匮乏是导致性别空间分离的主要原因, 为了更好的适应微生境, 雌株和雄株趋向于利用不同的环境和资源, 以减少种内竞争。在长期进化中, 两性植株极有可能对环境形成了不同的适应策略[10-14], 这最终将影响种群的性比组成、结构和空间分布等[15]。由此可见, 性比和空间分布格局对雌雄异株植物的繁殖策略、种群进化及发展起着至关重要的作用。
黄柳(Salix gordejevii Y. L. Chang et Skv.)是杨柳科(Salicaceae)柳属(Salix)的旱中生灌木, 具有生长迅速、耐沙埋、耐干旱等特性, 是流动、半流动沙丘的建群种或优势种, 甚至可以在沙地上形成纯群落[16-17]。因其较强的适应性和显著的防风固沙效益, 被誉为治理流沙的“活沙障”[18], 是京津风沙源治理区首选推广的灌木树种之一, 在改善沙区生态环境、逆转荒漠化过程中起着不可替代的作用。我们在营造黄柳人工林调查其成活率、保存率以及后期抚育的过程中发现, 雌性个体数量往往大于雄性个体数量, 由此引发了我们对天然种群调查的兴趣, 结果显示在天然种群中性比亦是如此。近年来, 关于黄柳的研究工作主要集中在营养器官解剖结构[16-17]、光合特性日变化[19]以及遗传多样性[20]等方面, 而从雌雄异株植物角度出发的研究还较为少见。同时, 浑善达克沙地地处我国北方半旱、半干旱地区, 降水量少、风沙大、土壤肥力低、资源异质性等恶劣环境既会影响天然种群性比和空间分布特征, 最终导致种群呈衰退趋势, 也不利于营造稳定的防风固沙林。因此, 本研究以黄柳天然种群为研究对象, 从种群性比和空间分布格局角度进行研究, 旨在解决以下问题(1)天然种群性比是否偏离1:1;(2)雌株和雄株是否出现性别空间分离现象。研究结果不仅有助于深化了解种群自身特性、发展动态以及与环境间的关系, 还可为黄柳在植被恢复治理中的性别选择、比例配用以及栽植方式和位置提供理论依据, 这对其种群建成和生态学效应最大化利用具有重要的实践意义。
1 材料与方法 1.1 研究区概况研究区位于内蒙古锡林郭勒盟正蓝旗, 地理位置为41°56′-43°11′ N, 115°00′-116°42′ E。该地区地处中温带大陆性季风气候区, 冬季寒冷漫长, 夏季温热短促。年平均温度1.7℃, 极端最高温度为35.9℃, 极端最低温度为-36.6℃, 平均日照时数3037 h, ≥10℃年积温为2000℃; 年平均降水量355 mm, 降雨季节分布不均, 主要集中于7-9月, 约占全年降雨量的80%-90%;年均蒸发量为1931.4 mm; 无霜期110 d; 年平均风速为4 m/s, 主要风向为西北风, 春秋季节风大沙多, 时有沙尘暴天气。该地区的土壤类型为栗钙土, 其上分布有风沙土。植被种类较为丰富, 木本植物主要有沙地榆(Ulmus pumila var. sabulosa)、黄柳、小红柳(Salix microstachya var. bordensis)、小叶锦鸡儿(Caragana microphylla)等, 草本植物主要有褐沙蒿(Artemisia halodendron)、羊草(Leymus chinensis)、星毛委陵菜(Potentilla acaulis)等[21]。
1.2 研究方法 1.2.1 样地设置黄柳虽然是浑善达克沙地流动、半流动沙丘的建群种或优势种, 但其分布连续性差, 普遍呈斑块性分布, 对大面积研究种群结构及动态发展带来了一定的困难。为了准确揭示黄柳雌雄群体数量比例、空间分布格局及空间关系, 2017年4月我们对浑善达克沙地进行了实地踏查, 根据其天然种群的实际分布情况随机选取了4个具有代表性的种群, 分别位于扎格斯台、饮马井、巴格来、宝绍岱, 具体位置见图 1。在每个地区分别设置1个50 m×50 m的样方, 对各样地的土壤含水量、土壤肥力指标以及伴生植物等环境因子进行了测定和调查(表 1)。

|
| 图 1 研究区的地理位置 Fig. 1 Location of study site |
| 样地 Plots | 地理位置 Geographical position | 海拔 Altitude/m | 土壤含水量 Soil moisture content/% | pH | 有机质 Organic matter/(g/kg) | 全氮 Total N/ (g/kg) | 全磷 Total P/ (g/kg) | 伴生植物 Associated plants |
| 扎格斯台 | 42°36′ N 116°14′ E | 1367 | 1.39±0.14b | 7.75±0.17a | 3.75±0.20a | 0.19±0.03a | 0.15±0.08a | 沙地榆、小红柳、小叶锦鸡儿等 |
