 2018, Vol. 38
2018, Vol. 38文章信息
- 曾立雄, 雷蕾, 王晓荣, 朱建华, 肖文发, 刘贤德, 敬文茂.
- ZENG Lixiong, LEI Lei, WANG Xiaorong, ZHU Jianhua, XIAO Wenfa, LIU Xiande, JING Wenmao.
- 海拔梯度对祁连山青海云杉林乔木层和土壤层碳密度的影响
- Effect of altitudinal variation on carbon density in arbor layer and soil layer of Picea crassifolia forest in Qilian Mountains
- 生态学报. 2018, 38(20): 7168-7177
- Acta Ecologica Sinica. 2018, 38(20): 7168-7177
- http://dx.doi.org/10.5846/stxb201711302152
-
文章历史
- 收稿日期: 2017-11-30
- 网络出版日期: 2018-07-09






2. 南京林业大学南方现代林业协同创新中心, 南京 210037;
3. 湖北省林业科学研究院, 武汉 430075;
4. 甘肃省祁连山水源涵养林研究院, 张掖 734000
2. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China;
3. Forestry Institute of Hubei Province, Wuhan 430075, China;
4. Academy of Water Resources Conservation Forests in Qilian Mountain of Gansu Province, Zhangye 734000, China
近几十年来, 由于大气CO2等温室气体浓度迅速上升引起的全球变暖已成为全球性的环境问题[1]。研究发现, 21世纪全球气候变暖趋势将更加严重, 全球表面温度预计将升高1.4—5.8℃[2]。森林是陆地生态系统上最大的碳库, 占据着全球植被碳库的86%和土壤碳库的73%, 在维持全球碳平衡以及应对气候变化中起着关键作用[3], 特别是高海拔高纬度地带植被有机碳对气候变暖响应更敏感[4-5], 碳输入和分解过程平衡的微小变化就能够引起全球CO2浓度的剧烈变化[6]。因此, 了解森林生态系统碳库分布特征以及对气候变化的响应规律, 探索并找到影响碳循环平衡机制, 已经成为国内外学者关注的热点[7-8]。
海拔梯度变化作为一种自然地理变化, 包含多种环境因子变化, 如温度、湿度、光照等环境因子, 这些环境因子的差异对森林土壤理化性质、植被分布以及因植被密度不同导致凋落物量的变化都有较大影响, 而这些因素的变化又会直接或间接的影响森林土壤碳库大小及碳库组成的变化[9]。因此, 利用海拔梯度变化来了解植物适应变化特征以及碳库组成动态变化规律, 对于了解植物响应气候变化和区域碳平衡发展方向具有重要意义[10]。目前国内外许多学者依据海拔梯度变化在我国许多高海拔区域的森林植被开展了土壤有机碳变化相关研究, 如长白山[8]、鼎湖山[9]、喜马拉雅山[11]、川西亚高山[12]、庐山[13]、天山[14]、神农架[15]等区域典型森林植被的土壤有机碳库、碳组分、分布格局以及影响因素等进行了探讨。这些研究多数是以不同海拔不同植被类型来进行研究, 而不同森林植被下土壤由于承接其凋落物和根系分泌物类型不同以及气候因子等差异[12], 使得研究者在分析森林碳库响应海拔梯度变化时会十分复杂。由于高海拔地区单一植被类型具有结构简单、类型单一[14], 在探讨海拔梯度变化上的环境水热配比、植被生长响应、有机碳周转以及预测气候变化方面更准确。但是, 不同区域的海拔高度、植被类型、气候条件等方面的不同导致有机碳变化规律必定存在差异, 目前针对单一植被类型山地生态系统沿海拔梯度分布格局的植被和土壤碳库研究还较少。
祁连山地处中国西北内陆, 位于青藏高原、黄土高原和蒙新荒漠的交汇处, 自然条件复杂、水热条件差异较大, 具有典型的高寒山地特征, 且海拔梯度垂直变化较大, 是研究植被响应气候变化的理想区域[15-16]。青海云杉(Picea crassifolia)为松科云杉属植物, 是中国特有植物, 主要分布在中国大陆的青海、甘肃、内蒙古、宁夏等地。青海云杉林是祁连山区主要森林类型, 面积约占该区乔木林总面积的75.7%, 垂直分布范围较高, 海拔跨度自2500—3300 m[17]。目前此区域针对青海云杉的研究主要包括林分生长与环境因子的关系[16]、生物量分布格局[17]、林分结构[18]、基于GIS技术青海云杉生物量和碳储量估算[19]、林斑表层土壤有机碳特征[20], 仅在一定分布范围内青海云杉林单个碳密度开展相关研究, 并未对基于青海云杉完整的垂直分布界限的乔木层和土壤层有机碳密度协同变化进行研究。
本研究依据祁连山青海云杉沿海拔梯度分布范围, 调查分析了青海云杉乔木层和土壤层有机碳密度的沿海拔梯度上的空间变化特征, 旨在深入认识祁连山青海云杉森林生态系统碳库功能, 为准确估算祁连山青海云杉林碳密度以及基于气候变化下有机碳变化趋势提供科学依据。
