 2018, Vol. 38
2018, Vol. 38文章信息
- 吴安驰, 邓湘雯, 任小丽, 项文化, 张黎, 葛蓉, 牛忠恩, 何洪林, 何立杰.
- WU Anchi, DENG Xiangwen, REN Xiaoli, XIANG Wenhua, ZHANG Li, GE Rong, NIU Zhongen, HE Honglin, HE Lijie.
- 中国典型森林生态系统乔木层群落物种多样性的空间分布格局及其影响因素
- Biogeographic patterns and influencing factors of the species diversity of tree layer community in typical forest ecosystems in China
- 生态学报. 2018, 38(21): 7727-7738
- Acta Ecologica Sinica. 2018, 38(21): 7727-7738
- http://dx.doi.org/10.5846/stxb201710191882
-
文章历史
- 收稿日期: 2017-10-19
- 网络出版日期: 2018-08-08





2. 南方林业生态应用技术国家工程实验室, 长沙 410004;
3. 中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室, 北京 100101;
4. 中国科学院大学, 北京 100049;
5. 沈阳农业大学, 沈阳 110866
2. National Engineering Laboratory for Applied Technology of Forestry & Ecology in South China, Changsha 410004, China;
3. Key Laboratory of Ecosystem Network Observation and Modeling, Institution of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China;
4. University of Chinese Academy of Sciences, Beijing 100049, China;
5. Shenyang Agricultural University, Shenyang 110866, China
生物多样性是指一个地区内生命形成的丰富程度, 是生物以及与环境有规律地结合所构成稳定的生态综合体以及与此相关的各种生态过程的综合。它包括物种多样性、遗传多样性及生态系统多样性3个层次。其中, 物种多样性是生物多样性的核心[1]。森林是陆地生态系统生物多样性最重要的资源库, 但森林砍伐、森林退化、气候变化等因素正威胁全球大约一半的树种[2]。植物群落物种多样性的空间分布格局是各种生态梯度变化的综合反映, 研究物种多样性沿环境梯度的变化格局以及影响因子是生物多样性研究的重要议题之一[3-4]。
国内外学者围绕森林物种多样性的分布格局进行了广泛、深入的研究[5-9], 全球和区域范围内低海拔地区不同生态系统的研究结果表明, 植物群落物种多样性具有明显的纬度和海拔梯度变化规律[10]。其中, 针对我国山地植物物种多样性垂直分布格局研究结果表明, 在人为干扰较少的山地, 木本植物物种丰富度一般随着海拔的升高而逐渐减少[11-13]; 但王志恒等[14]在高黎贡山的研究发现, 全部种子植物和不同生活型植物的物种丰富度随海拔升高呈现先增后减的趋势, 即在中海拔地段的丰富度最大。在大尺度上, 森林群落物种丰富度一般随纬度增加而减少[15]。冯建孟等[16]对中国种子植物物种多样性大尺度分布格局进行研究, 发现从随纬度增加, 物种密度呈显著递减趋势, 而物种丰富度的递减趋势不够明显; 陈圣宾等[17]对中国种子植物特有属地理分布格局进行了研究, 发现种子植物属丰富度及特有属比例具有很强的空间变异性, 且表现出显著的纬度梯度性。目前, 关于影响物种多样性分布格局的因素争议较大, 在较小的空间尺度上, 决定物种多样性分布格局的主要因素是生物、生态过程(如竞争、共生和迁徙等)、地形和土壤因子等[18-20]。在大尺度上, 很多学者提出了若干理论或假说解释物种多样性的空间格局, 如水热动态假说[21-22]、环境能量假说[23-24]、生理忍耐假说[25-26]、气候稳定性假说[27-28]和生产力假说[29-31]等。虽然以往的研究对各种假说进行了大量的验证, 但不同区域物种多样性的主导因素及各种假说的适用性存在很大差异。因此, 物种多样性分布格局及其影响因素的研究有待深入。
中国疆域辽阔, 地形复杂, 有丰富的木本植物以及多样的环境梯度, 从温暖潮湿的热带到极度干旱的沙漠和寒冷的北方地区[32], 适合开展大尺度物种多样性格局的研究。衡量不同区域物种多样性分布格局影响因素的相对贡献率成为现阶段物种多样性格局研究的重点[33]。同时, 使用不同区域、不同生态系统的实测数据是研究物种多样性地理格局的形成机制以及假说验证重要手段。本文基于国家生态系统观测研究网络(CNERN)的13个典型森林生态系统综合观测场样地乔木层调查数据, 利用物种丰富度指数、多样性指数和均匀度指数3类指标来测度森林群落乔木层的物种多样性, 并结合气候因子和植物特性数据进行统计分析, 旨在揭示:(1)中国典型森林生态系统乔木层的物种多样性随经纬度变化的空间分布格局; (2)确定植物特性和气候因子对乔木层的物种多样性分布格局的相对贡献率, 解释中国典型森林生态系统乔木层的物种多样性格局形成的主导因子。
1 研究区域和研究方法 1.1 研究区概况本研究选择国家生态系统研究网络(http://www.cnern.org.cn)13个典型森林生态系统综合观测场样地, 涵盖热带(尖峰岭和西双版纳)、亚热带(鼎湖山、哀牢山、会同、神农架和贡嘎山)和温带(秦岭、茂县、北京、长白山、帽儿山和大兴安岭)(图 1), 基本覆盖我国从南到北各种气候带和典型森林植被类型。各个综合观测场样地经过长期的封山育林, 人为干扰少, 植被保护良好, 对每个地区植被类型具有典型代表性。

|
| 图 1 中国13处典型森林生态系统综合性样地分布地点及物种多度直方图 Fig. 1 Locations and species abundance histograms of the comprehensive sites of 13 typical forest ecosystem in China 插图中y坐标为物种数量, x坐标以2为底的物种多度进行对数转换, 即物种个体数分别为1, 2, 3—4, 5—8, 9—16等进行分组 |
13个森林生态站的综合观测场样地均按照统一规范和方法设置, 根据物种面积曲线确定最小面积样方的大小, 且位于所在地区内最具有代表性的生态系统的典型地段[34]。对每各站综合观测场样地内乔木树种的所有个体进行每木检尺, 记录乔木层的物种、株数、胸径、树高、生长状况和空间相对位置等指标, 用卷尺记录物种个体空间坐标精确到±15 cm。各个森林生态系统的生物量数据通过相对生长方程计算法(allometric method)计算获得, 即通过胸径、树高与各器官地上、地下生物量构建经验方程估算森林生态系统的生物量。同时, 记录样地的经纬度、海拔和郁闭度等, 样地的基本信息[35]见表 1。由于在较大空间格局上, 乔木层的物种多样性与海拔没有显著的相关性(P>0.05), 本文的研究未涉及海拔对乔木层物种多样性的影响。