| 饮马井 | 42°28′ N 115°59′ E | 1422 | 3.37±0.08a | 7.79±0.54a | 3.61±0.09b | 0.19±0.01a | 0.14±0.03a | 小红柳、褐沙蒿、羊草等 |
| 巴格来 | 42°31′ N 115°52′ E | 1330 | 1.13±0.06c | 7.14±0.71a | 2.11±0.60c | 0.07±0.03b | 0.14±0.05a | 小红柳、小花花旗杆 (Dontostemon micranthus)、 地梢瓜(Cynanchum thesioides)等 |
| 宝绍岱 | 42°37′ N 115°31′ E | 1242 | 0.58±0.06d | 6.85±0.15a | 2.27±0.65c | 0.11±0.02b | 0.10±0.01a | 兴安虫实(Corispermum chinganicum)、狗尾草 (Setaria viridis)等 |
为了准确的测量样方内黄柳个体的空间位置, 采用相邻格子法将每个样方进一步划分成25个10 m×10 m的亚样方, 以亚样方的东西方向为X轴、南北方向为Y轴, 以坐标值(距离)表示, 测量其空间位置, 在分析时再将每个个体的坐标值转换为在50 m×50 m的样方内所对应的坐标值。根据繁殖器官(柔荑花序)识别并记录所有植株的性别, 并测量其基径、株高和冠幅等特征。为了进一步分析不同阶段的性比格局, 根据刘冠志对黄柳丛生枝年龄结构的划分方法, 即以“径级(基径)代替龄级”[22], 并结合样方内雌雄个体的数量特征, 将样方内的雌雄个体按基径大小划分为3个径级:径级Ⅰ(基径 < 10 mm), 代表幼树和小树; 径级Ⅱ(10 mm≤基径 < 20 mm), 代表中树; 径级Ⅲ(基径≥20 mm), 代表大树和老树。
1.3 数据分析 1.3.1 形态特征及性比分析采用SPSS 22.0软件进行形态特征和性比分析。独立样本T检验用于比较各样地中雌雄群体在基径、株高和冠幅间的显著性程度; χ2检验性比(雌/雄)偏离1:1零假设的显著性程度。
1.3.2 空间格局分析基于种群分布的二维坐标点信息, 采用点格局分析中的单变量K(t)函数[23]和经过校正边缘效应的L(t)函数[24]来分析雌雄群体的空间分布格局。单变量K(t)函数的表达式为:
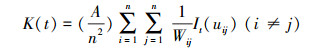
|
式中, t表示空间尺度; A表示样方面积; n表示样方内植物个体总数; I表示指示函数; uij表示点i和点j之间的距离, 当uij≤t时, It(uij)=1, 当uij﹥t时, It(uij)=0;Wij表示以点i为圆心, uij为半径的圆周长在面积A中的比例。为了使得方差更稳定、结果更可靠, Besag[24]对K(t)函数进行了修正, 得出了L(t)函数, 其表达式为:
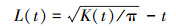
|
式中, 当L(t)=0时, 则在空间尺度t下的分布方式为随机分布; 当L(t)﹥0时, 则在空间尺度t下的分布方式为聚集分布; 当L(t)﹤0时, 则在空间尺度t下的分布方式为均匀分布。采用完全随机零假设来检验格局类型[25], 即使用随机点过程来检验分布偏离完全随机假设的显著性程度。
1.3.3 空间关系分析采用双变量K12(t)函数[26]和修正后的L12(t)函数[27]分析黄柳雌雄群体的空间关系。双变量K12(t)函数的表达式为:
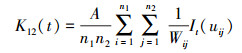
|
式中, n1和n2分别表示在面积A中不同性别的个体数, 其他参数含义同K(t)函数。同样, 为了使方差更稳定、结果更可靠, Diggle[27]对K12(t)函数进行了修订, 得出如下L12(t)函数:
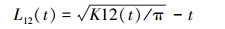
|
式中, 当L12(t)=0时, 则表示在空间尺度t下雌雄群体呈空间独立; 当L12(t)﹥0时, 则表示在空间尺度t下雌雄群体呈空间正相关(吸引); 当L12(t)﹤0时, 则表示在空间尺度t下雌雄群体呈空间负相关(排斥)。雌雄个体是在同等先决条件下生长的, 后天的空间过程才导致不同性别个体的分布及相互作用出现差异, 因此采用标签随机化假设进行双变量空间关系分析[28]。
点格局分析采用Programita(Wiegand, 2010版)软件进行, 其空间尺度t为0-25 m, 经100次Monte Carlo拟合检验计算上、下包迹线, 得到99%的置信区间[29]。当实际分布的L(t)值在包迹线之间时为随机分布, 在包迹线以上时为聚集分布, 在包迹线以下时为均匀分布。当L12(t)值在包迹线之间时表示空间独立, 当L12(t)值在包迹线之上时表示呈空间正相关(吸引), 当L12(t)在包迹线之下时表示空间负相关(排斥)[30]。