1 研究区概况研究区位于祁连山中段甘肃省肃南县西水林区(100°17′E, 38°24′N), 该区属温带高寒半干旱、半湿润山地森林草原气候, 年均气温-0.6—2.0℃, 年均降水量为435.5 mm, 年均蒸发量为1051.7 mm, 年均相对湿度为60%。该流域阳坡为山地干草原, 阴坡为森林景观, 以藓类青海云杉林为主, 海拔3300 m以上分布着亚高山湿性灌木林, 森林总面积168.3 hm2, 覆盖率65%。以青海云杉为主的建群种呈斑块状分布于阴坡、半阴坡, 林分年龄基本相同, 均为中龄林[16], 且零星分布着祁连圆柏(Sabina prezewalskii), 灌木以箭叶锦鸡儿(Caragana jubuta)、高山柳(Salix wihelmsiana)、吉拉柳(Salix gilashanica)和金露梅(Dasiphora fruticosa)为主, 草本主要有珠牙蓼(Polygonum viviparum)、黑穗苔(Carex atrata)和针茅(Shpa)等。土壤类型主要有山地森林灰褐土、山地栗钙土和亚高山灌丛草甸土, 土层较薄, 以粉砂快为主, 成土母质主要是泥炭岩、砾岩、紫色沙页岩等[17, 20]。
2 研究方法 2.1 样地设置及调查2011年, 在祁连山排露沟流域选择青海云杉林作为研究对象, 选择自然生长良好、无火灾影响的区域, 依据青海云杉分布界限, 采用梯度格局法, 在海拔2500—3300 m按每隔100 m的梯度与等高线平行设置样地, 每个海拔梯度上重复3次, 样地面积大小为20 m×20 m, 共计27块。对样方中胸径≥3 cm的所有树种进行每木检尺, 记录其林分起源、物种名称、郁闭度、胸径、树高等指标。记录每个样地的经纬度、海拔、坡度、坡向、土层厚度等环境因子, 样地基本特征见表 1。
| 林分类型 Stand type |
海拔 Altitudinal/m |
样地数量 Number of samples |
坡度 Slope/(°) |
坡向 Slope aspect |
郁闭度 Canopy density |
林分密度 Forest density/(株/hm2) |
平均树高 Mean tree height/m |
平均胸径 Mean diameter at breast height/cm |
平均土壤厚度 Mean soil thickness/cm |
| 青海云杉 | 2500 | 3 | 41 | 东北 | 0.75 | 1142 | 7.89 | 13.66 | 62.0 |
| P. crassifolia | 2600 | 3 | 40 | 东北 | 0.90 | 1375 | 8.58 | 13.05 | 63.6 |
| 2700 | 3 | 29 | 东北 | 0.85 | 1766 | 8.86 | 13.34 | 70.5 | |
| 2800 | 3 | 18 | 东北 | 0.75 | 2058 | 9.25 | 14.65 | 60.6 | |
| 2900 | 3 | 15 | 东北 | 0.60 | 1550 | 7.62 | 12.24 | 60.0 | |
| 3000 | 3 | 16 | 东北 | 0.70 | 1500 | 7.73 | 15.23 | 60.0 | |
| 3100 | 3 | 21 | 东北 | 0.60 | 1025 | 6.86 | 14.05 | 60.0 | |
| 3200 | 3 | 29 | 东北 | 0.50 | 708 | 8.85 | 18.90 | 56.7 | |
| 3300 | 3 | 35 | 东北 | 0.15 | 108 | 6.30 | 14.58 | 43.0 |
在每个样地随机挖取2个土壤剖面, 记录土壤类型、土层厚度、土壤质地等, 并按0—10、10—20、20—40、40—60 cm分层取土样, 将同样地同层次土样混合均匀为1份土样, 由于3300 m处土层厚度仅有43 cm, 仅取3层土样, 样品共计105份。带回实验室挑除其中的石块、根系、小动物等杂物, 做好标记装入自封袋, 带回实验室分析测定土壤有机碳。同时, 在每个土壤剖面按4层用环刀(体积为100 cm3)取样, 带回实验室测定容重等土壤物理性质。
2.3 样品处理及数据分析 2.3.1 乔木层生物量及碳密度计算(1) 生物量
根据王金叶等[21]对祁连山青海云杉解析木建立的林木各器官生物量相对生长模型, 利用单株树木的树高(H, m)和胸径(DBH, cm)计算出单株各器官生物量, 具体估算方程见表 2。
| 器官 Organs |
相对生长方程 Relative growth equation |
相关系数 Correlation coefficient |
| 干Stem | WS=0.0478(D2H)0.8665 | 0.9887 |
| 枝Branch | WB=0.0122(D2H)0.8905 | 0.9568 |
| 叶Leaf | WL=0.2650(D2H)0.4701 | 0.8622 |
| 根Root | WR=0.0478(D2H)0.8665 | 0.9707 |