| 样地 Sample plot | 经度 Longitude | 纬度 Latitude | 海拔/m Elevation | 植被类型 Vegetation types | 土壤类型 Soil types | 优势种 Dominant species |
| 尖峰岭 | 109.20 | 18.95 | 68 | 热带常绿季雨林 | 砖红壤 | 九节、大叶白颜 |
| 西双版纳 | 101.20 | 21.96 | 730 | 热带常绿阔叶季雨林 | 砖红壤 | 番龙眼、千果榄仁 |
| 鼎湖山 | 112.54 | 23.17 | 260 | 亚热带季风常绿阔叶林 | 赤红壤 | 锥、木荷、厚壳桂 |
| 哀牢山 | 101.02 | 24.53 | 2488 | 亚热带中山湿性常绿阔叶林 | 山地黄棕壤 | 硬壳柯、木果柯、变色锥 |
| 会同 | 109.60 | 26.85 | 350 | 亚热带常绿阔叶林 | 黄壤 | 红拷、青冈、刨花楠 |
| 神农架 | 110.50 | 31.32 | 1750 | 亚热带常绿落叶阔叶混交林 | 黄棕壤 | 米心青冈、多脉青冈、曼青冈 |
| 贡嘎山 | 101.99 | 29.57 | 3160 | 亚热带亚高山暗针叶林 | 棕色针叶林土 | 冷杉、五尖槭、糙皮桦 |
| 茂县 | 103.90 | 31.78 | 1816 | 暖温带亚高山落叶针叶混交林 | 褐土 | 华山松、油松、锐齿槲栎 |
| 秦岭 | 108.48 | 33.45 | 1585 | 暖温带落叶阔叶混交林 | 棕壤 | 油松、华山松、锐齿砾 |
| 北京 | 115.43 | 39.96 | 1263 | 暖温带落叶阔叶混交林 | 褐土 | 辽东栎、油松、白桦 |
| 长白山 | 128.10 | 42.40 | 784 | 中温带落叶针阔混交林 | 棕色针叶林土 | 红松、五角枫、假色槭 |
| 帽儿山 | 127.52 | 45.35 | 428 | 中温带落叶针阔混交林 | 暗棕壤 | 红松、白桦、水曲柳 |
| 大兴安岭 | 121.51 | 50.96 | 832 | 寒温带针叶林 | 暗棕针叶林土 | 兴安落叶松、白桦 |
| 九节Psychotria rubra; 大叶白颜Gironniera subaequalis; 番龙眼Pometia pinnata; 千果榄仁Terminalia myriocarpa; 锥Castanopsis chinensis Hance; 木荷Schima superba; 厚壳桂Cryptocarya chinensis; 硬壳柯Lithocarpus hancei; 木果柯Lithocarpus xylocarpus; 变色锥Castanopsis rufescens; 红拷Castanopsis hystrix; 青冈Cyclobalanopsis glauca; 刨花楠Machilus pauhoi ; 米心青冈Fagus engleriana; 多脉青冈Cyclobalanopsis multinervis; 曼青冈Cyclobalanopsis oxyodon; 冷杉Abies fabri; 五尖槭Acer maximowiczii Pax; 糙皮桦Betula utilis; 华山松Pinus armandii; 油松Pinus tabuliformis; 锐齿槲栎Quercus aliena; 锐齿砾Quercus aliena var. acuteserrata; 辽东栎Quercus wutaishanica; 白桦Betula platyphylla; 红松Pinus koraiensis; 色木槭Acer mono; 假色槭Acerpseudo-sieboldianum; 水曲柳Fraxinus mandschurica; 兴安落叶松Larix gmelinii | ||||||
根据中国典型森林生态系统的综合观测场样地乔木层调查数据, 本研究选用物种丰富度(S)、Shannon-Wiener指数(H′)、Simpson指数(D)和Pielou指数(E)4个常用指数测度群落乔木层的物种多样性[36], 具体计算公式如下:
物种丰富度(S):
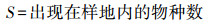
|
(1) |
Simpson指数(D):
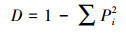
|
(2) |
Shannon-Wiener指数(H′):
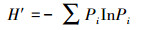
|
(3) |
Pielou指数(E):
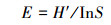
|
(4) |
式中, S为样方中物种数, Pi为群落中第i个物种的重要值。物种重要值(important value, IV)用来反映物种在群落中的相对重要性, 计算公式为:
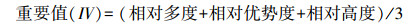
|
(5) |
根据各森林生态站实际观测的月均温和月降水数据(2000—2010年平均值), 计算年均温(annual mean temperature, MAT)、最暖月均温(mean temperature of the warmest month, MTWM)、最冷月均温(mean temperature of the coldest month, MTCM)、温度年较差(annual variation of mean annual temperature, AVMAT)、年降水量(annual mean precipitation, MAP)、最干燥季降水量(precipitation of driest quarter, PDQ)、最湿润季降水量(precipitation of wettest quarter, PWQ)、实际蒸散量(actual evapotranspiration, AET)和潜在蒸散量(potential evapotranspiration, PET), 其中潜在蒸散量和实际蒸散量根据魏焕奇等[37]方法计算。这些指标不仅与物种多样性有着密切关系, 也是验证各假说的关键指标[38]。
归一化植被指数(normalized difference vegetation index, NDVI)可以很好地反映植被生长状况, 与植被覆盖度和生产力显著相关[39]。本研究使用的NDVI数据来自美国国家航空航天局NASA网站(http://glovis.usgs.gov)下载的MOD13Q1数据产品, 其空间分辨率为250 m, 根据综合观测场样地的地理坐标在Arcgis 10.0软件中提取2000—2010年均值用于数据分析。