2 结果与分析 2.1 雌雄群体分布及植株形态大小图 2为各样方黄柳个体的空间分布图(50 m×50 m), 其中扎格斯台种群植株数量最多, 为106株(79:26:1, 雌株:雄株:无法确定性别的植株, 下同); 饮马井种群植株数量最少, 为55株(39:16:0);巴格来种群和宝绍岱种群植株数量居中, 分别为96株(65:28:3)和92株(65:22:5)。

|
| 图 2 各样方黄柳(Salix gordejevii)个体的空间分布图 Fig. 2 Spatial distribution map of S. gordejevii in different sampling plots |
如表 2所示, 从4个种群的总平均值来看, 黄柳雌株群体的基径和株高显著低于雄株群体(P < 0.001和P=0.036), 而雌雄群体在冠幅方面无显著差异(P=0.183)。在4个种群中, 扎格斯台种群雌株群体的基径和株高显著低于雄株群体(P < 0.001和P=0.008), 而冠幅无显著差异; 饮马井、巴格来和宝绍岱3个样地中雌株群体在基径、株高和冠幅的平均值上尽管均小于雄株群体, 但统计检验不显著。从种群间形态特征的比较结果来看, 宝绍岱种群中黄柳的雌、雄株群体的基径和株高最小, 而饮马井种群中的雌株群体和扎格斯台种群中的雄株群体则在上述两个指标上分别最大。上述形态特征的比较结果反映了黄柳雌雄群体间在形态上的差异与种群及其所处环境条件密切相关。
| 样地 Plots | 基径Basal diameter/mm | P | 株高Plant height/m | P | 冠幅Crown size/m2 | P | |||
| 雌株Females | 雄株Males | 雌株Females | 雄株Males | 雌株Females | 雄株Males | ||||
| 扎格斯台 | 15.27±6.07 | 21.68±7.60 | <0.001** | 1.71±0.44 | 2.06±0.59 | 0.008** | 5.12±4.13 | 5.99±4.27 | 0.369 |
| 饮马井 | 16.57±6.42 | 17.17±5.51 | 0.744 | 1.78±0.32 | 1.84±0.18 | 0.453 | 4.65±3.00 | 5.59±3.64 | 0.328 |
| 巴格来 | 14.16±5.28 | 16.54±7.32 | 0.128 | 1.69±0.36 | 1.65±0.36 | 0.606 | 4.06±4.42 | 3.59±3.28 | 0.614 |
| 宝绍岱 | 11.36±6.02 | 14.34±7.72 | 0.066 | 1.68±0.87 | 1.87±0.86 | 0.382 | 3.68±4.35 | 5.48±5.08 | 0.113 |
| 合计Total | 14.16±6.16 | 17.58±7.63 | <0.001** | 1.71±0.56 | 1.85±0.58 | 0.036* | 4.39±4.13 | 5.07±4.16 | 0.183 |
| P表示经独立样本T检验后的显著性概率, *表示雌雄间差异显著(P < 0.05), **表示雌雄间差异极显著(P < 0.01) | |||||||||
如表 3所示, 扎格斯台、饮马井、巴格来和宝绍岱种群的雌雄比例分别为3.04、2.44、2.32、2.95, χ2检验表明各种群性比均极显著偏雌性(P < 0.01)。扎格斯台、饮马井和巴格来种群雌株群体Ⅰ和Ⅲ级个体数量相差较小, 年龄结构均表现为稳定型; 而雄株群体Ⅰ级个体数量明显比Ⅲ级个体数量少, 年龄结构都表现为衰退型。宝绍岱种群雌株群体Ⅰ级和Ⅱ级个体数量占群体数量的84.62%, 表现增长型; 而雄株群体各径级个体数量相差不大, 表现为稳定型。为了进一步研究黄柳种群性比, 对每一径级雌雄群体的性比进行分析, 结果表明4个种群性比基本均为在第Ⅰ径级和第Ⅱ径级中表现出显著的偏雌性(P < 0.01), 第Ⅲ径级性比趋近于1:1。
| 径级 Diameter class | 扎格斯台 | 饮马井 | 巴格来 | 宝绍岱 | |||||||||||
| 雌株 Females | 雄株 Males | 性比 Sex ratio | 雌株 Females | 雄株 Males | 性比 Sex ratio | 雌株 Females | 雄株 Males | 性比 Sex ratio | 雌株 Females | 雄株 Males | 性比 Sex ratio | ||||
| Ⅰ | 14 | 2 | 7.00** | 8 | 1 | 8.00* | 14 | 6 | 2.33ns | 31 | 8 | 3.88** | |||