| WS:干生物量, stem biomass;WB:枝生物量, shoot biomass;WL:叶生物量, leaf biomass;WR:根生物量, root biomass | ||
然后将各器官生物量相加获得整株树木的生物量(WT, kg), 将样地内所有树木生物量累加得到样地总生物量(W, t/hm2)[17], 具体方程式如下:
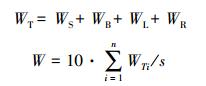
|
式中, WS, WB, WL, WR分别为单株树木的干、枝、叶和根生物量(kg);WTi为第i株树的生物量(kg);S为样地面积(m2);n为样地内树木株数;i为树木在样地内的编号。
(2) 乔木层碳密度
乔木层碳密度即为单位面积上乔木层生物量乘以转换系数(即干物质的碳质量分数), 通常生物量-碳换算系数通常是0.44—0.55, 本文参照已有报道的青海云杉各器官(干、枝、叶、根)碳转换系数平均值0.52作为青海云杉生物量-碳换算系数[21]。
2.3.2 土壤层碳密度计算土壤有机碳含量用重铬酸钾氧化-外加热法测定, 土壤容重采用浸泡法测定。
土壤层碳密度计算公式如下:
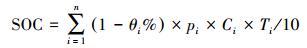
|
式中, SOC为土壤剖面有机碳密度(t/hm2);θi为第i层>2 mm砾石含量体积(%);pi为第i层土壤容重(g/cm3);Ci为第i层土壤有机碳含量(g/kg);Ti为第i层土壤厚度(cm);n为第i层参与计算的土壤层次总数。
2.3.3 生态系统碳密度森林生态系统碳密度由单位面积上森林乔木层和土壤层平均碳密度累计得到, 由于青海云杉林灌草本层和枯落物层碳密度所占整体碳密度比例较少, 本研究忽略了灌草本层和枯落物层碳密度的计算。
2.3.4 气候要素数据采用彭守璋等[19]依据祁连山青海云杉分布区域附近28个气象站1953—2008年的数据, 结合海拔和经纬度建立的气候要素与地理位置和海拔因子之间的多元线性回归方程, 来拟合各样地气候要素数据, 具体回归方程如下:
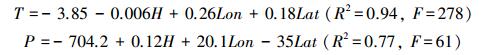
|
式中, T为夏季平均气温(℃);P为年均降水量(mm);H、Lon、Lat分别是海拔、经度、纬度。
2.3.5 数据处理采用Excel 2013和SPSS 19.0软件进行数据统计分析, 对碳密度采用单因素方程分析(one-way ANOVA), 且进行LSD多重比较, 以检验数据之间的差异显著性(P=0.05)。
3 结果与分析 3.1 不同海拔梯度青海云杉林生物量和碳密度变化及分配格局从表 3可知, 研究区青海云杉林生物量平均为115.83 t/hm2, 变化范围在7.66—197.10 t/hm2, 乔木层碳密度平均为60.23 t/hm2, 变化范围为3.90—102.49 t/hm2(图 1)。随海拔梯度增加, 生物量整体变化趋势为先增加后波动降低的趋势, 在各海拔梯度间具有极显著差异(P < 0.01), 海拔2800 m处最大, 3300 m处最低。从林分生物量各器官分配格局来看, 青海云杉在各海拔处均表现为干>根>枝>叶, 其中干生物量占林分总生物量的22.23%—72.70%, 根生物量分配比例为13.46%—40.24%, 枝生物量分配比例为6.94%—23.05%, 叶生物量分配比例为4.83%—15.01%, 各器官生物量随海拔梯度变化均存在极显著差异(P < 0.01), 且变化趋势与总生物量变化一致, 呈现先逐渐增加后波动降低趋势, 波动变化处为海拔2900—3200 m, 多重比较发现此海拔梯度变化差异性不显著(P>0.05)。
| 海拔梯度 Altitudinal gradient/m |
干生物量 Stem biomass/(t/hm2) |
枝生物量 Branch biomass/(t/hm2) |
叶生物量 Leaf biomass/(t/hm2) |
根生物量 Root biomass/(t/hm2) |
总生物量 Total biomass/(t/hm2) |
| 2500 | 43.81±3.73ab | 13.68±1.26ab | 9.52±0.48bc | 26.53±2.87bc | 93.54±3.53b |
| 2600 | 59.25±1.98ab | 18.58±0.63ab | 12.09±0.19bc | 32.75±0.29bc | 122.67±2.48ab |
| 2700 | 71.42±3.75ab | 22.27±1.19ab | 15.75±1.15ab | 43.36±3.90ab | 152.80±8.41ab |
| 2800 | 93.78±6.70a | 29.27±2.19a | 20.12±0.30a | 53.95±0.55a | 197.10±8.64a |
| 2900 | 63.21±18.84ab | 19.79±5.95ab | 13.53±3.39b | 37.53±8.13b | 134.06±36.30ab |
| 3000 | 72.98±8.15ab | 23.14±2.79ab | 13.30±1.25b | 35.61±5.76b | 145.03±9.79ab |