1.5 数据处理与分析运用一元回归模型检验物种多样性指数与经纬度的关系, 并对物种多样性指数与各自变量进行单因素方差分析(one-way ANOVA), 分别比较物种多样性指数和各自变量之间Pearson相关性系数, 所用的统计显著水平值设为P < 0.05。
将所有因子分为植物特性、能量和水分三大类。其中, 植物特性包括NDVI指数、郁闭度和生物量; 能量因子包括年均温、最暖月均温、最冷月均温、温度年较差、实际蒸散量和潜在蒸散量; 水分因子包括年降水量、最干燥季降水量和最湿润季降水量。对三类因子分别进行主成分分析(principal component analysis, PCA), 提取主成分作为各类因子的替代因子。
数据处理和分析采用Excel 2010、SPSS 18.0和R 3.3.3 (R-Development Core Team 2017)软件完成。物种丰富度在分析之前经过自然对数转换。方差分解(variation partitioning)分析采用R语言的“Vegan”数据包中“Varpart”功能完成。运用韦恩图显示各类因子的纯效应(pure effect)和共同效应(share effect)[40-41]。
物种的个体数转换成以2为底的对数, 并进行合并, 即按照物种的个体数分别为1, 2, 3—4, 5—8, 9—16等进行分组, 按照这种方法作物种多度直方图[42]。表明中国典型森林生态系统综合观测场样地乔木层的常见物种和稀有种差异(图 1)。统计分析在R软件包“sads”中完成。
2 结果与分析 2.1 物种多样性指数随经度和纬度的分布格局中国典型森林生态系统的乔木层物种多样性指数存在较大的空间变异(图 2)。其中, 物种丰富度与纬度(R2=0.765, P < 0.001)和经度(R2=0.410, P < 0.05)均呈显著负相关, 即随经纬度的升高物种丰富度呈逐渐下降趋势。而Shannon-Wiener指数(R2 = 0.694, P < 0.001)、Simpson指数(R2=0.592, P < 0.01)和Pielou指数(R2=0.315, P < 0.05)仅与纬度呈显著负相关, 与经度无显著的负相关关系(P>0.05)。物种丰富度、Shannon-Wiener指数和Simpson指数最高值位于热带的西双版纳, 最低值位于寒温带的大兴安岭。Pielou指数波动较大, 例如温带长白山的Pielou指数值(0.86)高于亚热带哀牢山的Pielou指数值(0.81)。虽然Shannon-Wiener指数、Simpson指数和Pielou指数没有物种丰富度随经纬度变化趋势明显, 但结果表现的趋势一致。

|
| 图 2 物种多样性指数随纬度和经度梯度的变化, 物种丰富度数据均经过自然对数转换 Fig. 2 Species diversity indices along the longitude and latitude gradient change. Species richness data were ln-transformed |
物种多样性指数与各因子的单因素相关性分析结果显示(表 2):物种丰富度与年均温、最冷月均温、温度年较差和潜在蒸散量之间的相关性最强(MAT: R2=0.803, P < 0.001; TEMP: R2 =0.827, P < 0.001; AVMAT: R2 =-0.725, P < 0.001; PET: R2 =0.734, P < 0.001), 与实际蒸散量、NDVI指数和郁闭度的相关性次之(AET: R2 =0.607, P < 0.01; NDVI: R2 =0.675, P < 0.01; CB: R2 =0.457, P < 0.01), 与最暖月均温、年降水量、最湿润季降水量和生物量的相关性较低(P < 0.05)。Shannon-Wiener指数与年均温、最冷月均温和潜在蒸散量的相关性最高(MAT: R2=0.767, P < 0.001; TEMP: R2=0.726, P < 0.001; PET: R2=0.718, P < 0.001), 与温度年较差、实际蒸散量、NDVI指数和郁闭度相关性次之(AVMAT: R2=-0.628, P < 0.01; AET: R2 = 0.651, P < 0.01; NDVI: R2=0.597, P < 0.01; CB: R2=0.537, P < 0.01), 与最暖月均温、年降水量、最湿润季降水量和生物量的相关性较弱(P < 0.05)。Simpson指数与年均温、最冷月均温、温度年较差、实际蒸散量和潜在蒸散量最强(MAT: R2=0.615, P < 0.01; TEMP: R2=0.632, P < 0.01; AVMAT: R2=-0.510, P < 0.01; AET: R2=0.617, P < 0.01; PET: R2=0.466, P < 0.01), 与最暖月均温和NDVI指数呈较低显著正相关(P < 0.05)。Pielou指数仅与年均温、最冷月均温、实际蒸散量、潜在蒸散量和郁闭度之间具有显著相关性(P < 0.05)。
| 变量 Variable | 物种丰富度对数值 ln(spices richness) | Shannon-Wiener指数 Shannon-Wiener index | Simpson指数 Simpson index | Pielou指数 Pielou index |
| 年均温MAT/℃ | 0.803*** | 0.767*** | 0.615** | 0.359* |
| 最暖月均温MTWM/℃ | 0.407* | 0.374* | 0.385* | 0.208ns |
| 最冷月均温TEMP/℃ | 0.827*** | 0.726*** | 0.632** | 0.306* |
| 温度年较差AVMAT/℃ | -0.725*** | -0.628** | -0.510** | -0.239ns |
| 实际蒸散量AET/mm | 0.607** | 0.651** | 0.466** | 0.356* |
| 潜在蒸散量PET/mm | 0.734*** | 0.718*** | 0.617** | 0.355* |
| 年降水量MAP/mm | 0.345* | 0.386* | 0.249ns | 0.191ns |
| 最干燥季降水量PDQ/mm | 0.200ns | 0.245ns | 0.219ns | 0.174ns |
| 最湿润季降水量PWQ/mm | 0.324* | 0.382* | 0.184ns | 0.166ns |
| 植被归一化指数NDVI | 0.675** | 0.597** | 0.312* | 0.171ns |
| 郁闭度CB | 0.457** | 0.537** | 0.267ns | 0.312* |
| 生物量B | 0.306* | 0.329* | 0.275ns | 0.184ns |