| Ⅱ | 48 | 10 | 4.80** | 17 | 8 | 2.13ns | 42 | 11 | 3.82** | 24 | 8 | 3.00** | |||
| Ⅲ | 17 | 14 | 1.21ns | 14 | 7 | 2.00ns | 9 | 11 | 0.82ns | 10 | 6 | 1.67ns | |||
| 合计Total | 79 | 26 | 3.04** | 39 | 16 | 2.44** | 65 | 28 | 2.32** | 65 | 22 | 2.95** | |||
| 径级Ⅰ(基径 < 10 mm)代表幼树和小树, 径级Ⅱ(10 mm≤基径 < 20 mm)代表中树, 径级Ⅲ(基径≥20 mm)代表大树和老树; *表示性比(雌/雄)显著偏离1:1(P < 0.05), **表示性比极显著偏离1:1(P < 0.01), ns表示性比不显著偏离1:1(χ2检验) | |||||||||||||||
对4个黄柳种群进行Ripley L(t)单变量空间格局分析(图 3), 结果表明扎格斯台种群全部植株在0-1 m尺度内表现为随机分布, 随着尺度的增加表现为聚集分布; 雌、雄群体均在0-4 m尺度内表现为随机分布, 4-25 m尺度内表现为聚集分布。饮马井种群全部植株和雌株群体分布格局一致, 即在0-1 m尺度内表现为随机分布, 1-25 m尺度内表现为聚集分布; 雄株群体则在0-14 m尺度内表现为随机分布, 14-25 m尺度内表现为聚集分布。巴格来种群和宝绍岱种群全部植株和雌、雄群体三者均表现为在0-25 m尺度内呈聚集分布。总体来看, 黄柳全部植株、雌株群体和雄株群体基本表现为聚集分布, 但部分种群或群体在小尺度内会表现为随机分布。

|
| 图 3 黄柳(S. gordejevii)的空间分布格局 Fig. 3 Spatial distribution pattern of S. gordejevii 有数据点的实线代表由实际数据计算所得的值, 无数据点的实线代表Monte Carlo拟合检验计算得出的上、下包迹线 |
采用标签随机化假设对雌雄群体的空间关系进行分析(图 4), 结果表明扎格斯台、饮马井、巴格来种群雌株和雄株的空间关系均表现为无空间相关性, 说明两性在空间分布上相互独立, 不存在资源竞争; 而宝绍岱种群两性的空间关系在全部尺度内表现为负相关, 说明雌株和雄株在空间分布上相互排斥, 对资源的需求存在竞争性。

|
| 图 4 黄柳(S. gordejevii)雌雄群体的空间关系 Fig. 4 Spatial relationships between male and female S. gordejevii populations |
性比是一个种群所有个体或某一龄级的个体中, 雌性和雄性个体的数目比例, 对种群的发展具有重要作用, 若两性个体数量相差过于悬殊, 则不利于种群增殖, 进而影响种群结构及其动态[31]。通过对浑善达克沙地4个种群雌雄植株个体数量分析得出, 黄柳种群性比极显著偏雌性(P < 0.01), 这与前人得出的柳属植物性比偏雌的观点相符。如Ueno等对龙江柳(Salix sachalinensis)3个种群的2992株个体进行了为期3年的观察, 发现其性比偏雌性[32]; Myers-Smith对加拿大地区5种柳属植物的性比进行研究也发现此规律[33]。雌雄群体对环境压力的响应差异是造成种群性比失衡的主要原因之一, 这种差异将直接决定雌雄之间的竞争和生存能力的大小。如在干旱胁迫下青杨(Populus cathayana)雄株具有较好的适应能力和保护机制, 而雌株的生长发育和生理生化过程更易遭到破坏[10, 34], 这种雄株对环境的适应能力强于雌株的差异就造成了其天然种群性比通常偏雄。柳属和杨属(Populus)同隶属于杨柳科, 但性比偏倚方向却完全相反, 即柳属植物性比偏雌。Jiang等以康定柳(Salix paraplesia)为代表植物对该现象进行了研究, 发现柳属植物在受到环境抑制时, 雌株会通过补偿生长将资源更多地投入于生长和繁殖, 表现出比雄株更强的抵御不良环境的能力[11], 所以柳属植物种群性比偏雌。这也说明柳属和杨属植物在性比偏倚方向上出现差异可能和二者在环境胁迫下雌雄植株表现出的不同适应机制有关。
对不同径级植株的性比分析表明, 第Ⅰ和第Ⅱ径级的雌雄个体数量是决定黄柳种群性比偏倚的主要组成部分, 而随着植株径级的增长性比逐渐趋近于1:1, 说明雌性的幼树、小树及中树的存活率高于雄性, 而大树和老树的存活率相似, 这与Myers-Smith等[33]的研究结果一致。环境条件是继种子萌发后决定幼苗存活和生长的主要因素, 崔秀萍等认为浑善达克沙地环境恶劣, 沙面温度高、土壤含水量低等环境条件对幼苗的存活和生长属于强烈的逆境[35], 而雌雄异株植物通常在逆境下表现出不同的适应策略[2], 因此雌性第Ⅰ和第Ⅱ径级的个体数量多于雄性是由于雌株较雄株对恶劣环境具有更强的适应性, 从而表现出更多的存活数量; 雌株较雄株具有更高的繁殖成本, 随着年龄的增加, 雌株较高的繁殖投资造成死亡率高于雄株, 性比由原本的偏雌性逐渐过渡为趋近于1:1, 故第Ⅲ径级的雌雄性比接近于平衡。这也是雌株群体年龄结构多表现为稳定型或增长型, 而雄株群体多表现为衰退型的原因。