| 3100 | 43.12±10.24b | 13.57±3.35b | 8.96±2.93bc | 23.92±2.60bc | 89.26±15.41b |
| 3200 | 53.30±21.56b | 16.89±6.92b | 0.86±0.22c | 21.24±6.11c | 100.39±37.19b |
| 3300 | 3.27±1.13c | 1.01±0.35c | 0.86±0.22d | 2.52±0.51d | 7.66±2.20c |
| 表格内数据为Mean ± SE, 同列不同字母表示差异显著(P < 0.05) | |||||

|
| 图 1 不同海拔梯度青海云杉乔木层碳密度分布特征 Fig. 1 Distribution characteristics of carbon density of arbor layer of P. crassifolia forest in the different altitudinal gradient 图中不同字母代表不同土层之间差异显著(P < 0.05) |
另外, 由于植物碳密度是基于生物量和碳系数的乘积, 因此青海云杉乔木层碳密度随海拔梯度变化与生物量变化一致, 符合二项式分布, 拟合方程为y=-3.3986x2+28.877x+23.471, P < 0.01, 平均值为60.23 t/hm2, 最大值出现在海拔2800 m, 为197.10 t/hm2, 最小值出现在海拔3300 m, 为7.66 t/hm2。
3.2 不同海拔梯度青海云杉林土壤有机碳含量变化如图 2所示, 不同海拔梯度的青海云杉林土壤有机碳含量平均值为53.99 g/kg, 变化范围为31.49—75.02 g/kg, 不同海拔梯度间呈极显著差异(P < 0.01), 仅在2500—2700 m呈不显著降低外, 2700—3300 m为逐渐增加的趋势。随土壤层次的增加, 除海拔3200 m和3300 m的土壤有机碳含量未表现出规律变化外, 其他海拔梯度则均为逐渐降低趋势, 且仅在2700 m和3000 m时, 40—60 cm土壤有机碳含量显著低于其余3层土壤有机碳(P < 0.05), 而在其他海拔上各土壤层次有机碳含量则没有存在显著性差异(P>0.05)。

|
| 图 2 不同海拔梯度不同土壤层次土壤有机碳含量变化 Fig. 2 Changes on the content of soil organic carbon in different soil layer in the different altitudinal gradient 图中不同字母代表不同土层之间差异显著(P < 0.05) |
由表 4可知, 青海云杉土壤有机碳密度平均值为194.62 t/hm2, 各海拔梯度间存在极显著差异(P < 0.0001), 在海拔2900 m达到最大值, 为245.40 t/hm2, 在海拔2700 m处最低, 为130.24 t/hm2。随着海拔梯度上升来看, 低海拔2500—2700 m表现为平缓降低趋势, 从海拔2800 m处急剧上升, 约为海拔2700 m土壤碳密度的1.88倍, 且海拔2800—3200 m呈现轻度波动变化, 无显著性差异, 在海拔3300 m又急剧降低, 其值仅为3200 m处的59.48%。从不同土壤层次土壤碳密度随海拔梯度变化来看, 0—10 cm层土壤碳密度变化范围在26.41—43.28 t/hm2, 呈近似W型变化, 且海拔梯度上具有极显著差异(P < 0.01), 在2900 m处最大, 2700 m处最小;10—20 cm层土壤碳密度变化范围在23.51—40.58 t/hm2, 呈不规则的波动变化, 具有极显著差异(P < 0.001), 在2800 m处最大, 2600 m处最小;20—40 cm层土壤碳密度变化范围在46.39—85.73 t/hm2, 随海拔梯度增加整体呈先增加后降低的趋势, 并具有极显著差异(P < 0.001), 3100 m处最大, 2700 m处最小;40—60 cm层土壤碳密度变化范围在30.49—83.56 t/hm2, 整体呈先降低后增加的趋势, 在3200 m处最大, 2700 m处最小。
| 海拔梯度 Altitudinal gradient/m |
土壤层次Soil layer/cm | 合计 Total |
|||
| 0—10 | 10—20 | 20—40 | 40—60 | ||
| 2500 | 27.35±5.49a | 29.53±4.09bc | 52.38±2.14b | 52.04±8.21b | 161.30±9.17b |
| 2600 | 27.44±0.84a | 23.51±4.47c | 54.98±8.99b | 45.55±3.57bc | 151.49±14.97b |
| 2700 | 26.41±1.69a | 26.95±0.55bc | 46.39±5.56bc | 30.49±8.43c | 130.24±14.43b |
| 2800 | 40.67±0.76b | 46.47±1.65a | 77.98±3.90a | 80.28±6.05a | 245.40±6.15a |
| 2900 | 43.28±1.29b | 42.30±3.10a | 78.62±7.94a | 81.91±9.60a | 246.12±20.13a |