| * P < 0.05; ** P < 0.01; *** P < 0.001; ns, P>0.05. MAT:年均温annual mean temperature; MTWM:最暖月均温mean temperature of the warmest month; MTCM:最冷月均温mean temperature of the coldest month; AVMAT:温度年较差annual variation of mean annual temperature; AET:实际蒸散量actual evapotranspiration; PET:潜在蒸散量potential evapotranspiration; MAP:年降水量annual mean precipitation; PDQ:最干燥季降水量precipitation of driest quarter; PWQ:最湿润季降水量precipitation of wettest quarter; DNVI:植被归一化指数normalized difference vegetation index; CB:郁闭度crown density; B:生物量biomass | ||||
物种多样性指数与植物特性或气候因子的单因素相关性相差较大。其中, 物种丰富度和Shannon-Wiener指数与多个因子具有显著相关性; Simpson指数与能量因子的相关性较高, 与水分因子不具有显著相关性(P>0.05);Pielou指数与各因子之间的相关性相对较低。温度年较差与物种丰富度、Shannon-Wiener指数和Simpson指数均呈强负相关(P < 0.01), 但与Pielou指数呈不显著负相关(R2=-0.239, P>0.05)。其中最干燥季降水量与物种多样性指数均没有显著相关性(P> 0.05)。
2.3 植物特性和气候因子对物种多样性分布格局的影响主成分分析表明, 能量因子第一个主成分的特征值为5.101, 贡献率为85.020%, 以最冷月均温(0.990)的载荷量最大; 水分因子第一主成分的特征值为2.443, 贡献率为81.432%, 以年降水量的载荷量最大, 为0.990;植物特性第一主成分的特征值为2.157, 贡献率为71.889%, 生物量的载荷量最大, 为0.881(表 3), 其成分矩阵见表 4。三类因子的第一主成分指标载荷量较大(载荷量绝对值均大于0.7)(表 4), 所以分别选择三类因子第一主成分代表各类因子主要信息的综合反映。
| 类型 Class | 成分 Component | 特征值 Eigenvalue | 方差贡献率 Contribution rate of variance/% | 方差累计贡献率 Cumulative contribution rate of variance/% |
| 能量因子 | 1 | 5.101 | 85.020 | 85.020 |
| Energy factors | 2 | 0.707 | 11.780 | 96.801 |
| 3 | 0.145 | 2.411 | 99.211 | |
| 4 | 0.029 | 0.487 | 99.699 | |
| 5 | 0.018 | 0.301 | 99.999 | |
| 6 | 0.001 | 0.001 | 100.000 | |
| 水分因子 | 1 | 2.443 | 81.432 | 81.432 |
| Moisture factors | 2 | 0.535 | 17.836 | 99.268 |
| 3 | 0.022 | 0.732 | 100.000 | |
| 植物特性 | 1 | 2.157 | 71.889 | 71.889 |
| Plant traits | 2 | 0.508 | 16.936 | 88.825 |
| 3 | 0.355 | 11.175 | 100.000 |
| 能量因子 Energy factors | 成分Component | |||||
| PCA1 | PCA2 | PCA3 | PCA4 | PCA5 | PCA6 | |
| 年均温MAT/℃ | 0.986 | 0.104 | -0.045 | 0.011 | 0.118 | 0.001 |
| 最暖月均温MTWM/℃ | 0.751 | 0.649 | -0.119 | 0.008 | -0.042 | -0.002 |
| 最冷月均温TEMP/℃ | 0.990 | -0.067 | -0.111 | -0.054 | -0.031 | 0.005 |
| 温度年较差AVMAT/℃ | -0.897 | 0.429 | 0.080 | 0.074 | 0.016 | 0.004 |
| 实际蒸散量AET/mm | 0.935 | 0.124 | 0.332 | -0.033 | -0.013 | 0.002 |
| 潜在蒸散量PET/mm | 0.953 | -0.268 | 0.005 | 0.140 | -0.03 | 0.001 |
| 水分因子 Moisture factors | 成分Component | |||||
| PCA1 | PCA2 | PCA3 | ||||
| 年降水量MAP/mm | 0.990 | -0.086 | -0.113 | |||
| 最干燥季降水量PDQ/mm | 0.811 | 0.584 | 0.045 | |||
| 最湿润季降水量PWQ/mm | 0.898 | -0.432 | 0.084 | |||
| 植物特性 Plant traits | 成分Component | |||||
| PCA1 | PCA2 | PCA3 | ||||
| 植被归一化指数NDVI | 0.807 | 0.582 | 0.102 | |||
| 郁闭度CB | 0.855 | -0.374 | 0.360 | |||
| 生物量B | 0.881 | -0.170 | -0.442 | |||
| 缩写代表含义同表 2 | ||||||
进一步通过方差分解分析, 结果表明能量因子和水分因子的共同作用对物种丰富度、Shannon-Wiener指数、Simpson指数和Pielou指数的解释率最大, 分别为42%、38%、26%和15%;植物特性、能量因子和水分因子三者共同作用对物种丰富度、Shannon-Wiener指数、Simpson指数和Pielou指数的解释率次之, 分别为25%、27%、24%和14%。能量因子对物种丰富度、Shannon-Wiener指数和Simpson指数单独解释率分别为17%、13%和17%, 对Pielou指数没有产生任何影响。植物特性与能量因子或者水分因子两者之间的共同作用以及植物特性和水分因子独立作用对物种多样性指数空间分布格局的解释率较小, 甚至无影响(图 3)。

|
| 图 3 方差分解分析植物特性、能量和水分因子对物种多样性指数的纯效应和共同效应 Fig. 3 Individual and shared effects of plant traits, energy and moisture factors to species diversity indices as derived from variation partitioning analysis |