3.2 雌雄群体的空间分布及空间关系种群分布格局是物种生物学特性、种间关系以及环境条件综合作用的表现, 一般可以概括为3种分布类型, 即随机分布、均匀分布和聚集分布[31]。黄柳种群空间格局(全部植株、雌株群体和雄株群体)均表现出明显的聚集分布, 研究表明聚集分布是植物种群最普遍的分布方式, 这是由于在自然情况下, 植物种子散布或无性繁殖体传播多是在母株附近, 加上环境异质性以及群落中植物种间关系的发展, 都会导致不同植物种在群落中占据其适合的生境, 从而形成聚集分布格局[15, 36]。结合黄柳自身的繁殖特性来看, 尽管种子以风媒传播, 具有远距离散布能力和次级散布能力, 但从种子萌发后到第二年幼苗保存率仅为18.3%, 天然更新困难[21], 这就使得种群在空间上很难呈现随机分布。然而黄柳克隆繁殖能力很强, 加之所处环境资源普遍呈斑块状分布, 这些因素都有利于雌、雄群体形成聚集分布。这一结果支持了潘春芳等对山杨(Populus davidiana)的研究结论[36]。性别空间分离现象仅出现在宝绍岱种群, 而其他3个种群雌雄群体的空间关系均表现为相互独立, 这种性别空间分离仅出现在部分种群的现象与前人的研究结果一致, 如Ueno等发现在日本宫城县附近分布的龙江柳种群存在性别空间分离现象[37], 而分布在北海道石狩川流域附近的种群却未出现此现象, 他认为这是环境资源的异质性所致[32]。根据黄柳种群所处位置的环境因子来看, 宝绍岱相较于其他地区环境更为恶劣, 土壤含水量显著低于其他3个种群, 全氮和有机质含量显著低于扎格斯台和饮马井种群(表 1), 同时结合其分布情况来看, 雌雄个体生长紧密, 仅约分布在调查样地内(50 m×50 m)的1/2区域(图 2), 雌雄个体对土壤水分、养分以及光照等资源的竞争更加强烈, 最终导致空间分离。
综上所述, 本研究对黄柳种群性比和空间分布格局以及空间关系进行了初步探究, 发现其种群性比极显著偏雌性(P < 0.01), 分析原因可能与雌株群体比雄株群体能够更好的适应浑善达克沙地的气候有关, 具体原因需进行进一步深入研究。黄柳雌、雄群体均呈现明显的聚集性分布; 除宝绍岱种群外, 其他种群的雌雄群体关系均为空间独立, 不同性别的个体可以占据不同的微环境, 资源竞争不激烈, 这被认为是雌雄异株植物的进化优势。此外, 雌雄异株植物种群的繁殖更新受性比及分布格局的影响, 因此在使用黄柳人工造林时应考虑雌雄个体的数量比例和空间位置, 这对其种群建成、稳定和发展以及生态效益的最大化发挥具有重要的实践意义。
| [1] | Renner S S, Ricklefs R E. Dioecy and its correlates in the flowering plants. American Journal of Botany, 1995, 82(5): 596–606. DOI:10.1002/j.1537-2197.1995.tb11504.x |
| [2] | 陈娟, 李春阳. 环境胁迫下雌雄异株植物的性别响应差异及竞争关系. 应用与环境生物学报, 2014, 20(4): 743–750. |
| [3] | Fisher R A. The Genetical Theory of Natural Selection. Oxford: Oxford University Press, 1930. |
| [4] | Obeso J R. The costs of reproduction in plants. New Phytologist, 2002, 155(3): 321–348. DOI:10.1046/j.1469-8137.2002.00477.x |
| [5] | Barrett S C H, Yakimowski S B, Field D L, Pickup M. Ecological genetics of sex ratios in plant populations. Philosophical Transactions of the Royal Society of London B:Biological Sciences, 2010, 365(1552): 2549–2557. DOI:10.1098/rstb.2010.0002 |