| 3000 | 31.78±1.88a | 38.58±3.52ab | 83.27±2.12a | 72.35±5.95ab | 225.97±11.41a |
| 3100 | 28.75±2.76a | 40.58±1.12ab | 85.73±3.14a | 80.43±5.47a | 235.49±9.83a |
| 3200 | 27.13±3.56a | 32.86±2.43b | 81.07±5.04a | 83.56±4.94a | 224.63±11.89a |
| 3300 | 31.00±0.56a | 33.61±1.83b | 69.00±7.92ab | — | 133.61±7.95b |
| 表格内数据为Mean ± SE, 同列不同小写字母代表差异显著(P < 0.05) | |||||
青海云杉生态系统平均总碳密度为255.15 t/hm2, 其中乔木层和土壤层占总碳密度的比例分别为23.61%和76.39%, 且不同海拔梯度间存在极显著差异(P < 0.0001)。随海拔梯度的上升, 表现为先增加后降低的趋势, 其中在2500—2700 m先缓慢增加, 在2800 m达到最大值, 为347.89 t/hm2, 随后又逐渐降低, 在海拔3300 m达到最低值, 为137.60 t/hm2(图 3)。从乔木层和土壤层碳密度所占总碳密度比例来看(图 4), 不同海拔梯度上, 乔木层所占比例变化范围为2.89%—37.89%, 土壤层所占比例变化范围为62.11%—97.11%。

|
| 图 3 青海云杉林生态系统总碳密度沿海拔梯度变化 Fig. 3 Variation of total carbon density of P. crassifolia forest ecosystem along altitudinal gradient 图中不同字母代表不同海拔之间差异显著(P < 0.05) |

|
| 图 4 青海云杉林乔木层和土壤层碳密度各占总碳密度比例沿海拔梯度变化 Fig. 4 Variation of the percentage of carbon density of arbor layer and soil layer to total carbon density of P. crassifolia forest along altitudinal gradient |
分析了祁连山青海云杉林乔木层碳密度和土壤层有机碳密度与不同影响因子间的相关性, 以了解影响其变化的主要因子(表 5)。海拔与夏季平均气温、林分密度、植被碳密度均呈现极显著负相关, 与年平均降水量、土壤有机碳含量、土壤全氮含量、土壤有机碳密度具有极显著正相关, 与碳氮比呈显著正相关;夏季平均气温与年平均降水量、土壤有机碳含量、土壤全氮含量、土壤有机碳密度具有极显著负相关, 与土壤碳氮比呈显著负相关, 与植被碳密度呈显著正相关;年平均降水量与土壤有机碳含量、土壤全氮含量、土壤有机碳密度呈极显著正相关, 与碳氮比呈显著正相关, 与林分密度呈显著负相关;土壤有机碳含量与土壤全氮含量、碳氮比、土壤有机碳密度呈极显著正相关, 与林分密度呈极显著负相关;土壤全氮含量与土壤有机碳密度呈极显著正相关, 与林分密度、胸高断面积、植被碳密度呈显著负相关;土壤容重与碳氮比具有显著负相关;林分密度、胸高断面积、植被碳密度两两之间均均具有极显著正相关。
| 指标 Indexes |
海拔 Altitudinal |
年夏季平均气温 Summer average temperature |
年平均降水量 Average annual precipitation |
土壤有机碳含量 Soil carbon content |
土壤全氮含量 Soil total nitrogen |
碳氮比 The ratio of carbon to nitrogen |
土壤容重 Soil bulk density |
土壤有机碳密度 Soil organic carbon density |
林分密度 Stand density |
胸高断面积 Breast height area |
植被碳密度 Vegetation carbon density |
| 海拔Altitudinal | 1 | ||||||||||
| 夏季平均气温 Summer average temperature |
-0.998** | 1 | |||||||||
| 年平均降水量 Average annual precipitation |
0.980** | -0.987** | 1 | ||||||||
| 土壤有机碳含量 Soil organic carbon content |
0.874** | -0.868** | 0.828** | 1 | |||||||
| 土壤全氮含量 Soil total nitrogen |
0.601** | -0.595** | 0.576** | 0.739** | 1 | ||||||
| 碳氮比 The ratio of carbon to nitrogen |
0.390* | -0.476* | 0.443* | 0.502** | -0.194 | 1 | |||||