在中国典型森林生态系统中乔木层物种丰富度与纬度相关性最强, 即随纬度升高物种丰富度呈显著下降, 与已有学者[16, 43]对我国木本植物和种子植物的物种丰富度与纬度分布规律的研究结果一致。其中, 乔木层的Shannon-Wiener指数、Simpson指数和Peilou指数空间分布格局随纬度呈显著负相关, 与物种丰富度的变化规律一致。在热带的西双版纳和尖峰岭地区, 稀有物种和中间物种个体数量较多, 物种多样性指数整体较高。亚热带森林受复杂地形因子和水热条件变化的影响, 森林结构比较复杂。鼎湖山、哀牢山和神农架的森林群落郁闭度约在85%—92%, 林龄超过200年, 都是典型的地带性顶极森林群落, 物种多样性指数仅次于热带森林群落。从暖温带到寒温带地区, 稀有物种逐渐减少, 常见物比例增大, 群落结构变得简单, 乔木层物种多样性逐渐减少。Peilou指数随纬度变化趋势与中国东部32个森林群落Pielou均匀度指数的研究结果一致[44]。在长白山群落中优势物种比例较大, 稀有物种较少, Peilou指数最高; 在海拔较高的哀牢山Peilou指数较高, 2—4个偶见种和常见物种个体数较多, 稀有物种个体数较少, 哀牢山乔木层的群落物种分布比较均匀。
乔木层的物种多样性指数在经度梯度上波动较大, 除了物种丰富度与经度呈显著负相关(P < 0.05)以外, Shannon-Wiener指数、Simpson指数和Pielou指数在经度上均呈不显著负相关关系。在经度上, 乔木层物种多样性指数在西双版纳和109—113°E之间明显较高, 较低值位于喜马拉雅-横断山区和北方地区。结果表明在中国典型森林生态系统中乔木层物种多样性南北方向的联系强度大于东西方向的联系, 与中国高山植物区系的地理空间格局研究一致[45]。
3.2 气候因子对物种多样性格局的影响水热动态假说认为物种多样性的大尺度格局由水分和能量共同决定。这一假说通常用潜在蒸散量和温度表示热能, 而水分一般指液态水, 通常用一个地区的年降水量表示, 也有用实际蒸散量反映一个地区的水热动态平衡[21-22]。能量和水分的空间分异越大, 意味着在能量和水分梯度上存在越多样的生境满足来不同物种水热的需要, 为植物造就更加多样的生境, 从而支持更多的物种得以共存和分布, 故物种多样性较高。方差分解结果表明, 能量和水分因子的共同作用对物种丰富度、Shannon-Wiener指数、Simpson指数和Pielou指数的解释率分别达到42%、38%、26%和15%, 这在一定程度上解释能量和水分因子共同作用对物种多样性指数的空间分布格局起决定作用, 进而影响物种多样性分布格局, 支持了水热动态假说。
能量因子对乔木层的物种丰富度、Shannon-Wiener指数和Simpson指数单独解释率分别为17%、13%和17%, 在单因素相关分析中年均温、最冷月均温和潜在蒸散量差异均显著影响物种多样性指数的分布格局(P < 0.05), 而且相对于其他因子的空间分异而言, 其相关性更高。所以环境能量假说适合解释中国典型森林生态系统乔木层物种多样性的分布格局。寒冷忍耐假说认为, 在寒冷地区, 很多物种由于不能忍受冬季的寒冷而无法生存。因此, 随着冬季温度的降低, 其物种多样性逐渐减少[38, 46]。本研究结果表明最冷月均温与物种多样性指数均呈强正相关, 说明冬季低温对物种多样性的分布格局构成了很强的限制作用, 比如在大兴安岭最低温度为-29℃, 主要乔木树种只有兴安落叶松(Larix gmelinii)和白桦(Betula platyphylla); 由于绝对大部分物种的祖先都是在湿热的环境中进化, 极端环境条件导致植物个体水平的死亡, 通过影响物种的种群动态, 最终影响到物种的分布范围和物种多样性的分布格局[47-48]。本研究结果同样支持寒冷忍耐假说。
根据气候稳定性假说, 在气候稳定地区物种的生态位相对较窄, 所以就有更多的空间容纳更多的物种; 反之, 在气候波动较少的地区, 可能资源的供应相对而言较不稳定, 物种的生态位相对较宽, 能容纳的物种相对较少, 所以稳定的自然资源更有利于物种的进化与适应[27-28]。这一假说通常用温度年较差来表示, 说明该地区温度的季节波动性, 用于表征气候的稳定性。物种丰富度、Shannon-Wiener指数和Simpson指数均与温度年较差有很强的负相关(P < 0.01), Pielou指数与温度年较差呈不显著负相关。这说明温度年较差在很大程度上决定了物种多样性的空间分布格局。随着纬度的增加, 温度年较差呈递减趋势, 这也解释了本研究中乔木层的物种多样性由南向北的递减趋势。
3.3 植物特性对物种多样性格局的影响生产力假说常采用归一化植被指数、实际蒸散量和年降水量作为生产力的替代指标[29-31]。单因素相关分析表明, 物种丰富度与NDVI指数、实际蒸散量和年降水量均有显著正相关(P < 0.05), 其中物种丰富度与NDVI指数的相关性最显著(R2=0.675, P < 0.01), 表明NDVI指数与实际蒸散量或年降水量相比能更好的解释物种丰富度的大尺度格局, 支持物种丰富度格局的生产力假说。NDVI指数与Shannon-Wiener指数和Simpson指数也具有显著相关性。由于Pielou指数反映群落内的个体密度和均匀情况, NDVI指数反映植被覆盖程度, Pielou指数和NDVI指数呈不显著相关性。此外, 郁闭度通过改变林内光照条件、植物物种组成和群落结构, 从而形成森林群落物种共存以及维持多样性的重要场所, 但郁闭度和物种多样性不存在线性关系[49]。南方森林群落结构复杂, 森林群落结构动态变化大, 而北方乔木层居优势地位, 群落结构稳定, 郁闭度相对南方较低。在13个典型森林乔木层的群落郁闭度与物种丰富度、Shannon-Wiener指数和Pielou指数均呈显著正相关, 与Simpson指数呈不显著的正相关。生物量与物种丰富度和Shannon-Wiener指数均呈显著正相关(P < 0.05), 生物量高的植物群落其物种多样性也相对较高, 这说明群落中的物种多样性和生物量是相互影响。在一定程度上促进森林群落生物量的发展, 同时也有利于提高群落物种多样性增加, 植物多样性的保护应优先保护植物生物量较高的区域。进一步通过方差分解发现, 植物特性、能量因子和水分因子三者共同作用对乔木层的物种多样性指数能解释到14%—27%的变异, 植物特性对乔木层的物种丰富度和Shannon-Wiener指数单独解释率分别为7%和4%。所以, 在解释物种多样性大尺度分布格局时, 也必须充分考虑植物因素的影响。
综上所述, 本研究主要考虑植物特性和气候因子的影响, 仍有一些影响物种多样性空间分布格局的因素未被涉及, 如微环境(坡度、坡向、海拔等)、土壤因子(有机碳、全氮、全磷等)、边缘效应、区域历史、生境异质性、人为干扰以及种子传播等。未来应该综合考虑各种生态因子, 从而更加全面的解释中国典型森林生态系统物种多样性空间分布格局及其影响因素。
4 结论基于中国典型森林生态系统乔木层群落调查数据, 结合植物特性和气候数据, 研究大尺度物种多样性的分布格局及其影响因素。主要结论如下:
(1) 乔木层的物种多样性指数在地理格局上存在较大差异, 均随经纬度上升而下降。其中, 物种多样性指数在纬度梯度上均呈显著相关性, 在经度上波动较大, 表明乔木层物种多样性在纬度梯度上变化规律更为明显。