| [6] | Lei Y B, Chen K, Jiang H, Yu L, Duan B L. Contrasting responses in the growth and energy utilization properties of sympatric Populus and Salix to different altitudes:implications for sexual dimorphism in Salicaceae. Physiologia Plantarum, 2017, 159(1): 30–41. DOI:10.1111/ppl.2017.159.issue-1 |
| [7] | 宋海凤, 张胜. 杨柳科植物随环境变化的性别响应差异. 山地学报, 2017, 35(5): 645–652. |
| [8] | Bierzychudek P, Eckhart V. Spatial segregation of the sexes of dioecious plants. The American Naturalist, 1988, 132(1): 34–43. DOI:10.1086/284836 |
| [9] | 赵亚洲, 辛雅芬, 马钦彦, 张春雨. 雌雄异株树种黄连木种群性比及空间分布. 生态学杂志, 2010, 29(6): 1087–1093. |
| [10] | Xu X, Peng G Q, Wu C C, Korpelainen H, Li C Y. Drought inhibits photosynthetic capacity more in females than in males of Populus cathayana. Tree Physiology, 2008, 28(11): 1751–1759. DOI:10.1093/treephys/28.11.1751 |
| [11] | Jiang H, Zhang S, Lei Y B, Xu G, Zhang D. Alternative growth and defensive strategies reveal potential and gender specific trade-offs in dioecious plants Salix paraplesia to nutrient availability. Frontiers in Plant Science, 2016, 7: 1064. |
| [12] | 胥晓. 植物根系识别的研究. 西华师范大学学报:自然科学版, 2016, 37(4): 365–369. |
| [13] | 竺诗慧, 董廷发, 刘刚, 肖娟, 朱娟, 曾贞, 陈德甫, 胥晓. 桑树(Morus alba)幼苗根系分泌物对雌雄植株生长发育的影响. 植物生理学报, 2016, 52(1): 134–140. |
| [14] | 朱娟, 刘刚, 肖娟, 竺诗慧, 曾贞, 胥晓, 董廷发. 桑树不同性别组合种植模式下的生物量. 生态学杂志, 2016, 35(9): 2336–2340. |
| [15] | 王志峰, 胥晓, 李霄峰, 杨鹏, 袁新利. 青杨雌雄群体沿海拔梯度的分布特征. 生态学报, 2011, 31(23): 7067–7074. |
| [16] | 崔秀萍, 刘果厚, 张瑞麟. 浑善达克沙地不同生境下黄柳叶片解剖结构的比较. 生态学报, 2006, 26(6): 1842–1847. DOI:10.3321/j.issn:1000-0933.2006.06.027 |
| [17] | 刘冠志, 刘果厚, 兰庆, 李红颖, 曹瑞, 王健, 刘利红. 黄柳与小红柳导管分子形态特征及其生态适应性比较研究. 西北植物学报, 2016, 36(2): 316–322. |
| [18] | 张瑞麟.浑善达克沙地黄柳活沙障的设置及防风固沙作用研究[D].呼和浩特: 内蒙古农业大学, 2007. http://www.cnki.com.cn/Article/CJFDTotal-ZLDF602.006.htm |
| [19] | 马木木, 李熙萌, 石莎, 冯金朝. 科尔沁沙地黄柳(Salix gordejevii)光合特性. 中国沙漠, 2015, 35(2): 352–357. |
| [20] | 马少薇, 董建芳, 刘果厚. 内蒙古地区黄柳遗传多样性的RAPD分析. 干旱区资源与环境, 2017, 31(8): 175–180. |
| [21] | 崔秀萍, 刘果厚. 浑善达克沙地黄柳更新特点与规律研究. 中国沙漠, 2012, 32(1): 60–64. |
| [22] | 刘冠志.浑善达克沙地黄柳异速生长与生态适应性研究[D].呼和浩特: 内蒙古农业大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10129-1017212370.htm |