| 土壤容重 Soil bulk density |
-0.152 | 0.136 | -0.153 | -0.261 | 0.049 | -0.384* | 1 | ||||
| 土壤有机碳密度 Soil carbon organic density |
0.550** | -0.556** | 0.534** | 0.640** | 0.529** | 0.291 | 0.145 | 1 | |||
| 林分密度 Stand density |
-0.571** | 0.545** | -0.438* | -0.513** | -0.408* | -0.216 | -0.132 | -0.236 | 1 | ||
| 胸高断面积 Breast height area |
-0.312 | 0.292 | -0.194 | -0.302 | -0.400* | 0.096 | -0.284 | 0.015 | 0.768** | 1 | |
| 植被碳密度 Vegetation carbon density |
-0.449** | 0.425* | -0.319 | -0.417* | -0.433* | -0.015 | -0.166 | -0.087 | 0.854** | 0.970** | 1 |
| *在0.05水平(双侧)上显著相关;**在0.01水平(双侧)上显著相关 | |||||||||||
同时, 为了进一步对影响植被碳密度和土壤有机碳密度的影响因子进行区分, 利用多元逐步回归分析方法, 构建了植被碳密度和土壤碳密度与主要影响因子的模拟关系式:
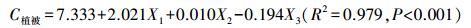
|
式中, C植被为植被碳密度, X1为胸高断面积, X2为林分密度, X3为土壤有机碳含量。
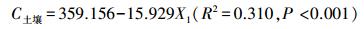
|
式中, C土壤为土壤有机碳密度, X1为年年夏季平均气温。
由拟合关系发现, 影响植被碳密度的主要因子包括胸高断面积、林分密度和土壤有机碳含量, 而影响土壤有机碳密度的主要因子为年夏季平均气温。
4 结论与讨论 4.1 祁连山青海云杉林生物量和植被碳密度随海拔变化本研究区域青海云杉乔木层生物量平均值为115.83 t/hm2, 与张雷等[17]在此区域研究结果(128.61 t/hm2)以及祁连山北坡平均值(109.24 t/hm2)[19]相近, 但低于祁连山区域平均值(169.80 t/hm2)[22], 主要与取样数量以及取样位置和林分质量差异有关, 且由于祁连山地区水热条件空间差异性较大[19], 导致不同区域青海云杉生长存在差异。随海拔梯度的增加, 青海云杉乔木层生物量和碳密度总体表现为先增加后降低趋势, 海拔2500—3200 m相对较好, 且在2800 m处达到最大, 在高山林线处的3300 m处急剧降低, 呈现明显的单峰变化趋势, 与张雷等研究结果相一致[18], 主要是由于青海云杉为喜阴湿, 水分和热量不充足、较强的日照都不利于其生长发育[23]。随海拔梯度升高, 温度逐渐降低而降水则增加, 往往在中间海拔区段形成良好的水热组合[24], 更适合青海云杉的生长, 而森林分布的海拔上、下限, 生境条件恶劣, 水分或热量条件亏缺, 导致青海云杉生长受到抑制[16, 18]。从相关分析可知, 植被碳密度与海拔呈极显著负相关, 与土壤全氮含量呈显著负相关, 与夏季平均气温、林分密度、胸高断面积为极显著正相关, 可见除了林分自身生长特征有关的指标外, 影响光照、温度、土壤养分、干扰状况等多种外部因素也成为影响其[19]。进一步逐步回归分析发现, 林分胸高断面积、林分密度和土壤有机碳含量成为影响祁连山青海云杉乔木层碳密度的关键生态因子, 赵维俊等[16]研究也发现土壤营养物质影响该区域青海云杉生长发育的关键生态因子。另外, 接近林线的海拔3300 m处土壤养分状况较好, 由于高海拔区域的低温、高光照和多降水有利于青海云杉林乔木层碳密度累积, 土壤条件并不是青海云杉林生长的制约因素, 因此, 未来随着全球气候变暖趋势下, 青海云杉生长上限有可能进一步增加其生长范围和优势, 导致乔木层生物量不断的增加[16]。
4.2 祁连山青海云杉林土壤有机碳密度随海拔变化土壤有机碳含量的多少主要取决于有机质的输入和分解速率, 输入与气候条件、土壤水分和养分状况、植被残体归还量以及人为扰动等因素, 而有机质分解速率更多与微生物数量、土壤温度和水分等有关[10, 13, 25]。海拔梯度作为环境因子的综合反映, 随着海拔梯度的增加, 土壤碳含量会逐渐增加, 由于海拔升高, 气温降低而水分含量增加, 导致土壤微生物活性降低或受到抑制, 林地凋落物和地被物分解速率变缓, 导致有机碳累积量增加[16, 25], 本研究也具有此种变化规律, 且土壤碳含量与海拔、年平均降水量、土壤有机碳含量、土壤全氮含量呈显著正相关, 与夏季平均气温存在显著负相关, 主要是与影响土壤有机碳累积和分解的环境因子有关。同时, 随着土壤深度的增加, 土壤碳含量在海拔2500—3100 m均呈现降低, 主要源于土壤有机碳来源地表植被枯落物分解, 且土壤环境条件和微生物活性又随土壤层次增加而逐渐降低[13], 而高海拔3200—3300 m未具有此种规律, 可能与林线环境恶劣且降水量变大有关。同时, 据已有研究发现, 对于同类植被在海拔梯度上有机碳状况是反映温度梯度作用的结果, 而土壤碳储量则没有明显分布规律[4], 本研究结果也证实了此结论, 逐步回归分析结果表明年夏季平均气温成为影响土壤有机碳密度的关键生态因子。另外, 本研究中乔木层碳密度与土壤层碳密度之间不存在显著相关, 主要是由于该区域青海云杉植被在地理空间上变异较高有关[19], 而高山区域的天山云杉林土壤碳密度变化与生物量则存在显著正相关[14], 说明不同区域环境异质性变化、植物生物特性以及人为干扰情况均会在一定程度上改变地上植被和地下土壤周转和循环过程。