(2) 物种多样性指数与各因子的单因素相关性相差较大。物种丰富度、Shannon-Wiener指数和Simpson指数与能量因子相关性较强, Pielou指数仅与年均温、最冷月均温、实际蒸散量、潜在蒸散量和郁闭度存在显著相关性。其中, Simpson指数、Pielou指数与水分因子无显著相关性, 物种丰富度和Shannon-Wiener指数均与DNVI指数、郁闭度和生物量具有显著的相关性。
(3) 植物特性和气候因子对乔木层的物种多样性指数分布格局的解释率存在较大差异。能量和水分因子的共同作用对乔木层物种多样性指数的解释率最高, 植物特性、能量和水分因子三者的共同作用解释率次之。其中, 能量因子单独解释率高于植物特性或水分因子, 植物特性与能量因子或水分因子之间的共同作用以及植物特性和水分因子单独作用对乔木层的物种多样性指数解释率较小, 甚至无影响。
致谢: 感谢国家生态系统观测研究网络(CNERN)、中国生态系统研究网络(CERN)以及野外观测站的支持。| [1] | 魏辅文, 聂永刚, 苗海霞, 路浩, 胡义波. 生物多样性丧失机制研究进展. 科学通报, 2014, 59(6): 430–437. |
| [2] | Liang J J, Crowther T W, Picard N, Wiser S, Zhou M, Alberti G, Schulze E D, McGuire A D, Bozzato F, Pretzsch H, de-Miguel S, Paquette A, Hérault B, Scherer-Lorenzen M, Barrett C B, Glick H B, Hengeveld G M, Nabuurs G J, Pfautsch S, Viana H, Vibrans A C, Ammer C, Schall P, Verbyla D, Tchebakova N, Fischer M, Watson J V, Chen H Y H, Lei X D, Schelhaas M J, Lu H C, Gianelle D, Parfenova E I, Salas C, Lee B, Kim H S, Bruelheide H, Coomes D A, Piotto D, Sunderland T, Schmid B, Gourlet-Fleury S, Sonké B, Tavani R, Zhu J, Brandl S, Vayreda J, Kitahara F, Searle E B, Neldner V J, Ngugi M R, Baraloto C, Frizzera L, Bałazy R, Oleksyn J, Zawiła-Niedźwiecki T, Bouriaud O, Bussotti F, Finér L, Jaroszewicz B, Jucker T, Valladares F, Jagodzinski A M, Peri P L, Gonmadje C, Marthy W, O'Brien T, Martin E H, Marshall A R, Rovero F, Bitariho R, Niklaus P A, Alvarez-Loayza P, Chamuya N, Valencia R, Mortier F, Wortel V, Engone-Obiang N L, Ferreira L V, Odeke D E, Vasquez R M, Lewis S L, Reich P B. Positive biodiversity-productivity relationship predominant in global forests. Science, 2016, 354(6309): aaf8957. DOI:10.1126/science.aaf8957 |
| [3] | Kratochwil A. Biodiversity in ecosystems: some principles//Kratochwil A, ed. Biodiversity in Ecosystems: Principles and Case Studies of Different Complexity Levels. Dordrecht: Springer, 1999: 5-38. |
| [4] | 赵常明, 陈伟烈, 黄汉东, 田自强, 陈玥, 谢宗强. 三峡库区移民区和淹没区植物群落物种多样性的空间分布格局. 生物多样性, 2007, 15(5): 510–522. DOI:10.3321/j.issn:1005-0094.2007.05.009 |
| [5] | 高贤明, 马克平, 陈灵芝. 暖温带若干落叶阔叶林群落物种多样性及其与群落动态的关系. 植物生态学报, 2001, 25(3): 283–290. DOI:10.3321/j.issn:1005-264X.2001.03.005 |
| [6] | Qian H, Ricklefs R E. A latitudinal gradient in large-scale beta diversity for vascular plants in North America. Ecology Letters, 2007, 10(8): 737–744. DOI:10.1111/ele.2007.10.issue-8 |
| [7] | Kraft N J B, Comita L S, Chase J M, Sanders N J, Swenson N G, Crist T O, Stegen J C, Vellend M, Boyle B, Anderson M J, Cornell H V, Davies K F, Freestone A L, Inouye B D, Harrison S P, Myers J A. Disentangling the drivers of β diversity along latitudinal and elevational gradients. Science, 2011, 333(6050): 1755–1758. DOI:10.1126/science.1208584 |
| [8] | Qian H, Jin Y, Ricklefs R E. Phylogenetic diversity anomaly in angiosperms between eastern Asia and eastern North America. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(43): 11452. DOI:10.1073/pnas.1703985114 |
| [9] | Wang Q G, Su X Y, Shrestha N, Liu Y P, Wang S Y, Xu X T, Wang Z H. Historical factors shaped species diversity and composition of Salix in eastern Asia. Scientific Reports, 2017, 7: 42038. DOI:10.1038/srep42038 |