| [23] | Ripley B D. Modelling spatial patterns. Series B (Methodological), 1977, 39(2): 172–212. |
| [24] | Besag J. Contribution to the discussion of Dr. Ripley's paper. Journal of the Royal Statistical Society. Series B (Methodological), 1977, 39(2): 193–195. |
| [25] | 张璞进, 清华, 张雷, 徐延达, 木兰, 晔薷罕, 邱晓, 常虹, 沈海花, 杨劼. 内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局. 植物生态学报, 2017, 41(2): 165–174. |
| [26] | Lotwick H W, Silverman B W. Methods for analysing spatial processes of several types of points. Journal of the Royal Statistical Society. Series B (Methodological), 1982, 44(3): 406–413. |
| [27] | Diggle P J. Statistical Analysis of Spatial Point Patterns. New York: Academic Press, 1983. |
| [28] | Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209–229. DOI:10.1111/oik.2004.104.issue-2 |
| [29] | 王磊, 孙启武, 郝朝运, 田胜尼, 张姗姗, 陈一锟, 张小平. 皖南山区南方红豆杉种群不同龄级立木的点格局分析. 应用生态学报, 2010, 21(2): 272–278. |
| [30] | 张金屯. 数量生态学. 北京: 科学出版社, 2011: 277–279. |
| [31] | 李俊清. 森林生态学. 北京: 高等教育出版社, 2006: 190–195. |
| [32] | Ueno N, Suyama Y, Seiwa K. What makes the sex ratio female-biased in the dioecious tree Salix sachalinensis?. Journal of Ecology, 2007, 95(5): 951–959. DOI:10.1111/jec.2007.95.issue-5 |
| [33] | Myers-Smith I H, Hik D S. Uniform female-biased sex ratios in alpine willows. American Journal of Botany, 2012, 99(7): 1243–1248. DOI:10.3732/ajb.1200107 |
| [34] | Zhang S, Chen L H, Duan B L, Korpelainen H, Li C Y. Populus cathayana males exhibit more efficient protective mechanisms than females under drought stress. Forest Ecology and Management, 2012, 275: 68–78. DOI:10.1016/j.foreco.2012.03.014 |
| [35] | 崔秀萍, 刘果厚, 贺晓. 浑善达克沙地黄柳种子发芽和幼苗生长特性的研究. 西北植物学报, 2006, 26(11): 2312–2317. DOI:10.3321/j.issn:1000-4025.2006.11.020 |
| [36] | 潘春芳, 赵秀海, 夏富才, 张春雨, 吴蕾, 周海成, 林凤友. 长白山山杨种群的性比格局及其空间分布. 生态学报, 2011, 31(2): 297–305. |
| [37] | Ueno N, Seiwa K. Gender-specific shoot structure and functions in relation to habitat conditions in a dioecious tree, Salix sachalinensis. Journal of Forest Research, 2003, 8(1): 9–16. DOI:10.1007/s103100300001 |