4.3 青海云杉林生态系统碳密度本研究发现, 青海云杉生态系统平均碳密度为255.15 t/hm2, 接近我国森林生态系统的平均碳密度258.83 t/hm2 [26]。乔木层和土壤层碳密度分别占总碳密度的23%和77%, 这与王金叶等[21]研究的结果相近, 说明土壤碳密度仍是森林生态系统最重要的碳库[27]。同时, 由于祁连山地区海拔和经纬度的跨度大, 气温和降水的空间变幅较大, 导致青海云杉生态系统碳密度在海拔梯度上空间变异很大, 但以中部海拔区段2800—3200 m范围内青海云杉生长较好的生态系统碳密度较高, 平均达到304.77 t/hm2, 在长白山[8]研究也发现植被和气候变化条件成为调节土壤有机碳变化的关键因子, 说明水热条件变化、植被长势以土壤属性变化共同决定了青海云杉林生态系统有机碳库的大小。
本研究虽然依照祁连山青海云杉沿海拔梯度分布生长界限进行样地设置和取样, 对于解释地上植被和土壤碳密度与海拔变化的关系是可行的, 但是由于样地数量有限, 且未考虑枯落物层、灌草层以及枯立木、枯倒木等层次, 同时山地生态系统具有较大的空间变异性, 因此对于准确评估该区域青海云杉林生态系统碳密度仍显不足。
| [1] | 刘彦春, 张远东, 刘世荣, 张笑鹤. 川西亚高山针阔混交林乔木层生物量、生产力随海拔梯度的变化. 生态学报, 2010, 30(21): 5810–5820. |
| [2] | 徐影, 丁一汇, 李栋梁. 青藏地区未来百年气候变化. 高原气象, 2003, 22(5): 451–457. DOI:10.3321/j.issn:1000-0534.2003.05.004 |
| [3] | Bonan G B. Forests and climate change:forcings, feedbacks, and the climate benefits of forests. Science, 2008, 320(5882): 1444–1449. DOI:10.1126/science.1155121 |
| [4] | 秦纪洪, 王琴, 孙辉. 川西亚高山-高山土壤表层有机碳及活性组分沿海拔梯度的变化. 生态学报, 2013, 33(18): 5858–5864. |
| [5] | Ohtsuka T, Hirota M, Zhang X Z, Shimono A, Senga Y, Du M G, Yonemura S, Kawashima S, Tang Y H. Soil organic carbon pools in alpine to nival zones along an altitudinal gradient (4400-5300 m) on the Tibetan Plateau. Polar Science, 2008, 2(4): 277–285. DOI:10.1016/j.polar.2008.08.003 |
| [6] | Pregitzer K S, Euskirchen E S. Carbon cycling and storage in world forests:biome patterns related to forest age. Global Change Biology, 2004, 10(12): 2052–2077. DOI:10.1111/gcb.2004.10.issue-12 |
| [7] | Johnson D W, Curtis P S. Effects of forest management on soil C and N storage:meta analysis. Forest Ecology and Management, 2001, 140(2/3): 227–238. |
| [8] | Zhang M, Zhang X K, Liang W J, Jiang Y, Dai G H, Wang X G, Han S J. Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain, China. Pedosphere, 2011, 21(5): 615–620. DOI:10.1016/S1002-0160(11)60163-X |
| [9] | 向慧敏, 温达志, 张玲玲, 李炯. 鼎湖山森林土壤活性碳及惰性碳沿海拔梯度的变化. 生态学报, 2015, 35(18): 6089–6099. |