| [10] | Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220–227. DOI:10.1038/35012228 |
| [11] | 唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释. 生物多样性, 2004, 12(1): 115–122. DOI:10.3321/j.issn:1005-0094.2004.01.014 |
| [12] | 刘增力, 郑成洋, 方精云. 河北小五台山北坡植物物种多样性的垂直梯度变化. 生物多样性, 2004, 12(1): 137–145. DOI:10.3321/j.issn:1005-0094.2004.01.017 |
| [13] | 郑成洋, 刘增力, 方精云. 福建黄岗山东南坡和西北坡乔木物种多样性及群落特征的垂直变化. 生物多样性, 2004, 12(1): 63–74. DOI:10.3321/j.issn:1005-0094.2004.01.008 |
| [14] | 王志恒, 陈安平, 朴世龙, 方精云. 高黎贡山种子植物物种丰富度沿海拔梯度的变化. 生物多样性, 2004, 12(1): 82–88. DOI:10.3321/j.issn:1005-0094.2004.01.010 |
| [15] | 陈圣宾, 欧阳志云, 郑华, 肖燚, 徐卫华. 美洲森林群落beta多样性的纬度梯度性. 生态学报, 2011, 31(5): 1334–1340. |
| [16] | 冯建孟. 中国种子植物物种多样性的大尺度分布格局及其气候解释. 生物多样性, 2008, 16(5): 470–476. DOI:10.3321/j.issn:1005-0094.2008.05.007 |
| [17] | 陈圣宾, 欧阳志云, 方瑜, 李振基. 中国种子植物特有属的地理分布格局. 生物多样性, 2011, 19(4): 414–423. |
| [18] | Whittaker R J, Willis K J, Field R. Scale and species richness:towards a general, hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4): 453–470. DOI:10.1046/j.1365-2699.2001.00563.x |
| [19] | 张学雷, 陈杰, 张甘霖. 海南岛不同地形上土壤性质的多样性分析. 地理学报, 2003, 58(6): 839–844. DOI:10.3321/j.issn:0375-5444.2003.06.006 |
| [20] | 丁佳, 吴茜, 闫慧, 张守仁. 地形和土壤特性对亚热带常绿阔叶林内植物功能性状的影响. 生物多样性, 2011, 19(2): 158–167. |
| [21] | O'Brien E. Water-energy dynamics, climate, and prediction of woody plant species richness:an interim general model. Journal of Biogeography, 1998, 25(2): 379–398. DOI:10.1046/j.1365-2699.1998.252166.x |
| [22] | Hawkins B A, Field R, Cornell H V, Currie D J, Guégan J F, Kaufman D M, Kerr J T, Mittelbach G G, Oberdorff T, O'Brien E M, Porter E E, Turner J R G. Energy, water, and broad-scale geographic patterns of species richness. Ecology, 2003, 84(12): 3105–3117. DOI:10.1890/03-8006 |
| [23] | Pianka E R. Latitudinal gradients in species diversity:a review of concepts. The American Naturalist, 1966, 100(910): 33–46. DOI:10.1086/282398 |
| [24] | O'Brien E M, Field R, Whittaker R J. Climatic gradients in woody plant (tree and shrub) diversity:water-energy dynamics, residual variation, and topography. Oikos, 2000, 89(3): 588–600. DOI:10.1034/j.1600-0706.2000.890319.x |
| [25] | Sakai A, Weiser C J. Freezing resistance of trees in north America with reference to tree regions. Ecology, 1973, 54(1): 118–126. DOI:10.2307/1934380 |
| [26] | Currie D J, Francis A P. Regional versus climatic effect on taxon richness in angiosperms:reply to Qian and Ricklefs. The American Naturalist, 2004, 163(5): 780–785. DOI:10.1086/383596 |
| [27] | Klopfer P H. Environmental determinants of faunal diversity. The American Naturalist, 1959, 93(873): 337–342. DOI:10.1086/282092 |
| [28] | Klopfer P H, MacArthur R H. Niche size and faunal diversity. The American Naturalist, 1960, 94(877): 293–300. DOI:10.1086/282130 |
| [29] | Currie D J, Paquin V. Large-scale biogeographical patterns of species richness of trees. Nature, 1987, 329(6137): 326–327. DOI:10.1038/329326a0 |
| [30] | Abramsky Z, Rosenzweig M L. Tilman's predicted productivity-diversity relationship shown by desert rodents. Nature, 1984, 309(5964): 150–151. DOI:10.1038/309150a0 |
| [31] | Hurlbert A H, Haskell J P. The effect of energy and seasonality on avian species richness and community composition. The American Naturalist, 2003, 161(1): 83–97. DOI:10.1086/345459 |
| [32] | Wang Z H, Fang J Y, Tang Z Y, Shi L. Geographical patterns in the beta diversity of China's woody plants:the influence of space, environment and range size. Ecography, 2012, 35(12): 1092–1102. DOI:10.1111/j.1600-0587.2012.06988.x |
| [33] | Wang Z H, Fang J Y, Tang Z Y, Lin X. Relative role of contemporary environment versus history in shaping diversity patterns of China's woody plants. Ecography, 2012, 35(12): 1124–1133. DOI:10.1111/j.1600-0587.2011.06781.x |
| [34] | 中国生态系统研究网络科学委员会. 陆地生态系统生物观测规范. 北京: 中国环境科学出版社, 2007: 13–85. |
| [35] | 邓小保, 唐建维. 中国生态系统定位观测与研究数据集:森林生态系统卷.云南西双版纳站(1998-2006). 北京: 中国农业出版社, 2010. |
| [36] | 方精云, 沈泽昊, 唐志尧, 王志恒. "中国山地植物物种多样性调查计划"及若干技术规范. 生物多样性, 2004, 12(1): 5–9. DOI:10.3321/j.issn:1005-0094.2004.01.002 |
| [37] | 魏焕奇, 何洪林, 刘敏, 张黎, 于贵瑞, 闵程程, 王辉民, 刘影. 基于遥感的千烟洲人工林蒸散及其组分模拟研究. 自然资源学报, 2012, 27(5): 778–789. |
| [38] | 王志恒, 唐志尧, 方精云. 物种多样性地理格局的能量假说. 生物多样性, 2009, 17(6): 613–624. |
| [39] | Defries R S, Townshend J R G. NDVI-derived land cover classifications at a global scale. International Journal of Remote Sensing, 1994, 15(17): 3567–3586. DOI:10.1080/01431169408954345 |
| [40] | Borcard D, Legendre P, Drapeau P. Partialling out the spatial component of ecological variation. Ecology, 1992, 73(3): 1045–1055. DOI:10.2307/1940179 |
| [41] | Peres-Neto P R, Legendre P, Dray S, Borcard D. Variation partitioning of species data matrices:estimation and comparison of fractions. Ecology, 2006, 87(10): 2614–2625. DOI:10.1890/0012-9658(2006)87[2614:VPOSDM]2.0.CO;2 |
| [42] | Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton: Princeton University Press, 2001: 375–375. |
| [43] | 方精云. 探索中国山地植物多样性的分布规律. 生物多样性, 2004, 12(1): 1–4. DOI:10.3321/j.issn:1005-0094.2004.01.001 |
| [44] | Qiao X J, Tang Z Y, Shen Z H, Fang J Y. What causes geographical variation in the species-area relationships? A test from forests in China. Ecography, 2012, 35(12): 1110–1116. DOI:10.1111/j.1600-0587.2011.06869.x |
| [45] | 沈泽昊, 杨明正, 冯建孟, 李新辉, 彭培好, 郑智. 中国高山植物区系地理格局与环境和空间因素的关系. 生物多样性, 2017, 25(2): 182–194. |
| [46] | Hawkins B A, Porter E E, Diniz-Filho J A F. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology, 2003, 84(6): 1608–1623. DOI:10.1890/0012-9658(2003)084[1608:PAHAPO]2.0.CO;2 |
| [47] | Ricklefs R E. History and diversity:explorations at the intersection of ecology and evolution. The American Naturalist, 2007, 170(S2): S56–S70. DOI:10.1086/519402 |
| [48] | Hawkins B A, Devries P J. Tropical niche conservatism and the species richness gradient of North American butterflies. Journal of Biogeography, 2009, 36(9): 1698–1711. DOI:10.1111/jbi.2009.36.issue-9 |
| [49] | Jules M J, Sawyer J O, Jules E S. Assessing the relationships between stand development and understory vegetation using a 420-year chronosequence. Forest Ecology and Management, 2008, 255(7): 2384–2393. DOI:10.1016/j.foreco.2007.12.042 |