| [10] | Zhu B, Wang X P, Fang J Y, Piao S L, Shen H H, Zhao S Q, Peng C H. Altitudinal changes in carbon storage of temperate forests on Mt Changbai, Northeast China. Journal of Plant Research, 2010, 123(4): 439–452. DOI:10.1007/s10265-009-0301-1 |
| [11] | Sheikh M A, Kumar M, Bussmann R W. Altitudinal variation in soil organic carbon stock in coniferous subtropical and broadleaf temperate forests in Garhwal Himalaya. Carbon Balance and Management, 2009, 4: 6. DOI:10.1186/1750-0680-4-6 |
| [12] | 向成华, 栾军伟, 骆宗诗, 宫渊波. 川西沿海拔梯度典型植被类型土壤活性有机碳分布. 生态学报, 2010, 30(4): 1025–1034. |
| [13] | 杜有新, 吴从建, 周赛霞, 黄良, 韩世明, 徐雪峰, 丁园. 庐山不同海拔森林土壤有机碳密度及分布特征. 应用生态学报, 2011, 22(7): 1675–1681. |
| [14] | 阿米娜木·艾力, 常顺利, 张毓涛, 仇瑶, 何平. 天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素. 生态学报, 2014, 34(7): 1626–1634. |
| [15] | 丛静, 刘晓, 卢慧, 薛亚东, 刘学端, 尹华群, 李迪强, 张于光. 神农架自然保护区土壤微生物生物量碳、氮沿海拔梯度的变化及其影响因素. 生态学杂志, 2014, 33(12): 3381–3387. |
| [16] | 赵维俊, 刘贤德, 成彩霞, 李小燕. 祁连山排露沟流域中山带青海云杉林生长与海拔梯度上环境因子的关系. 中南林业科技大学学报, 2015, 35(2): 72–78. |
| [17] | 张雷, 于彭涛, 王彦辉, 王顺利, 刘贤德. 祁连山北坡青海云杉中龄林生物量随海拔的变化. 林业科学, 2015, 51(8): 1–7. |
| [18] | 张雷, 于彭涛, 王彦辉, 王顺利, 刘贤德, 金铭, 张学龙. 祁连山北坡青海云杉中龄林结构随海拔的变化. 林业科学研究, 2015, 28(4): 557–564. DOI:10.3969/j.issn.1001-1498.2015.04.016 |
| [19] | 彭守璋, 赵传燕, 郑祥霖, 许仲林, 何磊. 祁连山青海云杉林生物量和碳储量空间分布特征. 应用生态学报, 2011, 22(7): 1689–1694. |
| [20] | 何志斌, 赵文智, 刘鹄, 苏永中. 祁连山青海云杉林斑表层土壤有机碳特征及其影响因素. 生态学报, 2006, 26(8): 2572–2577. DOI:10.3321/j.issn:1000-0933.2006.08.020 |
| [21] | 王金叶, 车克钧, 蒋志荣. 祁连山青海云杉林碳平衡研究. 西北林学院学报, 2000, 15(1): 9–14. DOI:10.3969/j.issn.1001-7461.2000.01.002 |
| [22] | 许仲林.祁连山青海云杉林地上生物量潜在碳储量估算[D].兰州: 兰州大学, 2011. |
| [23] | 刘铮, 白英, 赵传燕, 南忠仁. 气候影响下祁连山自然保护区青海云杉林材积生长量差异性研究. 干旱区资源与环境, 2014, 28(7): 171–176. |
| [24] | Namgail T, Rawat G S, Mishra C, Van Wieren S E, Prins H H T. Biomass and diversity of dry alpine plant communities along altitudinal gradients in the Himalayas. Journal of Plant Research, 2012, 125(1): 93–101. DOI:10.1007/s10265-011-0430-1 |
| [25] | 张鹏, 张涛, 陈年来. 祁连山北麓山体垂直带土壤碳氮分布特征及影响因素. 应用生态学报, 2009, 20(3): 518–524. |
| [26] | 周玉荣, 于振良, 赵士洞. 我国主要森林生态系统碳贮量和碳平衡. 植物生态学报, 2000, 24(5): 518–522. DOI:10.3321/j.issn:1005-264X.2000.05.002 |
| [27] | 申家朋, 张文辉, 李彦华, 何景峰, 张辉. 陇东黄土高原沟壑区刺槐和油松人工林的生物量和碳密度及其分配规律. 林业科学, 2015, 51(4): 1–7. |